
Abstract
Passively dispersing organisms should optimise the time and direction of dispersal by employing behaviours that increase their probability of being successfully transported by dispersal agents. We rigorously tested whether two agriculturally important passively-dispersing eriophyoid species, wheat curl mite (WCM) and cereal rust mite (CRM), display behaviours indicating their readiness to depart from current host plants in the presence of potential dispersal cues: wind, an insect vector and presence of a fresh plant. Contrary to our expectations, we found that both species decreased their general activity in the presence of wind. When exposed to wind, WCM (but not CRM) significantly increased behaviour that has previously been considered to facilitate dispersal (in this case, standing vertically). Our study provides the first sound test of the function of what have been interpreted as dispersal-related behaviours of eriophyid mites. The low proportion of WCM exhibiting dispersal behaviour suggests there may be predisposed dispersers and residents in the population. Moreover, we found that WCM was generally more active than CRM, which is likely a contributing factor to its high invasive potential.
Similar content being viewed by others

Introduction
Dispersal is a fundamental biological process that has consequences for population dynamics, population genetic structure, and geographical distribution of species1,2,3,4,5. At the individual level, dispersal can influence fitness by reducing inbreeding and competition among kin, by distributing offspring from the same parents over different conditions (‘bet hedging’), or by allowing individuals to escape unfavourable local conditions6, 7. However, this process also involves costs that can be incurred at every stage of dispersal, i.e., departure, transit, and settlement2. Costs of dispersal may differ in actively vs. passively dispersing organisms. An actively dispersing organism can control its own movement and can make decisions about timing, direction and speed in all three stages of dispersal. In contrast, in passively dispersing organisms, direction and speed of movement are largely outside of their control and depend instead on external forces such as gravity, wind or water currents, or on other organisms that act as vectors6. In particular, passive dispersers cannot control the transit and settlement stages of dispersal. However, they may partially control departure, e.g., by making the decision whether to take off and by engaging in behaviours that increase the possibility of being lifted by dispersal agents8. Existence of directed dispersal in passively dispersing organisms has recently been proposed for plants9. As another example, ‘pre-ballooning’ behaviours in spiders increase the probability of their being picked up by the wind, and include raising the abdomen, producing silk and jumping10. Additionally, spiders can choose the time of ballooning based on wind conditions11 and cloud cover12. Pre-dispersal postures have also been well documented in scale insect (Coccoidea)13, predatory phytoseiid mites14,15,16 and phytophagous tetranychid spider mites17,18,19, which additionally exhibit collective behaviours facilitating dispersal20, 21, and have been proposed to occur in phytophagous eriophyoid mites22.
Eriophyoid mites (Eriophyidae, Phytoptidae and Diptilomiopidae) are obligatory plant parasites that are limited in ambulatory dispersal due to their extremely small size (most are less than 300 µm long), vermiform body shape, and possession of only two pairs of short anteriorly positioned legs (Fig. 1a and b). Passive aerial or phoretic dispersal has been suggested as the major way that eriophyoids spread22, 23. Behaviours that have been interpreted as facilitating dispersal in these mites include standing up on their anal lobes, arching their bodies and moving their legs rapidly, raising the hind part of the body while standing on their forelegs (all of which have been termed ‘take-off postures’), as well as forming groups of attached individuals (chains)23,24,25,26 (Fig. 1c and d). Additionally, these ‘take-off postures’ have been observed in eriophyoid mites when on an unsuitable host plant species and have been interpreted as indicating host-rejection and intent to disperse27.

SEM images of eriophyid mites. Scale bars are 10 μm on each photo. (a) – Wheat curl mite (Aceria tosichella WCM MT-1 genotype) on wheat leaf; (b) – Cereal rust mite (CRM, Abacarus hystrix) on wheat leaf; (c) – standing-erect mite belonging to the WCM complex; (d) – chain- forming mites belonging to the WCM complex.
The decision whether or not to disperse is a trade-off between risky movement and possible benefits from successful colonization1. The energetic investment in dispersal behaviours or the production of drifting structures can be costly10, 28, 29. Dispersal may expose individuals to predators (mortality costs)30. Finally, making an incorrect decision and taking off under unsuitable environmental conditions may increase the mortality during transit or settlement31. Direct mortality costs in aerially dispersing arthropods may increase with prolonged time spent in the air due to depletion of energy and water reserves10. Eriophyoid mites are especially at risk of dispersal-associated starvation as they are relatively host-specific and cannot survive for long if they land on an inappropriate plant species23.
Although it has been well documented that departure-specific behaviours affect the likelihood of aerial dispersal of spider mites19, 32, there has been no empirical evidence that similar behaviours in eriophyoid mites promote the probability of their becoming airborne or being picked up by vector organisms. Melo et al.33 found that aerial dispersal in the eriophyid coconut mite, Aceria guerreronis Keifer, is only occasionally preceded by a raised body stance at take-off. They concluded that raised body position may assist departure but it is not a prerequisite for take-off. It has also been argued that eriophyoid mites are simply blown off accidentally from their host plants and that they may have no specific behavioural adaptations to promote dispersal33, 34. Conversely, increased walking activity in the presence of a suitable host plant, due to chemical cues released by the plant, has been suggested as an important mode of eriophyoid dispersal22, 23. Additionally, this increased activity may increase the chance of the mite being picked up by the wind and potentially being blown to the nearby plant. Undoubtedly, the question whether eriophyoid mites have evolved any specific behaviours playing a role in their dispersal and colonization success remains unanswered.
Here we test whether specific departure behaviours occur in two very tiny (body length ca. 0.20–0.25 mm) cereal-feeding eriophyid mites by investigating their behavioural responses to the presence of three potential dispersal signals: wind, insects that could act as vectors, and fresh foliage from a suitable host plant. We used two economically important species with worldwide distributions: Aceria tosichella Keifer, wheat curl mite (WCM), and Abacarus hystrix (Nalepa) (sensu lato), cereal rust mite (CRM). Both belong to species complexes consisting of lineages that can be distinguished principally by differences in cytochrome c oxidase subunit I (COI) gene fragment35, 36. We tested the hypothesis that in the presence of dispersal cues mites display behaviours indicating their readiness to disperse, including increasing walking activity, initiating take-off postures and forming chains of individuals. Additionally we examined whether the particular behaviours shown by the mites are influenced by the type of dispersal cue or rather are invariant within a species, and whether responses to cues differ between the two species.
Results
All mite behaviours (feeding, walking, standing erect and chain formation) were strongly and significantly influenced by species and treatment, but were not influenced by their interaction (Table 1).
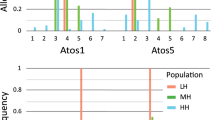
Wind significantly influenced the proportions of mites feeding and walking for both species, and the proportion standing erect for WCM (Fig. 2, see also Supplementary Tables S1 and S2). Higher probabilities of feeding in CRM (83.3%, 95% CI = 76.1–89.2%) and WCM (37.2%, 95% CI = 29.4–45.5%) but lower probabilities of walking in CRM (14.1%, 95% CI = 9.1–20.3%) and WCM (55.0%, 95% CI = 47.3–62.5%) were observed in the presence of wind when compared to other treatments. The probability of WCM standing erect in the presence of wind was low (4.0%, 95% CI = 3.1–5.2%), but nonetheless was significantly higher when compared to the proportion standing erect in other treatments (Fig. 2, see also Supplementary Table S2). The presence of fresh plant material significantly influenced WCM behaviour resulting in lower probability of mites forming chains (0.9%, 95% CI = 0.5–1.4%) compared to other treatments (Fig. 2, Supplementary Table S2). There was no significant behavioural response of mites to the presence of a potential insect vector (the thrips Anaphothrips obscurus Müller) (Fig. 2).

Results of Simultaneous Tests for General Linear Hypotheses. Effects of potential dispersal cues on the proportion of mites displaying particular behaviours (feeding, walking, standing erect, chain formation) when comparing to the control for the two species of eriophyid mites. Values on vertical axes are partial residuals. Significance codes: ***p < 0.001; **p < 0.01; *p < 0.05; ·p < 0.1. Exact p values can be found in Supplementary Table S2.
Overall, CRM had a higher proportion of individuals feeding (68.5%, 95% CI = 64.2–72.6%), but lower proportions walking (29.5%, 95% CI = 25.7–33.5%), standing erect (1.4%, 95% CI = 1.1–1.7%), and forming chains (1.2%, 95% CI = 0.9–1.5%) than did WCM (feeding: 23.3%, 95% CI = 19.6–27.3%; walking: 70.9%, 95% CI = 66.9–74.7%; standing erect: 2.1%, 95% CI = 1.7–2.6%; chain formation: 2.0%, 95% CI = 1.6–2.4%).
Discussion
Passive dispersal by wind or by larger organisms is a common phenomenon in wingless, small-bodied organisms, which are not able to control the direction of their movement once airborne or on the body of a vector37, 38. However, timing of the launch phase of passive dispersal can potentially be actively controlled by the dispersing individuals8. In this study we analyzed the behaviours that have been long suggested to facilitate passive dispersal in eriophyoid mites, the smallest plant-feeding arthropods. The three potential dispersal cues tested in our study reflect three possible modes of eriophyid mite dispersal: (i) air dispersal: wind; (ii) active walking: fresh plant presence; (iii) phoresy: insect vector presence. We hypothesized that mites would change their behaviour by increasing walking activity, initiating take-off postures and forming chains of individuals when dispersal cues were present. However, contrary to our expectation, mites did not increase walking in the presence of any of the potential dispersal cues, and moreover they did not respond at all to the insect vector. Dispersal by biotic vectors has been suggested as the dispersal mode in eriophyid mites by several authors, as eriophyoid mites have sometimes been found attached to the bodies of larger arthropods24, 39,40,41,42. Moreover, Liu et al.43 found very strong evidence for obligate phoresy of an eriophyid mite, Aceria pallida Keifer, which uses the psyllid Bactericera gobica (Loginova) as a vector when moving to its winter hibernation sites on its woody perennial host. One of our test species, the wheat curl mite (WCM), has been found on thrips and aphids (reviewed in Michalska et al.)22. However, some authors have doubted that the phoresy plays a major role in the spreading of eriophyoid mites. Lindquist & Oldfield34 pointed out that eriophyoids do not show any clear morphological adaptations for attaching to hosts, and Galvão et al.44 did not find efficient dispersal of eriophyoids with insect vectors. Our results also indicate the lack of evident behavioural adaptations to phoretic dispersal. It is possible that attaching to insect vectors by the studied grass-feeding eriophyoid mites (reviewed in Michalska et al.22 as well as our own observations) is accidental rather than intentional. However, even if accidental, such attachment might occasionally result in dispersal of eriophyoids to new areas. It is also possible that phoresy may be common in eriophyoid mites inhabiting long-lived hosts (such as A. pallida mentioned above) and accidental or very rare in eriophyoids associated with short-lived hosts, such as grasses.
The mode of movement that has been suggested as the most efficient for eriophyoid mites is dispersal by wind currents23, 34, similar to plant-associated spider mites45, 46 and phytoseiid mites15. Indeed, our study suggests that wind was the main factor changing mite behaviour in both studied species. However, our results were not consistent with the hypotheses existing in the literature. Instead of behaving in a manner that would increase the probability of being picked up by the wind, both mite species reduced their activity by increasing feeding and decreasing walking in the wind presence. This could be interpreted as unwillingness to being dispersed.
Another explanation could be attributed to some of our experimental conditions. Outcomes of this and previous studies on eriophyoid dispersal were drawn from the behaviour of mites held at one particular light, wind speed, temperature and humidity regime. These environmental variables may affect dispersal propensity, distance travelled, locomotor performance or speed47,48,49,50. In particular, temperature-related factors have a large impact on behaviour of ectotherms since it directly affects metabolic rates, and can alter activity patterns or movement speed47. In wind-borne arthropods, dispersal propensity is often strongly correlated with meteorological conditions51,52,53,54. For example, launch postures of spiders and phytoseiid mites frequently occur in conditions of warm ambient temperature and light breezes12, 14, 51,52,53,54,55. Moreover, spider ballooning is known to be positively correlated with increased cloud cover12 while wind speed is an important factor determining successful take-off and transfer52, 54. The velocity of 4.2 m/s used in our study may seem to be relatively high when compared with studies on other arthropods. For example, spider ballooning was most common at speeds below 3 m/s52, and take-off response of phytoseiid mites was maximal at 2 m/s; however their dispersal activity was still very high at speeds of 4 m/s and 8 m/s55. Dispersal of other eriophyoid species at wind speeds of 4–4.5 m/s has been shown to be efficient33, 39, 56. It is possible that different groups of air-borne arthropods require different wind speed for effective dispersal due to morphological differences between them. Indeed, spiders, phytoseiids, tetranychids and eriophyoids differ in their body size and shape, setae number and length, and use of silk, all of which may influence the probability of being blown away at different wind speeds. Even if they are not phylogenetically related, seeds and spores of some plants and fungi present morphological adaptation to particular wind conditions and their propensity to be blown away tends to decrease at high humidity57, 58. Changes in other abiotic factors may also influence the propensity of eriophyid mites to disperse (e.g., mites may be more inclined to engage in wind dispersal when humidity is high and less likely to result in desiccation). It has been suggested that passively dispersed organisms select conditions at take-off that maximize their likelihood of successful dispersal because this will increase their expected fitness after landing59. Undoubtedly, understanding the factors that influence the evolution of dispersal propensity and ability is of great interest to evolutionary biologists and ecologists. Thus, further experiments should investigate above mentioned abiotic conditions to appraise the practical relevance of wind tunnel experiments and to test the generality of our findings for these and other species of plant-feeding mites.
However, WCM did respond to the wind presence by increasing the proportion standing erect (Fig. 2), a posture that has been interpreted as a “dispersal position”, but the proportion of standing WCM mites remained low. It is possible that only a small fraction of individuals in a given population of eriophyids are inclined to disperse at any given time. The existence of genetically and phenotypically distinct dispersers and non-dispersers (residents) in a single population has been demonstrated in almost all taxonomic groups, from unicellular organisms to insects and mammals1, 51, 60. Dispersers may differ from residents in having physiological, morphological or behavioural traits associated with increased dispersal success, such as fertilization status61, tolerance to environmental stress, larger or smaller body size, or greater general activity1, 62,63,64,65,66. The small proportion of WCM mites that engaged in the standing-up posture may suggest that in our lab population, there are only a few individuals predisposed to disperse. Also, Cote et al.67 wondered whether dispersing individuals differ in behaviour from residents across their lives or only when the decision to disperse is about to be made and appropriate dispersal conditions exist. Behaviour of mites in our study was more supportive of the second hypothesis, as they changed their general activity and took up standing-erect position significantly more often when the potential dispersal cue of wind was present.
On the other hand, Melo et al.33 urged caution in interpretation of standing erect as “the dispersal posture”, since other functions of this behaviour may be possible as well. For example, according to Lindquist & Oldfield34, chemosensory sensilla (solenidia) present on the tips of the mite’s legs can better inform the mite about the chemical composition of the environment when the mite stands up and stretches out its legs. Thus, standing erect position may be a helpful behaviour in detecting host plants, predators, competitors or potential mates. The role of this behaviour needs to be studied in more detail in the future.
It has been suggested that the formation of chains is helpful to take-off in aerial dispersal by increasing the overall surface area exposed to wind24, 68. In our case, mites did not significantly increase chain formation in the presence of wind. There is a possibility that this behaviour does not facilitate eriophyid dispersal. Similar to the standing-erect position, chain formation may have other functions, e.g. chemical or mechanical communication among individuals (reviewed in Michalska et al.)22. It is also possible that chain forming occurs only seasonally in wheat-feeding mites (e. g. during crop maturation), as was proposed by Nault & Styer68.
Our results showed that both studied eriophyid species (WCM and CRM) expressed similar patterns of behavioural responses, e.g. increased feeding and decreased walking in the presence of wind (Fig. 2). However, there were significant behavioural differences between WCM and CRM (Table 1), reflected mainly by higher general activity of WCM. This result fits with current knowledge of the ecology of WCM MT-1, which is the most important eriophyid agricultural pest as well as the most invasive and globally distributed WCM genotype69, 70. High activity of WCM may facilitate its dispersal and spreading as well as its invasive potential. Indeed, it has been shown that many invasive or successful alien species are better dispersers when compared to native or non-invasive ones71.
In conclusion, we did not find any strong evidence that two studied species of eriophyids responded to any of the provided cues (wind, fresh plants, potential phoretic hosts) in a way that could be interpreted as intention to disperse, except for the slightly higher proportion of WCM mites standing erect in the presence of wind. This indicates that we need to look more closely at the function of behaviours and activity of these tiny and economically important mites and also raises many interesting questions about how and when they do disperse in field conditions. This is of particular importance in cases of invasive species whose spread may be aided by global changes in environmental conditions.
Methods
Experimental plants and stock colonies
In our study we focused on WCM lineage MT-1 (see Skoracka et al.)72 and CRM A. hystrix (sensu stricto), both of which are well adapted to feed on wheat. All stock colonies as well as all experimental animals in this study were maintained on bread wheat, Triticum aestivum L. growing in pots from commercially available seeds. Stock colonies of mites were maintained under laboratory conditions (22–24 °C, photoperiod 16/8, 60% RH) in rearing cages consisting of metal frames wrapped with nylon bags. The stock colony of each species was kept in 7–10 separate pots as a safeguard should one or more fail. To keep the colony homogenous, wheat leaves with mites were transferred between pots every two months. New pots of wheat were put into rearing cages when the previous host plant wilted. WCM and CRM were held in separate laboratories in the Faculty of Biology, Adam Mickiewicz University. WCM MT-1 specimens were collected from wheat in Choryń (loc. 52°02′36″N 16°46′02″E, GenBank Acc. No: JF920077), and specimens of CRM were collected from quack grass, Elymus repens (L.) Gould, in Poznań (loc. 52°28′04″N 16°55′36″E, GenBank Acc. No: FJ387550.1). Field-collected specimens were transferred under a stereomicroscope from the collected plant shoots to potted wheat using an eyelash glued to a preparatory needle. In the course of establishing each stock colony several specimens were collected into Eppendorf tubes containing 180 µL of ATL buffer (Qiagen GmbH) for DNA extraction and subsequent DNA sequencing to confirm the molecular identification of studied mites. For DNA isolation a non-destructive method of DNA extraction was performed73. The cytochrome c oxidase subunit I (COI) gene fragment was amplified by PCR with the degenerate primers bcdF01 and bcdR04 following Skoracka & Dabert35. Subsequently, the PCR amplicon (650 bp) was sequenced with BigDye Terminator v3.1, in accordance with the manufacturer’s instructions, and products from sequencing reactions were analyzed on an ABI Prism 3130XL or 3730 Analyzer (Applied Biosystems). Trace files were checked, edited and aligned using MEGA6 (Tamura et al.74), and subsequently compared with reference sequences of each studied species downloaded from GenBank. This procedure was repeated every few weeks to test for evidence of contamination of one species’ colony by the other species. No such contamination was ever observed.
Anaphothrips obscurus Müller (Thysanoptera), the most abundant thrips species on wheat and other grasses in Poland75, was used as a potential dispersal vector for mites. We chose A. obscurus because it is associated with grasses including cereals and on our preliminary observations of 12 530 field-collected grass shoots: this species was the most common winged insect on wheat infested by WCM and CRM. Moreover during inspections we found WCM and CRM attached to the thrips specimens. Additionally, A. obscurus inhabits and explores the same plant microhabitats as WCM and CRM (e.g., leaf sheaths and bases, leaf furrows, inner parts of the leaf), which increases the likelihood of contact between eriophyoids and thrips compared to other insects (e.g., bees and syrphids) that visit grasses sporadically and land mostly on the outer leaf surfaces and on flowers. Other authors40 have found grass-associated eriophyoid mites on thrips. Thrips were collected from quack grass in Huby Moraskie (52°28′04.3″N 16°55′36.2″E). The plant shoots were inspected using a stereo-microscope (Olympus SZ40) and thrips were transferred into potted wheat using a small paintbrush. For species identification, thrips specimens were mounted on microscopic slides and examined under a compound microscope (Olympus BX41) and identified using Mirab-Balou et al.76.
Experimental design
In stock colonies, mites were allowed to multiply for ca. 30 generations under laboratory conditions (22–24 °C, photoperiod 16/8, 60% RH), ensuring a sufficient population size for the experiments (mean number of mites per leaf ~1500, which according to our earlier observations can be considered as highly crowded conditions). We examined the stock colonies in pots every few days and for experiments we chose leaves with similar (ca. 1500 specimens) densities. We ensured that all tested individuals within the species were descendants from the same stock colony in which population density was relatively stable. Therefore, variation in population density was not a factor influencing the behaviour of our experimental specimens. Additionally, leaves taken from the same pot for the purpose of behavioural observations were exposed to different treatments of the experiment, which ensures relatively even distribution of population densities across treatments. Additionally, we found that mites behavioural responses were independent of density (see Supplementary Methods online and Supplementary Figs S1 and S2).
All observations were carried out using a stereo-microscope (Olympus SZ51) and the whole procedure was identical for both eriophyid mite species. A fragment of wheat leaf (1.5 cm length) was cut and glued to a circular piece of adhesive tape (∅ 2.5 cm) to provide an arena for observations. The slow drying of the leaf during the experiment was expected to act as a signal for mites to disperse. Mean density on the cut leaves was 73.1 (95% CI: 61.2–86.5) for CRM and 132.2 (95% CI: 112.9–153.6). Mites could not walk cross the band of adhesive tape surrounding the leaf fragment. For WCM observations, the leaf fragment was cut near the leaf sheath, which is a place occupied by WCM69, whereas for CRM observations, the median leaf part was cut, since this species inhabits whole leaves27. The tape with the leaf fragment was placed into a small petri dish or on a square glass plate. Just after cutting the leaf, the number of mites of all life stages in the arena was counted to assess the size of the observed group. After 5 minutes, mite behaviour was surveyed by counting the number of individuals engaged in each activity listed below (see Testing influence of potential dispersal cues section). During each observation period, the behaviour of each mite was checked once and immediately recorded. Each observation period was replicated at least 20 times for each mite species and for each dispersal cue plus control. Each replicate was observed for three minutes. Observations were carried out between noon and 4 p.m. under room temperature (22–24 °C) and constant artificial light conditions (337.5 ± 4.95 lux during setting up the experiment, 599.0 ± 7.36 lux during observations under stereomicroscope). Lux was measured using Voltcraft BL-10 L Luxmeter.
The behavioural categories
To describe mite behaviour we used four previously documented activities22, 23, 26, 27, 56, 77,78,79,80,81:
-
Feeding: mite lies immobile on the leaf surface, feeding on plant tissues;
-
Walking: mite moves on the leaf surface;
-
Standing erect: mite stands on its anal lobes and moves the legs rapidly, or (much less commonly) mite lifts its hind body off of the leaf surface when standing on the legs (Fig. 1c);
-
Chain formation: three or more mites contact each other using their anal lobes, setae or legs. Mites were counted as individuals involved in each chain formation event (Fig. 1d).
Standing erect and chain formation have been explicitly interpreted as facilitating dispersal23,24,25,26.
Testing influence of potential dispersal cues
We expected that drying of the leaf fragment during the observation time would be an important background factor encouraging mites to initiate dispersal behaviour, and tested three other potential dispersal cues: (i) wind; (ii) presence of a potential insect vector; iii) presence of fresh plant material. As a control, mites were observed on leaf fragments with none of the other three potential dispersal.
Wind
Observations were carried out within a wind tunnel equipped with a fan (design modified from Melo et al.)33. The tunnel consisted of a transparent PVC (polyvinyl chloride) tube (∅ 4.5 cm, 20 cm length) with a free air inlet at one end and with the other end connected to an axial fan (AC Axial Fan DP202A212, Sunon) via a PVC tube. The fan was connected to a controller (AVT1813, AVT) that allowed for wind-speed regulation. At the distance of 1.5 cm from the free inlet, an open window (∅ 3.0 cm) at the top of the tube was present that allowed stereomicroscope observations. The experimental arena, consisting of the leaf fragment glued to the circle of adhesive tape on a square glass plate (12.25 cm2), was put in the tunnel under the open window. Wind speed in the tube was measured using a digital anemometer (Benetech model GM816) placed at the free end of the wind tunnel. The wind blew with a constant speed of 4.2 m/s (=15.12 km/h). This speed was chosen based on our preliminary observations showing that the optimal conditions inside the tunnel (those allowing mites to be dispersed but not forcing them to be blown) was between 3.5 and 4.5 m/s. We also followed previous research indicating that a speed around 4 m/s is suitable for effective eriophyid dispersal33, 56. Generally, it was difficult to calibrate whether the air currents inside the tunnel exactly matched that of atmospheric conditions. However, this constraint is present in all experimental set-ups using wind tunnels. Our equipment was built according to similar designs successfully used for studying wind dispersal in mites, including eriophyoids33, 55. Our replication of these conditions allows for comparability among studies.
Vector
One winged adult thrips (Anaphothrips obscurus) was transferred from the stock colony to the leaf fragment with the use of a small paintbrush and needle. During the experimental observations, thrips either walked on the leaf surface or they fed on the plant, occasionally changing feeding position. The thrips specimen was present on the leaf during the whole experiment.
Fresh plant material
A new clean wheat leaf cut into small fragments (1.5 cm length) and pressed on to the adhesive tape so that the pieces adhered were put near the leaf fragment with mite population on each side, with no gap between them.
Data Analysis
To investigate whether a specific behaviour was linked with one or more of the potential dispersal stimuli and whether the response differed between the studied species, a generalized linear model (GLM) was used with quasi-binomial distribution for proportions and the logit link function. The response variable was the proportion of mites performing a given behaviour during an experimental trial, and predictors were treatment (wind, vector, fresh plant material, control), species (CRM, WCM) and treatment x species interactions. For each behavioural category a separate model was built.
For each mite species separately, the contrasts between each treatment and respective control were compared, controlling the error rate by applying the method proposed by Hothorn et al.82. Statistical analyses were performed with R 3.3.183.
Data Availability
The datasets generated and analysed during the current study are available in the Zenodo repository [https://zenodo.org/record/437136] under: doi:10.5281/zenodo.437136.
References
Bowler, D. E. & Benton, T. G. Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. Biol. Rev. 80, 205–225 (2005).
Ronce, O. How does it feel to be like a rolling stone? Ten questions about dispersal evolution. Annu. Rev. Ecol. Evol. Syst. 38, 231–253 (2007).
Ochiai, K. & Susaki, K. Causes of natal dispersal in monogamous ungulate, the Japanese serow. Capricornis cripus. Anim. Behav. 73, 125–131 (2007).
Clobert, J., Baguette M., Benton T. G. & Bullock J. M. Dispersal ecology and evolution. (Oxford University Press, 2012).
Chust, G. et al. Dispersal similarly shapes both population genetics and community patterns in the marine realm. Sci. Rep. 6, 28730 (2016).
Matthysen, E. Multicausality of dispersal: a review in Dispersal Ecology and Evolution (eds Clobert, J., Baguette, M., Benton T. G. & Bullock, J. M.) 3–18 (Oxford University Press, 2012).
Yang, D. et al. Stepping-stones and dispersal flow: establishment of a meta-population of Milu (Elaphurus davidianus) through natural re-wilding. Sci. Rep. 6, 27297 (2016).
Reynolds, A. M. & Reynolds, D. R. Aphid aerial density profiles are consistent with turbulent advection amplifying flight behaviours: abandoning the epithet “passive”. Proc. R. Soc. B 276, 137–143 (2009).
Soons, M. B. et al. Directed dispersal by an abiotic vector: wetland plants disperse their seeds selectively to suitable sites along the hydrological gradient via water. Funct. Ecol. doi:10.1111/1365-2435.12771 (2016).
Bell, J. R., Bohan, D. A., Shaw, E. M. & Weyman, G. S. Ballooning dispersal using silk: world fauna, phylogenies, genetics and models. Bull. Entomol. Res. 95, 69–114 (2005).
Bonte, D., Bossuyt, B. & Lens, L. Aerial dispersal plasticity under different wind velocities in a salt marsh wolf spider. Behav. Ecol. 18, 438–443 (2007).
Greenstone, M. H. Meteorological determinants of spider ballooning: the roles of thermals vs. the vertical wind speed gradient in becoming airborne. Oecologia 84, 164–168 (1990).
Washburn, J. O. & Washburn, L. Active aerial dispersal of minute wingless arthropods: exploitation of boundary-layer velocity gradients. Science 223, 1088–1089 (1984).
Johnson, D. T. & Croft, B. A. Laboratory study of the dispersal behavior of Amblyseius fallacis (Acarina: Phytoseiidae). Ann. Entomol. Soc. Am. 69, 1019–1023 (1976).
Hoy, M. A. Aerial dispersal and field efficacy of a genetically improved strain of the spider mite predator Metaseiulus occidentalis. Entomol. Exp. Appl. 32, 205–212 (1982).
Croft, B. A. & Jung, C. Phytoseiid dispersal at plant to regional levels: a review with emphasis on management of Neoseiulus fallacis in diverse agroecosystems. Exp. Appl. Acarol. 25, 763–784 (2001).
Smitley, D. R. & Kennedy, G. G. Photo-oriented aerial-dispersal behavior of Tetranychus urticae (Acari: Tetranychidae) enhances escape from the leaf surface. Ann. Entomol. Soc. Am. 78, 609–614 (1985).
Margolies, D. C. Conditions eliciting aerial dispersal behavior banks grass mite, Oligonychus pratensis (Acari: Tetranychidae). Environ. Entomol. 16, 928–932 (1987).
Li, J. & Margolies, D. C. Quantitative genetics of aerial dispersal behavior and life-history traits in Tetranychus urticae. Heredity 70, 544–552 (1993).
Clotuche, G. et al. The formation of collective silk balls in the spider mite Tetranychus urticae Koch. PLoS One 6(4), e18854 (2011).
Clotuche, G., Navajas, M., Mailleux, A. C. & Hance, T. Reaching the ball or missing the flight? Collective dispersal in the two-spotted spider mite Tetranychus urticae. PLoS One 8(10), e77573 (2013).
Michalska, K., Skoracka, A., Navia, D. & Amrine, J. W. Behavioural studies on eriophyoid mites: an overview. Exp. Appl. Acarol. 51, 31–59 (2010).
Sabelis, M. W. & Bruin, J. Evolutionary ecology: Life history patterns, food plant choice and dispersal in Eriophyoid Mites: Their Biology, Natural Enemies and Control (eds Lindquist, E. E., Sabelis, M. W. & Bruin J.) 329–365 (Elsevier Science Publishers, 1996).
Gibson, W. W. & Painter, R. H. Transportation by aphids of the wheat curl mite, Aceria tulipae (K.), a vector of the Wheat Streak Mosaic Virus. J. Kans. Entomol. Soc. 30, 147–153 (1957).
Duffner, K., Schruft, G. & Guggenheim, R. Passive dispersal of the grape rust mite Calepitrimerus vitis Nalepa 1905 (Acari, Eriophyoidea) in vineyards. J. Pest. Sci. 74, 1–6 (2001).
Ozman, S. K. & Goolsby, J. A. Biology and phenology of the eriophyid mite, Floracarus perrepae, on its native host in Australia, Old World climbing fern, Lygodium microphyllum. Exp. Appl. Acarol. 35, 197–213 (2005).
Skoracka, A., Kuczyński, L. & Rector, B. G. Divergent host acceptance behavior suggests host specialization in populations of the polyphagous mite Abacarus hystrix (Acari: Prostigmata: Eriophyidae). Environ. Entomol. 36, 899–909 (2007).
Craig, C. L. Evolution of arthropod silks. Annu. Rev. Entomol. 42, 231–267 (1997).
Cheung, S. G., Luk, K. C. & Shin, P. K. S. Predator-labeling effect on byssus production in marine mussels Perna viridis (L.) and Brachidontes variabilis (Krauss). J. Chem. Ecol. 32, 1501–1512 (2006).
Young, O. P. & Lockley, T. C. Dragonfly predation upon Phidippus audax (Araneae, Salticidae). J. Arachnol. 16, 121–122 (1988).
Bonte, D. et al. Costs of dispersal. Biol. Rev. Camb. Philos. Soc. 87, 290–312 (2012).
Osakabe, M. et al. Aerodynamic advantages of upside down take-off for aerial dispersal in Tetranychus spider mites. Exp. Appl. Acarol. 44, 165–183 (2008).
Melo, J. W. S., Lima, D. B., Sabelis, M. W., Pallini, A. & Gondim, M. G. C. Behaviour of coconut mites preceding take-off to passive aerial dispersal. Exp. Appl. Acarol. 64, 429–443 (2014).
Lindquist, E. E. & Oldfield, G. N. Evolution of eriophyoid mites in relation to their host plants in Eriophyoid Mites: Their Biology, Natural Enemies and Control (eds Lindquist, E. E., Sabelis, M. W. & Bruin J.) 277–300 (Elsevier Science Publishers, 1996).
Skoracka, A. & Dabert, M. The cereal rust mite Abacarus hystrix (Acari: Eriophyoidea) is a complex of species: evidence from mitochondrial and nuclear DNA sequences. Bull. Entomol. Res. 100, 263–272 (2010).
Skoracka, A. et al. Cryptic species within the wheat curl mite Aceria tosichella (Keifer) (Acari: Eriophyoidea), revealed by mitochondrial, nuclear and morphometric data. Invertebr. Syst. 26, 417–433 (2012).
Gillespie, R. G. et al. Long-distance dispersal: A framework for hypothesis testing. Trends Ecol. Evol. 27, 47–55 (2012).
Costa, D. et al. Genetic structure of soil invertebrate populations: Collembolans, earthworms and isopods. Appl. Soil Ecol. 68, 61–66 (2013).
Massee, A. M. The Life-history of the Black Currant Gall Mite, Eriophyes ribis (Westw.). Nat. Bull. Entomol. Res. 18, 277–307 (1928).
Slykhuis, J. T. Aceria tulipae Keifer (Acarina: Eriophyidae) in relation to the spread of wheat streak mosaic. Phytopathology 45, 116–128 (1955).
Shvanderov, F. A. Role of phoresy in dispersal of four- legged mites. Zool. Zh. 54, 458–461 (in Russian) (1975).
Waite, G. K. & McAlpine, J. D. Honey bees as carriers of lychee erinose mite Eriophyes litchii (Acari: Eriophyiidae). Exp. Appl. Acarol. 15, 299–302 (1992).
Liu, S. et al. Seasonal phoresy as an overwintering strategy of a phytophagous mite. Sci. Rep. 6, 25483 (2016).
Galvão, A. S. et al. Dispersal strategies of Aceria guerreronis (Acari: Eriophyidae), a coconut pest. Exp. Appl. Acarol. 57, 1–13 (2012).
Brandenburg, R. L. & Kennedy, G. G. Intercrop relationships and spider mite dispersal in a corn/peanut agro-ecosystem. Entomol. Exp. Appl. 32, 269–276 (1982).
Hoy, M. A., van de Baan, H. E., Groot, J. J. R. & Field, R. P. Aerial movements of mites in almonds: implications for pest management. Calif. Agric. 38, 21–23 (1984).
Weinstein, R. B. Effects of temperature and water loss on terrestrial locomotor performance in land crabs: integrating laboratory and field studies. Amer. Zool. 38, 518–527 (1998).
Capinha, C. & Anastacio, P. Assessing the environmental requirements of invaders using ensembles of distribution models. Divers. Distrib. 17, 13–24 (2011).
Capinha, C., Brotons, L. & Anastacio, P. Geographical variability in propagule pressure and climatic suitability explain the European distribution of two highly invasive crayfish. J. Biogeogr. 40, 548–558 (2011).
Fialho, C., Banha, F. & Anastácio, P. M. Factors determining active dispersal capacity of adult chinese mitten crab Eriocheir sinensis (decapoda, varunidae). Hydrobiologia 767, 321–331 (2016).
Richter, C. J. J. Aerial dispersal in relation to habitat in eight wolf spider species (Pardosa, Araneae, Lycosidae). Oecologia 5, 200–214 (1970).
Suter, R. B. An aerial lottery: the physics of ballooning in a chaotic atmosphere. J. Arachnol. 27, 281–293 (1999).
Reynolds, A. M., Bohan, D. A. & Bell, J. R. Ballooning dispersal in arthropod taxa: conditions at take-off. Biol. Lett. 3, 237–240 (2007).
Vugts, H. F. & van Wingerden, W. K. R. E. Meteorological aspects of aeronautic behaviour in spiders. Oikos 27, 433–444 (1976).
Sabelis, M. W. & Afman, D. P. Synomone-induced suppression of take-off in the phytoseiid mite Phytoseiulus persimilis Athias-Henriot. Exp. Appl. Acarol. 18, 711–721 (1994).
Smith, B. D. The behaviour of the black currant gall mite (Phytoptus ribis Nal.) during the free living phase of its life cycle. Ann. Rep. Agric. hort. Res. Sta., Long Ashton, Bristol 13, 130–136 (1960).
Pedgley, D. E. Windborne pests and diseases: meteorology of airborne organisms. (Ellis Horwood, 1982).
Dingle, H. Migration: the biology of life on the move. (Oxford University Press, 1996).
Reynolds, A. M. Beating the odds in the aerial lottery: passive dispersers select conditions at take-off that maximise their expected fitness on landing. Am. Nat. 181, 555–561 (2013).
Swingland, I. R. Intraspecific differences in movement in The Ecology of Animal Movement (eds Swingland I. R. & Greewood P. J.) 102–115 (Clarendon Press, 1983).
Glida, H., Bertrand, M. & Peyrusse, V. A limiting factor in the abundance of predatory phoretic mites (Acari: Macrochelidae): the seasonal abundance of their phorionts (dung beetles) in southern France. Can. J. Zool. 81, 2066–2072 (2003).
Aragón, P., Meylan, S. & Clobert, J. Dispersal status-dependent response to the social environment in the Common Lizard, Lacerta vivipara. Funct. Ecol. 20, 900–907 (2006).
Clobert, J., Le Galliard, J. F., Cote, J., Meylan, S. & Massot, M. Informed dispersal, heterogeneity in animal dispersal syndromes and the dynamics of spatially structured populations. Ecol. Lett. 12, 197–209 (2009).
Chapple, D. G., Simmonds, S. M. & Wong, B. B. M. Can behavioral and personality traits influence the success of unintentional species introductions? Trends Ecol. Evol. 27, 57–64 (2012).
Marty, P. R., Hodges, K., Agil, M. & Engelhardt, A. Determinants of immigration strategies in male crested macaques (Macaca nigra). Sci. Rep. 6, 32028 (2016).
Weiß, B. M., Kulik, L., Ruiz-Lambides, A. V. & Widdig, A. Individual dispersal decisions affect fitness via maternal rank effects in male rhesus macaques. Sci. Rep. 6, 32212 (2016).
Cote, J., Clobert, J., Brodin, T., Fogarty, S. & Sih, A. Personality-dependent dispersal: characterization, ontogeny and consequences for spatially structured populations. Phil. Trans. R. Soc. B 365, 4065–4076 (2010).
Nault, L. R. & Styer, W. E. The dispersal of Aceria tulipae and three other grass-infesting eriophyid mites in Ohio. Ann. Entomol. Soc. Am. 62, 1446–1455 (1969).
Navia, D. et al. Wheat curl mite, Aceria tosichella, and transmitted viruses: An expanding pest complex affecting cereal crops. Exp. Appl. Acarol. 59, 95–143 (2013).
Skoracka, A. et al. Global spread of wheat curl mite by its most polyphagous and pestiferous lineages. Ann. Entomol. Soc. Am. 165, 222–235 (2014).
Rehage, J. S. & Sih, A. Dispersal behavior, boldness, and the link to invasiveness: a comparison of four Gambusia species. Biol. Invasions 6, 379–391 (2004).
Skoracka, A., Kuczyński, L., Szydło, W. & Rector, B. The wheat curl mite Aceria tosichella (Acari: Eriophyoidea) is a complex of cryptic lineages with divergent host ranges: evidence from molecular and plant bioassay data. Biol. J. Linn. Soc. 109, 165–180 (2013).
Dabert, J., Ehrnsberger, R. & Dabert, M. Glaucalges tytonis sp. n. (Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa 1719, 41–52 (2008).
Tamura, K. et al. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739 (2011).
Zawirska, I. Thysanoptera in Diagnosis of Plant Pests and Their Natural Enemies (ed. Boczek, J.), pp.145–174 (in Polish) (Wydawnictwo SGGW, 1994).
Mirab-Balou, M., Minaei, K. & Chen, X. X. An illustrated key to the genera of Thripinae (Thysanoptera, Thripidae) from Iran. Zookeys 317, 27–52 (2013).
Krantz, G. W. Observations on the morphology and behaviour of the filbert rust mite Aculus comatus (Prostigmata: Eriophyoidea) in Oregon. Ann. Entomol. Soc. Am. 66, 709–717 (1973).
Gibson, R. W. Studies on the feeding behaviour of the eriophyid mite Abacarus hystrix, a vector of grass viruses. Ann. Appl. Biol. 78, 213–217 (1974).
McCoy, C. W. & Albrigo, L. G. Feeding injury to the orange caused by the citrus rust mite, Phyllocoptruta oleivora (Prostigmata: Eriophyoidea). Ann. Entomol. Soc. Am. 68, 289–297 (1975).
Nuzzaci, G. Behaviour of eriophyoid mites during the uptake of food. Entomologica, Bari 12, 75–80 (in Italian) (1976).
Westphal, E. & Manson, D. C. M. Feeding effects on host plants: gall formation and other distortions in Eriophyoid Mites: Their Biology, Natural Enemies and Control (eds Lindquist, E. E., Sabelis, M. W. & Bruin J.) 231–242 (Elsevier Science Publishers, 1996).
Hothorn, T., Bretz, F. & Westfall, P. Simultaneous inference in general parametric models. Biom. J. 50, 346–363 (2008).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing https://www.R-project.org/ (2016).
Acknowledgements
We gratefully acknowledge Justyna Pińkowska for help in performing molecular work, Wiktoria Szydło for sharing stock colonies of WCM mites, Alicja Laska for information about the dispersal of eriophyoids in different wind speeds, Prof. Wojciech Kuczyński for valuable discussion on the physics of wind tunnels and Hanna Legutowska (WULS-SGGW) for help in thrips identification. The study was partially supported by a grant from the Dean of the Faculty of Biology, AMU (GDWB-05/2016). This work was supported by the KNOW RNA Research Centre in Poznań (No. 01/KNOW2/2014).
Author information
Authors and Affiliations
Contributions
A.K., A.S. and L.K. conceived and conceptualized the research. A.K., A.S., L.K., M.L. designed the methodology. A.K. performed behavioural observations and prepared data for the analysis; L.K. performed statistical analyses. A.S., A.K. provided resources. A.K., A.S., L.K., M.L. and H.P. discussed the results and interpretation, drafted the manuscript, revised the manuscript critically for important intellectual content and gave final approval for publication. The manuscript is part of Agnieszka Kiedrowicz’s PhD thesis.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kiedrowicz, A., Kuczyński, L., Lewandowski, M. et al. Behavioural responses to potential dispersal cues in two economically important species of cereal-feeding eriophyid mites. Sci Rep 7, 3890 (2017). https://doi.org/10.1038/s41598-017-04372-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-04372-7
This article is cited by
-
Effects of temperature on the movement and feeding behaviour of the large lupine beetle, Sitona gressorius
Journal of Pest Science (2023)
-
A rotatory funnel-shaped collector for trapping airborne mites in a glycerin-based adhesive surface
Experimental and Applied Acarology (2022)
-
Hitchhiking or hang gliding? Dispersal strategies of two cereal-feeding eriophyoid mite species
Experimental and Applied Acarology (2021)
-
Temperature-dependent development and survival of an invasive genotype of wheat curl mite, Aceria tosichella
Experimental and Applied Acarology (2021)
-
A comprehensive and cost-effective approach for investigating passive dispersal in minute invertebrates with case studies of phytophagous eriophyid mites
Experimental and Applied Acarology (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
