
Abstract
The continued success of malaria control efforts requires the development, study and implementation of new technologies that circumvent insecticide resistance. We previously demonstrated that fungal pathogens can provide an effective delivery system for mosquitocidal or malariacidal biomolecules. Here we compared genes from arthropod predators encoding insect specific sodium, potassium and calcium channel blockers for their ability to improve the efficacy of Metarhizium against wild-caught, insecticide-resistant anophelines. Toxins expressed under control of a hemolymph-specific promoter increased fungal lethality to mosquitoes at spore dosages as low as one conidium per mosquito. One of the most potent, the EPA approved Hybrid (Ca++/K+ channel blocker), was studied for pre-lethal effects. These included reduced blood feeding behavior, with almost 100% of insects infected with ~6 spores unable to transmit malaria within 5 days post-infection, surpassing the World Health Organization threshold for successful vector control agents. Furthermore, recombinant strains co-expressing Hybrid toxin and AaIT (Na+ channel blocker) produced synergistic effects, requiring 45% fewer spores to kill half of the mosquitoes in 5 days as single toxin strains. Our results identify a repertoire of toxins with different modes of action that improve the utility of entomopathogens as a technology that is compatible with existing insecticide-based control methods.
Similar content being viewed by others

Introduction
Some of the most important human diseases are borne by mosquitoes including malaria (Anopheles), filariasis (Culex, Mansonia and some Anopheles spp.), viral encephalitides (Culex), dengue and yellow fever (principally Aedes aegypti). An estimated 2 billion people live in areas where these diseases are endemic. The burden is heaviest in sub-Saharan Africa, where ~200 million cases of malaria are reported annually and many children succumb1. Emergence and spread of resistance to pyrethroids, organophosphates and carbamates is a particular threat, as most disease control programs rely heavily on these broad-spectrum chemical insecticides to reduce vector populations2,3,4,5. In recent years, anopheline malaria vectors in sub-Saharan Africa have gained sufficient resistance to render chemical insecticides largely ineffective5, 6.
The chemical insecticides used in mosquito control programs are directed at neuronal voltage-gated sodium (NaV) channels, as are many insect-specific neurotoxins derived from arthropods, including the scorpion toxin AaIT that has been approved for release in several countries7. A recombinant strain of the entomopathogenic fungus Metarhizium anisopliae expressing AaIT killed Ae. aegypti mosquitoes faster and at lower spore doses than wild-type (WT) fungus8. Since AaIT and pyrethroids bind to different sites on insect NaV channels and channel mutations that confer resistance to pyrethroids actually increase binding of AaIT, the use of both pyrethroids and AaIT should mitigate against resistance acquisition9. Furthermore, combining toxins with different targets, such as voltage-gated sodium, potassium (KV) and calcium (CaV) channels, could provide useful additive or synergistic effects and minimize the potential for cross-resistance. CaV and KV channels are previously unexploited insecticide targets, reducing the likelihood of pre-existing resistance.
We compared the efficacy against Anopheles gambiae of recombinant Metarhizium pingshaense co-expressing green fluorescent protein (GFP) and one of several arthropod-derived toxins with different modes of action: AaIT, the NaV channel inhibitor μ-CUTX-As1a (As1a)10, the KCa/CaV blocker ω/κ-hexatoxin-Hv1a (Hybrid), the CaV channel blocker ω-hexatoxin-Hv1a (Hv1a)11, and U1-AGTX-Ta1a (Ta1a), a weaponized insect hormone which has an unknown neuronal target12. With the exception of AaIT, all these toxins are derived from spider venom. To express the transgenes we used the Mcl1 promoter that targets expression to the hemocoel of infected insects8, 13, 14.
We previously determined that hyphal bodies appear in the hemolymph 2–3 days post infection and that Mcl1 expression commences within 20 min of contact with hemolymph15.
Transformants were selected by GFP expression and PCR/sequencing confirmation of the toxin gene, and initially bioassayed by spraying spore suspensions (~1 conidium/mosquito; ~3 conidia/mosquito; and ~6 conidia/mosquito) onto cold (4 °C) anesthetized adult female An. gambiae. Although they have different molecular targets (NaV, CaV and KCa channels), each toxin significantly (p < 0.05) improved the median lethal time (LT50) with infection loads of 3 or more spores (Fig. 1a,b, and Table 1) compared to WT, and therefore they constitute a potential arsenal that could be rotated and/or used in combination in a mosquito control program aimed at mitigating resistance. Hybrid, also known as VersitudeTM, was chosen for further study as the US EPA has already approved it for use as a stand-alone insect control agent. Since prevention of transmission of malaria is of primary importance in assessing a mosquito control technology, we took a holistic view of the life cycle of the parasite to determine if pre-lethal effects could contribute to Metarhizium preventing the spread of malaria.

(a) LT50 values for mosquitoes treated with 1 × 105, 1 × 106 and 1 × 107 M. pingshaense conidia/ml suspensions resulting in ~1, ~3 and ~6 conidia/mosquito, respectively, of M. pingshaense WT or M. pingshaense expressing Hybrid, AaIT or both AaIT and Hybrid. LC50 values are reported along the x-axis as the inverse of the estimated spore inoculum. Lettering represents statistical differences (p < 0.05) based on a log-rank test comparing the Kaplan-Meier survival curves. The LC50 dose for untreated mosquitoes was fixed at zero and reported for all spore concentrations for comparison. (b) Schematic representing infection timing (beige) and LT50s of mosquitoes treated with conidial suspensions and exposed to cloth impregnated with toxin expressing strains or WT. Lettering groups toxins by statistical significance (p < 0.05) based on a log-rank test comparing the Kaplan-Meier survival curves.
We tested fungal efficacy against three anopheline vectors of human malaria: two wild-caught, insecticide-resistant species (An. coluzzii and An. gambiae s.s.) and one lab-reared, insecticide-susceptible species (An. gambiae Kisumu An. kisumu). We applied the fungus to sheets, as fungal-impregnated sheets hung in houses provide a resting area for mosquitoes that have taken blood meals16. Most malaria vectors prefer to blood feed and rest inside houses, thus maximizing the likelihood of fungal contact and infection17. We exposed mosquito populations for one hour to sheets impregnated with either: 1) Met-RFP, a strain with WT virulence expressing red fluorescent protein (RFP) as a marker, and 2) Hybrid toxin expressing fungus (Met-Hybrid) co-expressing GFP. We found mosquitoes exposed to the cloth for one hour in a WHO mosquito bioassay tube picked up an average of 129 ± 18 spores, a sufficient dose to kill all mosquitoes exposed to each fungal treatment. Mortality was monitored over 14 days and compared to uninfected controls.
Insecticide (Knockdown) resistance (kdr) in each mosquito population was quantified using PCR. The kdr mutation reduces sensitivity to DDT and pyrethroids and is the most prevalent form of insecticide resistance for West African mosquitoes6. The levels of kdr resistance in wild-caught An. coluzzii and An. gambiae s.s. were 98.3% and 92.9%, respectively (Supplementary Table 1). An. gambiae Kisumu mosquitoes are an established laboratory population of mosquitoes with stable susceptibility to insecticides, so their resistance was not tested.
Overall, insecticide resistance did not alter the susceptibility of the three mosquito species to Met-RFP and Met-Hybrid (Fig. 2). Within 2.5 days post-infection, mosquitoes exposed to Met-Hybrid were dying faster than those exposed to Met-RFP. LT50 (LT80) values for Met-Hybrid and Met-RFP were 4.14 ± 0.16 (5.47 ± 0.25) and 6.18 ± 0.14 (7.71 ± 0.16) days, respectively (mean ± standard error is reported). Fluorescent Metarhizium mycosis was observed on fungus-exposed cadavers, confirming mortality due to treatment. The number of mosquitoes surviving in the uninfected control group never dropped below 84.4% (Fig. 2).

Survival following WHO tube exposure: these three graphs represent survival curves for RFP expressing Metarhizium (Met-RFP) and Hybrid toxin-expressing Metarhizium (Met-Hybrid) against two wild-caught, insecticide-resistant (An. coluzzii and An. gambiae s.s.) and one lab-reared (An. gambiae Kisumu) human malaria vector mosquitoes. There are no significant differences in time to kill insecticide resistant and susceptible strains, but Met-Hybrid is more effective on all mosquito strains.
The wild-type M. pingshaense strain has a narrow host range. To investigate if expressing Hybrid toxin impacts host specificity, we assayed transformants against honeybees (Apis mellifera adansonii) as a representative local pollinator. Honeybees were infected with black cloth impregnated with ~2 × 106 conidia/cm2 in WHO tubes and spore dose was estimated using our selective media protocol. Met-RFP and Met-Hybrid did not kill any honeybees in the two weeks they were observed following infection (there was no difference in mortality rates between fungal exposed and unexposed bees, and no fungal emergence was observed on any bee cadaver). This is in agreement with previous studies indicating that the expression of insect toxins did not increase the host range of M. acridum 13. Interestingly, honeybees picked up significantly fewer spores (80 ± 3 spores, p < 0.05) than mosquitoes under the same conditions, despite the much smaller size of mosquitoes. Spraying bees with 1 × 108 spores/ml also failed to cause mortality.
Willingness and ability to blood feed were also tested, as even small changes in coordination could potentially interfere with behavior and disease transmission. Host-seeking (blood feeding) interest was quantified as the percentage of the mosquito population choosing the chamber closest to the host in a guinea pig choice chamber (Fig. 3a,b and Supplementary Figure 1a,b). At one day post-infection, 94.3% of untreated (controls) and treated (Met-RFP or Met-Hybrid) mosquitoes flew toward the blood source with no significant differences between treatments. The willingness of mosquitoes in the control group to blood feed did not change over the course of the experiment. In contrast, significantly (p < 0.05) fewer (56.9%) mosquitoes treated with Met-Hybrid flew to the blood source on day 3 as compared to Met-RFP (82.1%). By day 4 the number of Met-Hybrid infected mosquitoes in the guinea pig choice chamber (38.9%) was not significantly different than the 30% entering the chamber in the absence of a guinea pig. Met-RFP infected mosquitoes only became significantly (p < 0.01) less responsive than uninfected controls on day 4 (Fig. 3a). These results suggest a pre-lethal advantage to using transgenic fungi for mosquito control.

(a) Impact of fungal infection on blood-feeding at 1–5 days post-infection with either Met-Hybrid or Met-RFP. Mosquitoes were placed in a choice chamber with the guinea pig host outside of the chamber and just out of reach (Supplementary Figure 1a,b). Host-seeking (blood feeding) interest was quantified as the percentage of the mosquito population choosing the chamber closest to the host. The symbol “*” denotes no significant differences in mosquito choices with or without a host: 30 ± 3.05% of the mosquitoes chose the “host” chamber even in the absence of a host. (b) Mortality and transmission of mosquitoes exposed to fungus, the light area represents the percent survival of mosquitoes for each treatment, while the dark area shows the proportion of mosquitoes in each treatment that are alive and would seek a host to blood feed (capable of malaria transmission). The upper dashed line represents the LT50 while the lower dashed line represents the 80% control threshold suggested by the World Health Organization (WHO) for a successful vector control agent18.
With this information, we projected the measured proportion of mosquitoes interested in blood feeding onto the mortality of mosquitoes to identify the fraction capable of malaria transmission (Fig. 3b). In contrast to Met-RFP, by day 5, Met-Hybrid infected mosquitoes passed the threshold in both metrics (8.35% malaria transmission and >80% mortality) for the 80% control threshold suggested by the World Health Organization (WHO) for a successful vector control agent18.
Like Hybrid, AaIT is US EPA approved. As AaIT targets NaV channels and the Hybrid toxin targets CaV and KCa channels, we examined whether their different sites of action produce synergistic effects. We found a clear benefit in terms of both effective spore doses and speed of kill in expressing both Hybrid and AaIT in a single strain (Met-Hybrid/AaIT) (Table 1). Mosquitoes sprayed with 1 × 105 spores/mL (~1 spore/mosquito) of Met-Hybrid/AaIT, Met-Hybrid or WT had LT50s of 8.90 ± 1.09, 9.08 ± 0.44, and 9.49 ± 0.60 days, respectively, and the differences between them fell short of significance (Fig. 1a and Table 1). At this low spore dose, Met-Hybrid/AaIT, Met-Hybrid and WT significantly (p < 0.05) reduced lifespan compared to untreated mosquitoes (LT50 of 11.4 ± 0.52 days), and high variation in Met-AaIT bioassays resulted in no statistical difference from untreated mosquitoes. However, increasing the spore doses reduced the LT50s of toxin expressing strains to a greater extent than WT, consistent with the toxins having threshold effects (Fig. 1a,b). Comparing toxins, mosquitoes sprayed with 1 × 106 Met-Hybrid/AaIT spores/ml (~3 spores/mosquito) achieved an LT50 of 5.30 ± 0.42 days that was significantly faster than the 6.39 ± 0.35 and 7.20 ± 0.84 days for Met-Hybrid and Met-AaIT alone. The Tabashnik synergism equation19 confirmed synergistic interactions as early as 4.5 days post-infection, and at 5 days it takes less than half the dosage of Met-Hybrid/AaIT to kill mosquitoes at the same rate as either Met-Hybrid or Met-AaIT applied alone (Supplementary Table 2). Such synergies suggest that optimizing the overall efficacy of the control strategy will require multiple transgenes, and a toxin arsenal could reduce effective conidial doses thereby reducing end-user costs (Fig. 1a and Table 1).
The spray infection method successfully probes efficacy differences between transgenes by delivering low spore doses, but contributes to variation in LT50 values because some mosquitoes escape infection at these low dosages. We estimated that inocula containing 1 × 107, 1 × 106 and 1 × 105 spores/ml deliver at least one spore to 95%, 75%, and 50% of the mosquitoes sprayed, respectively. As mosquito mortality increases with inoculum load, we plotted spore counts per mosquito versus the probability of death derived from our bioassays for the WT and transgenic strains (Met-AaIT, Met-Hybrid, and Met-Hybrid/AaIT) (Fig. 4). Together, these data revealed that the lethal dose per mosquito at which 100% of mosquitoes die (LD100) is 10 spores, 8 spores, 7 spores, and 6 spores for WT, Met-AaIT, Met-Hybrid and Met-Hybrid/AaIT, respectively. The results explain the incomplete mortality seen even with our highest spore concentration (1 × 107), and the complete mortality seen in mosquitoes treated through contact with oil-impregnated cloth, which delivers far more spores to every mosquito. This bodes well for oil-impregnated cloth as a delivery system for entomopathogenic fungal spores and for the translatability of spray method results to the field.

The number of spores infecting each mosquito after spraying with three different spore concentrations (1 × 105, 1 × 106, and 1 × 107 spores/mL in 0.01% Tween80) plotted against the probability of death. The mean number of spores delivered with each suspension is marked with a blue crossbar. Assuming that mosquitoes with a higher dose are more likely to die, we calculated the chance of death for each mosquito based on our survival data for each treatment at each concentration. Mosquitoes with an estimated 100% chance of death are colored in red, and 0–99% is represented with a green to orange gradient. The red dashed line indicates the estimated LD100 in each treatment (10 spores for WT, 8 spores for AaIT, 7 spores for Hybrid and 6 spores for Hybrid/AaIT).
In conclusion, fungi can be genetically modified to strategically maximize their success as biocontrol agents13, 20. When their impact on malaria transmission is considered, transgenic fungi applied on sheets meet WHO standards for effective control of malaria within 5 days post-exposure, indicating that the inclusion of transgenic Metarhizium in pre-existing control efforts would effectively decrease malaria transmission. Some female mosquitoes infected with transgenic fungi will lay eggs 3–4 days after a bloodmeal, so infected mosquitoes may still pass their genes onto the next generation, but infection will prevent further oviposition. Our study emphasizes the need to consider the effect of fungi on blood feeding for modeling of existing mosquito control techniques in conjunction with transgenic Metarhizium 21.
Methods
Ethics statement
Ethical permissions were obtained through the Institutional Review of Institut de Recherche en Science de la Santé (IRSS) and Centre Muraz ethics committee. In addition, prior authorization was granted from the Burkina Faso National Biosecurity Agency for importing and using Metarhizium transgenic fungi expressing Hybrid toxin for semi-field and lab work (Supplementary File). All experiments with guinea pigs were carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. In addition, the protocols followed the IRSS Animal Welfare Assurance A5926-01. Trained personnel and veterinarians cared for all animals involved in this study.
Mosquito colony
The study was carried out in a joint IRSS and Centre Muraz laboratory in Bobo Dioulasso, Burkina Faso. For bioassays, we used adult mosquitoes An. coluzzii and An. gambiae reared from larval collections at Vallée du Kou (11°23′N, 4°24′W) and Soumousso (11°04′N, 4°03′W), respectively. Mosquitoes from these areas are resistant to multiple insecticides6. The laboratory-reared Kisumu strain of Anopheles gambiae s.s. was used as an insecticide-susceptible reference strain. All larvae were simultaneously reared under standard, controlled conditions (26 ± 2 °C, 80 ± 10% relative humidity (RH) and 12:12 light-dark cycle). Larvae were fed daily with Tetramin™. Pupae were collected and transferred into holding cages measuring 30 cm × 30 cm × 30 cm covered with mosquito netting. Upon emergence, mosquitoes were immediately morphologically identified and sexed using a key and were maintained in cages on 6% sugar solution for 2–3 days prior to fungal infection. Only two to three day old females were used for bioassays with fungi.
Honeybee rearing
Honeybees, a locally reared Apis mellifera adansonii Latreille 1804, were kept in groups of 25 to 30 in cages in an insectarium at 25.3 ± 1 °C, and fed on 10% sterile glucose with syrup of honey and sterile tap water.
Fungal strains and fungal transformation
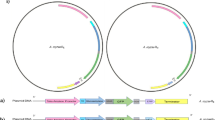
The African isolate of Metarhizium pingshaense used in this study was maintained on potato dextrose agar at 26 °C and 70% RH. This M. pingshaense strain was chosen because it has a narrow host range and does not infect local Burkina Faso insects Apis mellifera adansonii (honeybees) and Schistocera gregaria (locusts). Toxin genes were chemically synthesized with the MCL1 signal peptide to ensure secretion, and they were cloned in a common transfer plasmid pPK2-Bar-GFP22, downstream of the Mcl1 promoter as described13, to target high-level production of each toxin into the hemolymph. To produce the dual toxin expressing strain Met-Hybrid/AaIT, we used the plasmid pAaIT1 expressing AaIT1 described previously13. To construct the Ti plasmid expressing Hybrid, the cassette that contains the hybrid encoding sequence downstream of the Mcl1 was excised from the plasmid pMcl1-Hybird13 with EcoRI and NotI, treated with T4 DNA polymerase, and inserted into the EcoRV site of pPK2-Sur-GFP22 to form the Ti plasmid. The pBar-Hybrid and pSur-Hybrid plasmids were consecutively transformed into M. pingshaense using Agrobacterium 13 as described15 with Bar and Sur genes, respectively, as selection markers. The Ti plasmid also contains the gene for green fluorescent protein (GFP) under a constitutive promoter (GdpA). An RFP-expressing strain was produced as described23 and confirmed to show WT virulence and growth against Anopheles. The presence of each toxin gene in the Metarhizium genome was confirmed by PCR. PCR products from each of the transformants were electrophoresed on agarose gel, excised and sequenced to confirm the presence of transgenes in transformed Metarhizium strains. The primers used in this study and their respective usages are summarized in Supplementary Table 3.
Fungal formulations and infection of insects in World Health Organization tubes
Fungal strains were harvested after 2 weeks of incubation at 26 °C. Five ml of sesame oil formulation at 8.0 × 107 conidia ml−1 were used to impregnate pieces of 12 × 15 cm black cotton sheets. This produced a final density of ~2 × 106 conidia/cm2. Control treatments were oil preparations without fungal spores. Impregnated sheets were dried for 16 h overnight at ambient temperature (26 ± 1 °C, 80 ± 10% RH) before placing inside WHO bioassay tubes (15 cm long, 4 cm diameter; Supplementary Figure 2). 25–30 mosquitoes or bees were exposed in the WHO tube for 1 h. After exposure, each group was transferred into 20 cl plastic cups for twice-daily observations of mortality and, in the case of the mosquitoes, blood feeding choice experiments (Supplementary Figure 3).
Mortality was counted twice daily. For any deaths, cadavers were surface sterilized and placed on 1.5% agar plates to promote fungal emergence and confirm if cause of death was fungal. Mortality caused by other factors was not included in survival analysis.
Fungal growth observations
Mosquito mortality was measured twice daily. Cadavers were immediately removed from their cups and each was washed once for 20 s with 1% sodium hypochlorite for 20 s and twice with sterile distilled water for 40 s. Washed cadavers were then placed on a 1.5% agar plate. Fungal growth on mosquitoes was confirmed microscopically after 3–5 days incubation.
PCR assay of insecticide resistance
During mortality bioassays, two randomly chosen legs from each mosquito were dissected. This subsample was then analyzed by PCR to definitively identify the species, molecular form24 and the kdr mutation levels25.
Dual transgene fungal infections
Comparative transgene experiments were conducted at the University of Maryland as described15. Briefly, in three replicates, 30 female mosquitoes (An. gambiae Keele) were cold anesthetized (4 °C) and sprayed 8 times with an atomizer containing a conidial suspension in 0.01% Tween-80. Treated mosquitoes were checked daily for mortality for 11 days.
Blood feeding choice tunnel
A 60-cm long, glass choice tunnel (25 cm × 25 cm area) was used for blood feeding assays (Supplementary Figure 1a,b). A 25-cm square of polyester netting was fitted at one end of the tunnel as a compartment. A netted barrier was placed one-third along the length of the glass tunnel separating the tubes into short and long sections. The barrier was 400 cm2 (20 × 20 cm), with nine 1 cm diameter holes for passage; one hole was located at the center of the square and the other eight were equidistantly located 5 cm from the border. This choice chamber is designed as a miniaturized proxy for a traditional West African home. The largest section of such a home is the veranda that serves as a sitting area. This corresponds to the first compartment of the tunnel (40 cm long). The second smaller part of a traditional house is the bedroom where residents sleep under bed netting. This corresponds to the smaller compartment of the tunnel (20 cm long). We placed the guinea pig within 5 cm of this compartment to represent a sleeping occupant at night (Supplementary Figure 1a,b). Fifty non-blood-fed female mosquitoes (An. coluzzii) were released into the long section of the tunnel. In this design, female mosquitoes are normally attracted through the barrier into the smaller compartment by the guinea pig to blood feed, but they cannot bite the guinea pig. In full darkness between 6 pm and 6 am, mosquitoes interested in blood feeding were free to fly through the tunnel, locate the holes and pass through them to reach the guinea pig. The location of mosquitoes after this period was recorded, and those in the section closest to the guinea pig were considered to have interest in blood feeding. Mosquitoes were removed from each section of the tunnel and counted separately.
The choice chambers were maintained at 27.34 °C average temperature (range 27.06 °C to 27.60 °C). The average relative humidity was 76.60% (range 75.90% to 77.00%) The mortality during the assay was recorded, but only live mosquitoes were considered for analysis.
WHO Tube Cloth Spore Dose Estimation
Two protocols were used to determine the number of spores picked up per insect from cloth impregnated with ~2 × 106 conidia/cm2 Met-Hybrid. Black cotton cloth cheaply available in Burkina Faso was used for bioassays. Paper supports were stapled outside of the cloth to maintain rigidity inside the WHO cylinders during the assay. First, 25 female mosquitoes or bees were introduced in the WHO cylinder with impregnated cloths. After 1 h of contact, insects were transferred to plastic cups for 2 h then individually washed in 200 μl of 0.05% Tween-80 by vigorously vortexing them for 5 min. The number of spores picked up by each insect was counted using an improved Neubauer haemocytometer. No fluorescent spores remained on the insects after washing.
In the second protocol used only for mosquitoes, instead of washing the mosquitoes and counting the number of spores, individual mosquitoes were crushed in 200 μl of 0.05% Tween-80, vortexed and the homogenate spread onto Metarhizium selective media26. Five days later, the number of spores per mosquito was determined by counting the colony forming units (CFUs). There were no significant differences in the number of spores per mosquito determined using these two protocols.
The second protocol was also used to count spores delivered by spraying aliquots of 20 female mosquitoes per concentration.
Data analysis
Statistical analyses were carried out with R (version 3.2.4; Supplementary Code). Mosquitoes were considered alive if they could stand upright and dead if they were unresponsive to stimuli following the 2013 recommendations by the WHO Pesticides Evaluation Scheme18.
References
Organization, W. H. & others. World Malaria Report 2015. WHO, Geneva (2015).
Dabiré, K. R. et al. Distribution of pyrethroid and DDT resistance and the L1014F kdr mutation in Anopheles gambiae s.l. from Burkina Faso (West Africa). Trans. R. Soc. Trop. Med. Hyg. 103, 1113–1120, doi:10.1016/j.trstmh.2009.01.008 (2009).
Ranson, H. et al. Insecticide resistance in Anopheles gambiae: data from the first year of a multi-country study highlight the extent of the problem. Malar. J. 8, 299, doi:10.1186/1475-2875-8-299 (2009).
Slater, H. C. et al. Assessing the potential impact of artemisinin and partner drug resistance in sub-Saharan Africa. Malar. J. 15, 10, doi:10.1186/s12936-015-1075-7 (2016).
Bhatt, S. et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 526, 207–211 (2015).
Namountougou, M. et al. Multiple Insecticide Resistance in Anopheles gambiae s.l. Populations from Burkina Faso, West Africa. PLoS One 7, e48412, doi:10.1371/journal.pone.0048412 (2012).
Zlotkin, E., Fishman, Y. & Elazar, M. AaIT: From neurotoxin to insecticide. Biochimie 82, 869–881, doi:10.1016/S0300-9084(00)01177-9 (2000).
Wang, C. & St. Leger, R. J. A scorpion neurotoxin increases the potency of a fungal insecticide. Nat. Biotechnol. 25, 1455–6, doi:10.1038/nbt1357 (2007).
Gordon, D. et al. The differential preference of scorpion α-toxins for insect or mammalian sodium channels: Implications for improved insect control. Toxicon 49, 452–472, doi:10.1016/j.toxicon.2006.11.016 (2007).
Bende, N. S. et al. The insecticidal neurotoxin Aps III is an atypical knottin peptide that potently blocks insect voltage-gated sodium channels. Biochem. Pharmacol. 85, 1542–1554, doi:10.1016/j.bcp.2013.02.030 (2013).
Fletcher, J. I. et al. The structure of a novel insecticidal neurotoxin, ω-atracotoxin-HV1, from the venom of an Australian funnel web spider. Nat. Struct. Biol. 4, 559–566, doi:10.1038/nsb0797-559 (1997).
Undheim, E. A. B. et al. Weaponization of a hormone: convergent recruitment of hyperglycemic hormone into the venom of arthropod predators. Structure 23, 1283–1292, doi:10.1016/j.str.2015.05.003 (2015).
Fang, W., Lu, H.-L., King, G. F. & St. Leger, R. J. Construction of a hypervirulent and specific mycoinsecticide for locust control. Sci. Rep. 4, 7345, doi:10.1038/srep07345 (2014).
Pava-Ripoll, M., Posada, F. J., Momen, B., Wang, C. & St. Leger, R. Increased pathogenicity against coffee berry borer, Hypothenemus hampei (Coleoptera: Curculionidae) by Metarhizium anisopliae expressing the scorpion toxin (AaIT) gene. J. Invertebr. Pathol. 99, 220–226, doi:10.1016/j.jip.2008.05.004 (2008).
Fang, W. et al. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 331, 1074–7, doi:10.1126/science.1199115 (2011).
Scholte, E.-J., Knols, B. G. J. & Takken, W. Infection of the malaria mosquito Anopheles gambiae with the entomopathogenic fungus Metarhizium anisopliae reduces blood feeding and fecundity. J. Invertebr. Pathol. 91, 43–49, doi:10.1016/j.jip.2005.10.006 (2006).
Ogoma, S. B. et al. Screening Mosquito House Entry Points as a Potential Method for Integrated Control of Endophagic Filariasis, Arbovirus and Malaria Vectors. PLoS Negl. Trop. Dis. 4, e773, doi:10.1371/journal.pntd.0000773 (2010).
WHO [World Health Organization]. Guidelines for efficacy testing of spatial repellents. WHO/HTM/NTD/WHOPES/2013 (2013).
Tabashnik, B. E. Evaluation of synergism among Bacillus thuringiensis toxins. Appl. Environ. Microbiol. 58, 3343–6 (1992).
Hu, X. et al. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. Proc. Natl. Acad. Sci. USA 111, 16796–801, doi:10.1073/pnas.1412662111 (2014).
Konrad, B. P. et al. Assessing the optimal virulence of malaria‐targeting mosquito pathogens: a mathematical study of engineered Metarhiziumanisopliae. Malar. J. 13, 11, doi:10.1186/1475-2875-13-11 (2014).
Xu, C. et al. A High-Throughput Gene Disruption Methodology for the Entomopathogenic Fungus Metarhizium robertsii. PLoS One 9, e107657, doi:10.1371/journal.pone.0107657 (2014).
Wang, S., O’Brien, T. R., Pava-Ripoll, M. & St. Leger, R. J. Local adaptation of an introduced transgenic insect fungal pathogen due to new beneficial mutations. Proc. Natl. Acad. Sci. USA 108, 20449–54, doi:10.1073/pnas.1113824108 (2011).
Santolamazza, F. et al. Distribution of knock-down resistance mutations in Anopheles gambiae molecular forms in west and west-central Africa. Malar. J. 7, 74, doi:10.1186/1475-2875-7-74 (2008).
Martinez-Torres, D. et al. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol. Biol. 7, 179–184, doi:10.1046/j.1365-2583.1998.72062.x (1998).
Hu, G. & St. Leger, R. J. Field Studies Using a Recombinant Mycoinsecticide (Metarhizium anisopliae) Reveal that It Is Rhizosphere Competent. Appl. Environ. Microbiol. 68, 6383–6387, doi:10.1128/AEM.68.12.6383-6387.2002 (2002).
Acknowledgements
This work was supported by NIH-NIAID under award RO1-AI106998 to R.J.S. and A.D. and Australian Research Council Discovery Grant DP130103813 to G.F.K.
Author information
Authors and Affiliations
Contributions
R.J.S., A.D., G.F.K., B.L., and E.B. wrote the manuscript and designed experiments. E.B., B.L., W.F., and N.B. performed experiments. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bilgo, E., Lovett, B., Fang, W. et al. Improved efficacy of an arthropod toxin expressing fungus against insecticide-resistant malaria-vector mosquitoes. Sci Rep 7, 3433 (2017). https://doi.org/10.1038/s41598-017-03399-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-03399-0
This article is cited by
-
The need for assessment of risks arising from interactions between NGT organisms from an EU perspective
Environmental Sciences Europe (2023)
-
Genomic Determinants of Entomopathogenic Fungi and Their Involvement in Pathogenesis
Microbial Ecology (2023)
-
Resistance to insecticides in Anopheles gambiae complex in West Africa: A review of the current situation and the perspectives for malaria control
International Journal of Tropical Insect Science (2021)
-
Infection of highly insecticide-resistant malaria vector Anopheles coluzzii with entomopathogenic bacteria Chromobacterium violaceum reduces its survival, blood feeding propensity and fecundity
Malaria Journal (2020)
-
Opinions of key stakeholders on alternative interventions for malaria control and elimination in Tanzania
Malaria Journal (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
