
Abstract
Pregnancy-associated malaria (PAM) is associated with poor pregnancy outcomes. Hemoglobin S (HbS) and hemoglobin C (HbC) mutations are frequently encountered in malaria-endemic areas of Africa, where they protect children from severe and uncomplicated Plasmodium falciparum malaria. However, scant epidemiological data exist on the impact of these Hb variants on PAM. A prospective cohort of 635 Beninese pregnant women was recruited before 24 weeks of gestational age and followed until the end of pregnancy. HbAA, HbAC, and HbAS genotypes were determined and tested for association with pregnancy outcomes and PAM indicators using linear and logistic multivariate models. Newborns from HbAC mothers had higher birthweights than those from HbAA mothers among women infected at any time during pregnancy (mean difference 182.9 g, p = 0.08), or during the first half of pregnancy (654.3 g, p = 0.0006). No such birthweight differences were observed between newborns from HbAS and HbAA mothers. HbAC and HbAS were not associated with other pregnancy outcomes or PAM indicators. In conclusion, HbAC but not HbAS is associated with an improved birth outcome in pregnant women with documented PAM. Higher-birthweight newborns from HbAC mothers may have a survival advantage that contributes to the natural selection of HbC in malaria-endemic areas.
Similar content being viewed by others
Introduction
In endemic areas, the morbidity and mortality associated with Plasmodium falciparum malaria have naturally selected mutations in the β-chain of hemoglobin gene (HBB), giving rise to the HbS and HbC hemoglobinopathies1. While HbS is encountered at frequencies up to 18% across sub-Saharan Africa, the Middle East, and India, HbC is prevalent only in West Africa2. HbS homozygosity results in sickle-cell disease and is associated with high mortality rates while HbC homozygosity is clinically benign2, as are the heterozygous HbAS and HbAC traits. These HBB mutations are known to confer protection from life-threatening P. falciparum malaria3,4,5,6,7,8. For example, HbAS has been associated with 50% and 80% reduced risks of developing uncomplicated and severe malaria6, and HbAC and HbCC have been associated with 29% and 93% reduced risks of developing severe malaria, respectively4. Additional studies have confirmed the malaria-protective effects of HbC5, 9, 10.
Each year, over 50 million pregnant women living in malaria-endemic areas are at risk of developing pregnancy-associated malaria (PAM) caused by P. falciparum 11. PAM is characterized by massive accumulation of parasitized erythrocytes and monocytes in the placental intervillous blood spaces12, leading to serious adverse clinical outcomes for both the mother and the child13, 14. PAM-associated low birthweight is estimated to cause about 62,000 to 363,000 infant deaths every year in Africa alone15. Whether HbAS and HbAC confer protection against this and other detrimental PAM outcomes has not been adequately assessed. One study found no difference in the prevalence of peripheral P. falciparum parasitemia between pregnant HbAA and HbAS women in Gabon16, while another did not associate HbAS with protection from PAM or its adverse effects in pregnant women in Malawi17. Studying the patterns of differential susceptibility to PAM could help in identifying correlates of both malaria pathogenesis and immunity, and then candidate protective molecular mechanisms. Such mechanisms could then be the foundation for the rational design of vaccine candidates as well as preventive measures and treatments against PAM. Our study thus aimed to investigate, for the first time within a single cohort, the effects of maternal HbAS and HbAC genotypes on pregnancy outcomes.
Results
Study Population and HBB genotyping
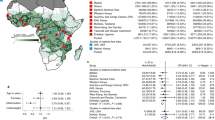
The study was conducted in Comè District, 70 km west of Cotonou, Benin (Fig. 1). The characteristics of 602 pregnant women included in this study are shown in Table 1. Among them, 53 (8.8%) and 95 (15.8%) carried HbAC and HbAS genotypes, respectively. The average ages of HbAA, HbAC, and HbAS women were 27.1, 28.1, and 26.6 years, and did not differ by HBB genotype. Peda was the more prevalent ethnicity in HbAA and HbAS women, while Watchi was the most prevalent in HbAC women. The mean gestational age determined by ultrasound was 39.7 weeks for all three groups of women, which also did not differ in BMI, ABO blood group, or number of P. falciparum infections.

Map of the study area. The study was conducted in Comè District, 70 km west of Cotonou, Benin, where perennial malaria transmission peaks from April to July and September to November. Three dispensaries (Comè, Akodeha, and Ouedeme Pedah), 10 km away from each other, were involved. Comè is a semi-rural area and the two other sites are located in more rural settings. Maps credit: http://www.carte-monde.org/cartes-du-benin/ and P. Deloron.
Assessment of maternal HBB genotype on newborn birthweight
We first assessed the effects of maternal HBB genotype on newborn birthweight. Analyses excluded premature births (n = 35) and stillbirths (n = 16). In the whole cohort, mean (SD) birthweights for HbAA, HbAC, and HbAS mothers were 3049.4 g (412), 3165.2 g (453), and 3106.1 g (408), respectively (Fig. 2a). In a multivariate regression model adjusted on low maternal BMI, gestational age, parity status, fetal sex, malaria infection at inclusion, ethnicity, and season at the beginning of pregnancy, no effects of HBB genotype in mothers were observed. Birthweights were significantly lower among primigravid mothers (mean difference −107.9 g; 95% confidence interval [95% CI] −198.8, −17.02; p = 0.02), women with parasite infection at inclusion (mean difference −137.6 g; 95% CI −258.4, −16.8; p = 0.02), and women exposed to the peak malaria season during the first trimester of pregnancy (mean difference −89.03 g; 95% CI −158.4, −19.6; p = 0.02). Mean birthweight appeared to be lower for women from Adja and Peda ethnic groups compared to the others, but no significant differences were observed between ethnic groups (global p = 0.36).

Stacked histograms of newborn birthweights according to maternal HBB genotypes in the whole cohort and in women with malaria infection during pregnancy. (a) Entire cohort (n = 551; 417 HbAA, 46 HbAC, 88 HbAS). (b) Mothers infected at least once during pregnancy (n = 218; 168 HbAA, 17 AC, 33 AS). (c) Mothers infected before 20 weeks of pregnancy (n = 80; 61 HbAA, 7 HbAC, 12 HbAS). Premature births and stillbirths were excluded.
Analyses were further conducted on the sub-samples of women infected at least once during pregnancy (n = 218) and women with an early infection during pregnancy (n = 80), since we previously reported in the same cohort that only early infections during pregnancy were associated with a mean birthweight decrease of 98.5 g (95% CI −188.5, −8.5)18. Among women with at least one P. falciparum infection during pregnancy, 168, 17, and 33 of them carried the HbAA, HbAC, and HbAS genotypes, respectively. Mean (SD) birthweights in these three groups were 3013.4 g (426), 3197.7 g (507), and 3044.2 g (368) (Fig. 2b). After adjustment, although birthweights were similar among infants born to HbAA and HbAS mothers, infants born to HbAC mothers showed a trend towards higher birthweights, although this did not reach statistical significance (mean difference 182.9 g; 95% CI −18.9, 384.8; p = 0.08) (Table 2).
Exposure to the peak malaria season during the first trimester of pregnancy (p = 0.0096) and parasite infection at inclusion (p = 0.05) were also associated with a lower mean birthweight.
For women infected before 20 weeks of gestation, newborn mean (SD) birthweights were 2934.1 g (416), 2918.3 g (359) and 3611.4 g (451) for mothers carrying HbAA (n = 61), HbAS (n = 12), and HbAC (n = 7) genotypes, respectively (Fig. 2c). In a multivariate regression model, HbAC mothers had significantly higher newborn birthweights than HbAA mothers (mean difference 654.3 g; 95% CI 298.3, 1019.1; p = 0.0006) (Table 2). After removing one outlier (a 4500 g newborn from an HbAC mother), this birthweight difference remained significant (418.7 g; 54.5, 783.0; p = 0.025). As in the group of infected women, newborn birthweights among HbAS and HbAA mothers did not differ (Table 2).
Since we previously reported in this same cohort that sub-microscopic P. falciparum infections (those detected only by PCR, not by thick blood smear examination) are associated with decreased maternal Hb levels and low newborn birthweight19, we tested the interactions between maternal HBB genotype and presence of sub-microscopic parasitemia on birthweight. Newborn birthweights among 44 mothers with sub-microscopic parasitemia early in pregnancy did not differ from those of uninfected mothers (interaction p = 0.79).
When analysing low birthweight (<2500 g) using a multivariate logistic regression model, we detected no effect of HBB genotype or any covariates (data not shown). However, this analysis is underpowered since low birthweights were observed in only 5.4% of newborns (after excluding premature births), and included only one from an HbAC mother and five from HbAS mothers.
Assessment of maternal HBB genotype on prematurity, number of infections during pregnancy, and parasite density
We further analysed prematurity, number of infections during pregnancy, and parasite density according to HBB genotype. Of the 35 premature births, five and six occurred in HbAC and HbAS mothers, respectively. Premature births were associated with low BMI (p = 0.004) and first pregnancy (p = 0.05), but not HBB genotype (p = 0.24 for HbAC, p = 0.64 for HbAS) or P. falciparum infection.
We next investigated the relationship between HBB genotype and number of infections during pregnancy and parasite density (Fig. 3). HbAC women had fewer infections than HbAA and HbAS women (Fig. 3a), but this difference was not significant (Kruskal-Wallis test, p = 0.10). We also performed a multivariate analysis on the entire cohort to test for effects on the number of infections during pregnancy, after excluding women with <10 weeks of follow-up (n = 598). After adjustment on length of follow-up and season of pregnancy, increased number of infections during pregnancy was associated with low BMI (p = 0.038) and first pregnancy (p = 0.011), but not maternal HbAC (p = 0.32) or HbAS (p = 0.64) genotypes. In the subset of infected women, mean parasite densities were similar among HbAA, HbAS, and HbAC women (Kruskal-Wallis test, p = 0.73) (Fig. 3b). Therefore, the higher birthweights among HbAC mothers do not seem attributable to differences in the number or density of parasite infections.

Number (a) and density (b) of Plasmodium falciparum infections among women infected at least once (n = 234), according to HBB genotype. (a) Stacked bar chart of the number of P. falciparum infections by genotype group. The number of infections per woman is divided in four categories: 1, 2, 3, or 4 and more infections during the pregnancy (grey scale). Each bar represents the percentage of each infection categories according to HBB genotype group. (b) Parasite density measured by microscopy according to genotype group. Values are log-transformed parasite densities (parasites/µl) for women with one infection or the geometric mean of parasite densities for women with more than one infection. Median, interquartile range, range, and outliers are represented.
Discussion
Although it is well documented that HbAS confers malaria protection to African children3, 5, 6, HbAS had no discernible impact on newborn birthweight or other pregnancy outcomes in our study of Beninese women. These results are consistent with a recent study of pregnant Malawian women, which found no association between HbAS and PAM indicators17. The increased newborn birthweights we observed among HbAC but not HbAS women infected with P. falciparum during pregnancy suggest that these two HBB genotypes differentially impact PAM. Indeed, mean birthweight among HbAA and HbAS mothers, but not HbAC mothers, decreases successively in the whole cohort, the subset of all infected women, and the subset of early-infected women in our study. Early infections during pregnancy are known to impact fetal development and thus associated with the occurrence of low birthweight. Strikingly, HbAC women infected before 20 weeks of gestation delivered newborns with a much higher birthweight than did early-infected HbAA and HbAS women, and even higher than newborns delivered by uninfected women. Given our study design and the follow-up strategy, it is still possible that we missed some infections, especially those during the first trimester before the inclusion of women. This could explain the difference observed even among women classified here as uninfected, and further suggests that the protective effect of HbAC reported in this study is significant and worthy of additional investigation.
Although few HbAC primigravidae were sampled in our study, and none were infected early in pregnancy, these limitations do not account for higher birthweights among HbAC mothers because this effect was adjusted on parity in multivariate analysis. Although we observed no differences in the number or density of peripheral blood parasitemias, these data may not adequately reflect the placental parasite load and thus the severity of PAM.
It was previously shown that parasitized HbAS and HbAC erythrocytes have reduced capacities to cytoadhere to host receptors on microvascular endothelial cells, due to significantly decreased and aberrant exposure of the parasite’s cytoadherence ligand PfEMP1 on their surface20, 21. Since the PfEMP1 variant VAR2CSA mediates cytoadhesion to chondroitin sulfate A (CSA) on placental syncytiotrophoblast cells22,23,24,25, we expected to observe increased newborn birthweights among both HbAS and HbAC women resulting from similarly reduced parasite sequestration in the placenta.
It is therefore possible that HbAC increases birthweights among early-infected women by a different mechanism, or that HbAC more strongly impairs parasite cytoadherence in the placenta, where blood flow in the intervillous space is slower than in the microvasculature, and where CSA (instead of CD36) is the predominant host cytoadherence receptor. In support of this latter hypothesis, two recently-published studies have shown that in static adhesion assays, VAR2CSA-expressing FCR3 parasites in HbAS and HbAC erythrocytes exhibit significant reductions in adherence to CSA compared to those grown in HbAA erythrocytes, this decreased binding being much greater for parasitized HbAC than HbAS erythrocytes26, 27. However, these studies assessed only a single parasite strain in static but not flow adhesion assays. This observation supports and could explain our finding of a protective effect of HbAC, but not HbAS, against PAM.
This study was primarily designed to characterize the effects of malaria infections in pregnant women, rather than the effects of hemoglobin variants, and thus has limitations. The main limitation is obviously owing to the size of the study population. Although a significant number of women were included, only limited proportions of them carried HbS (15.8%) or the HbC (8.8%) genotypes, or were infected (36.2%) during pregnancy. Therefore, selected analyses were conducted on a limited number of women, and were relatively underpowered. Nevertheless, this does not diminish the value of our main significant finding that infected HbAC mothers had significantly higher newborn birthweights than infected HbAA mothers. Secondly, for obvious ethical reasons, all women received two doses of sulfadoxine-pyrimethamine as intermittent preventive treatment during pregnancy, and were also treated if an infection was detected at unscheduled visits or when they presented to clinic for health-related reasons. Therefore, our results are likely to be hampered by the effects of these drug treatments.
In conclusion, we report for the first time that HbAC is associated with an improved birth outcome in infected pregnant women, which may perhaps contribute to the natural selection of HbC in the human populations of malaria-endemic West Africa. Further studies are needed to confirm that HbAC is associated with an improve birth outcome in women with PAM, and to discover the precise mechanism by which HbAC increases newborn birthweight in order to recapitulate it as a new therapy or vaccine against PAM.
Methods
Ethics statement
The study received ethical approval from the Comité Consultatif de Déontologie et d’Éthique of IRD in France and the Comité d’Éthique de la Faculté des Science de la Santé, Université d’Abomey Calavi in Benin (FSS 026/2007/CE/FSS/UAC). All study procedures were performed in accordance with the institutional policies, guidelines and regulations pertaining to research involving human subjects. All adult participants, or the adult caretakers of minor participants, provided written informed consent.
Study area and population
The study was conducted between 2008 and 2011 in Comè District (Fig. 1), a semi-rural area 70 km west of Cotonou, Benin, where perennial malaria transmission peaks from April to July and September to November. A detailed description of the study area has been given elsewhere18, 28, 29. At the time of follow-up, the entomological inoculation rate was 35–60 infective bites per person per year30 and P. falciparum was the predominant species.
A prospective cohort of pregnant women was recruited before 24 weeks of gestation. Women were followed monthly from inclusion to delivery, and received two doses of sulfadoxine-pyrimethamine as intermittent preventive treatment during pregnancy (IPTp) per national guidelines. At inclusion, at each antenatal visit, and at unscheduled visits, when women presented to the clinic for health reasons, a rapid diagnostic test (Parascreen™, Zephyr Biomedicals, Goa, India) for P. falciparum infection was performed on capillary blood, venous blood was drawn, and Hb concentration was measured by the Hemocue® Hb 201+ system (HemoCue AB, Angelholm, Sweden).
Pregnancy was followed by four transabdominal ultrasound scans with a portable system (Titan, Sonosite, Bothell, WA) by specifically trained midwives. The first scan estimated the gestational age, and the others assessed intrauterine growth and fetal morphology. Placental and peripheral blood samples were collected at delivery. Thick blood smears were prepared from all peripheral blood samples, and Giemsa-stained P. falciparum parasites were counted against 200 leukocytes by two independent experienced microscopists, achieving a detection threshold of 40 parasites/µl. Four 50 µl drops of blood were spotted onto Whatman 3 paper, dried at room temperature, and stored until DNA was extracted by the Chelex method31. A real-time PCR assay to detect sub-microscopic P. falciparum infection was performed on samples collected at the times of inclusion, second IPTp uptake, one month before delivery, and at delivery19.
HBB genotyping
HBB genotyping was performed on the first exon of the β-globin gene amplified (678 bp) by polymerase chain reaction (PCR) with Jump Taq (Sigma). The following oligonucleotides were used in the PCR reaction: oligonucleotide sense 5′-AGGAGCAGGGAGGGCAGGAG-3′ and antisense 5′-GCAATCATTCGTCTGTTTCCCA-3′. PCR products were sent to GATC Biotech (Mulhouse, France) for Sanger sequencing.
Statistical analysis
Linear and logistic multivariate models were used to test the effects of HbAC and HbAS on pregnancy outcomes (newborn birthweight, low birthweight (<2500 g), premature birth, maternal anemia at delivery) and PAM indicators (number and density of P. falciparum infections during pregnancy). Data from 33 women were excluded from analysis due to the presence of HbCC (n = 2) or HbSC (n = 3), lack of prenatal consultation (n = 10), or twin pregnancy (n = 18). Covariates for adjustment in multivariate models were selected on the basis of previous works published by our team on birthweight and maternal anemia in the same cohort18, 19. We considered gestational age at delivery (estimated by an ultrasound scan at inclusion, and transformed into a four-class variable using quartiles); low body mass index (BMI) at inclusion (<18.5, ≥18.5); gravidity (primigravid, multigravid); fetal sex; and the number of antenatal visits; P. falciparum infection status during pregnancy (0, ≥1); total number of P. falciparum infections during pregnancy (0, 1–2, ≥3); P. falciparum infection at inclusion (yes, no); and exposure to the peak malaria season during the first trimester of pregnancy (beginning of pregnancy between April and September). Then a stepwise procedure with a backward elimination was used to choose the covariates to retain in the final models (p < 0.20). For birthweight outcome, as malaria infection may act as a modifier of the relationship between HBB genotype and birthweight, analyses were performed on all women; a subset of women infected at least once during pregnancy; and a subset of women infected in the first 4 months of pregnancy.
During the first 4 months of pregnancy, the effects of sub-microscopic parasitemia (detected by PCR, not thick blood smear examination) were also assessed. To distinguish the proper effect of sub-microscopic parasitemia, a three-class variable was defined: “microscopically infected” (at least one infection detected by thick blood smear), “sub-microscopically infected” (one infection detected by PCR but not by thick blood smear), and “not infected” (otherwise). This variable was included in the linear multivariate model of birthweight on the whole cohort. The interaction between infection status during the first 4 months of pregnancy and HBB genotype was also tested.
The number of infections detected by microscopy during pregnancy was compared using a negative binomial regression on the entire cohort and on a subset of women infected at least once. Analyses were performed after excluding women with <10 weeks of follow-up and were adjusted on the length of follow-up (in weeks), BMI, ABO blood group, gravidity, and season of pregnancy. Among women with at least one infection, the mean parasite densities by subject were compared within HBB genotype groups using Kruskal-Wallis test. For each subject, we computed the mean of log-transformed parasite densities observed by microscopy.
All regression models were adjusted on ethnic group. Analyses were performed using R software v3.2.3 (http://www.R-project.org/)32.
References
Weatherall, D. J. & Provan, A. B. Red cells I: inherited anaemias. Lancet 355, 1169–1175, doi:S0140673600020730 (2000).
Piel, F. B. et al. The distribution of haemoglobin C and its prevalence in newborns in Africa. Sci Rep 3, 1671, doi:10.1038/srep01671 (2013).
Lopera-Mesa, T. M. et al. Effect of red blood cell variants on childhood malaria in Mali: a prospective cohort study. Lancet Haematol 2, e140–149, doi:10.1016/S2352-3026(15)00043-5 (2015).
Modiano, D. et al. Haemoglobin C protects against clinical Plasmodium falciparum malaria. Nature 414, 305–308, doi:10.1038/35104556 (2001).
Taylor, S. M., Parobek, C. M. & Fairhurst, R. M. Haemoglobinopathies and the clinical epidemiology of malaria: a systematic review and meta-analysis. Lancet Infect Dis 12, 457–468, doi:10.1016/S1473-3099(12)70055-5 (2012).
Williams, T. N. et al. Sickle cell trait and the risk of Plasmodium falciparum malaria and other childhood diseases. J Infect Dis 192, 178–186, doi:10.1086/430744 (2005).
Williams, T. N. et al. Negative epistasis between the malaria-protective effects of alpha+ -thalassemia and the sickle cell trait. Nat Genet 37, 1253–1257, doi:ng1660 10.1038/ng1660 (2005).
Williams, T. N. Red blood cell variants and malaria: a long story not yet over. Lancet Haematol 2, e130–131, doi:10.1016/S2352-3026(15)00049-6 (2015).
Agarwal, A. et al. Hemoglobin C associated with protection from severe malaria in the Dogon of Mali, a West African population with a low prevalence of hemoglobin S. Blood 96, 2358–2363 (2000).
Travassos, M. A. et al. Hemoglobin C Trait Provides Protection From Clinical Falciparum Malaria in Malian Children. J Infect Dis 212, 1778–1786, doi:10.1093/infdis/jiv308 (2015).
WHO. World Malaria Report 2015. (2015).
Muehlenbachs, A., Mutabingwa, T. K., Edmonds, S., Fried, M. & Duffy, P. E. Hypertension and maternal-fetal conflict during placental malaria. PLoS Med 3, e446, doi:10.1371/journal.pmed.0030446 (2006).
Umbers, A. J., Aitken, E. H. & Rogerson, S. J. Malaria in pregnancy: small babies, big problem. Trends Parasitol 27, 168–175, doi:10.1016/j.pt.2011.01.007 (2011).
Desai, M. et al. Epidemiology and burden of malaria in pregnancy. Lancet Infect Dis 7, 93–104, doi:10.1016/S1473-3099(07)70021-X (2007).
Steketee, R. W., Nahlen, B. L., Parise, M. E. & Menendez, C. The burden of malaria in pregnancy in malaria-endemic areas. Am J Trop Med Hyg 64, 28–35, doi:10.4269/ajtmh.2001.64.28 (2001).
Bouyou-Akotet, M. K. et al. Prevalence of Plasmodium falciparum infection in pregnant women in Gabon. Malar J 2, 18, doi:10.1186/1475-2875-2-18 (2003).
Patel, J. C. et al. Absence of Association Between Sickle Trait Hemoglobin and Placental Malaria Outcomes. Am J Trop Med Hyg 94, 1002–1007, doi:10.4269/ajtmh.15-0672 (2016).
Huynh, B. T. et al. Influence of the timing of malaria infection during pregnancy on birth weight and on maternal anemia in Benin. Am J Trop Med Hyg 85, 214–220, doi:10.4269/ajtmh.2011.11-0103 (2011).
Cottrell, G. et al. Submicroscopic Plasmodium falciparum Infections Are Associated With Maternal Anemia, Premature Births, and Low Birth Weight. Clin Infect Dis 60, 1481–1488, doi:10.1093/cid/civ122 (2015).
Fairhurst, R. M. et al. Abnormal display of PfEMP-1 on erythrocytes carrying haemoglobin C may protect against malaria. Nature 435, 1117–1121, doi:10.1038/nature03631 (2005).
Fairhurst, R. M., Bess, C. D. & Krause, M. A. Abnormal PfEMP1/knob display on Plasmodium falciparum-infected erythrocytes containing hemoglobin variants: fresh insights into malaria pathogenesis and protection. Microbes Infect 14, 851–862, doi:10.1016/j.micinf.2012.05.006 (2012).
Srivastava, A. et al. Full-length extracellular region of the var2CSA variant of PfEMP1 is required for specific, high-affinity binding to CSA. Proc Natl Acad Sci USA 107, 4884–4889, doi:10.1073/pnas.1000951107 (2010).
Viebig, N. K. et al. A single member of the Plasmodium falciparum var multigene family determines cytoadhesion to the placental receptor chondroitin sulphate A. EMBO Rep 6, 775–781, doi:7400466 10.1038/sj.embor.7400466 (2005).
Gamain, B. et al. Identification of multiple chondroitin sulfate A (CSA)-binding domains in the var2CSA gene transcribed in CSA-binding parasites. J Infect Dis 191, 1010–1013, doi:JID33609 10.1086/428137 (2005).
Viebig, N. K. et al. Disruption of var2csa gene impairs placental malaria associated adhesion phenotype. PLoS One 2, e910, doi:10.1371/journal.pone.0000910 (2007).
Cyrklaff, M. et al. Oxidative insult can induce malaria-protective trait of sickle and fetal erythrocytes. Nat Commun 7, 13401, doi:10.1038/ncomms13401 (2016).
Kilian, N. et al. Hemoglobin S and C affect protein export in Plasmodium falciparum-infected erythrocytes. Biol Open 4, 400–410, doi:10.1242/bio.201410942 (2015).
Ndam, N. T. et al. Protective Antibodies against Placental Malaria and Poor Outcomes during Pregnancy, Benin. Emerg Infect Dis 21, 813–823, doi:10.3201/eid2105.141626 (2015).
Huynh, B. T. et al. Consequences of gestational malaria on birth weight: finding the best timeframe for intermittent preventive treatment administration. PLoS One 7, e35342, doi:10.1371/journal.pone.0035342 (2012).
Kelly-Hope, L. A. & McKenzie, F. E. The multiplicity of malaria transmission: a review of entomological inoculation rate measurements and methods across sub-Saharan Africa. Malar J 8, 19, doi:10.1186/1475-2875-8-19 (2009).
Wooden, J., Kyes, S. & Sibley, C. H. PCR and strain identification in Plasmodium falciparum. Parasitol Today 9, 303–305, doi:10.1016/0169-4758(93)90131-X (1993).
Core-Team, R. R A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria (2013).
Acknowledgements
This study was part of the STOPPAM collaborative project “Strategies To Prevent Pregnancy-Associated Malaria,” and P.D. and N.T.N. were supported by the European 7th Framework Programme (contract no. 200889). R.M.F. is supported by the Intramural Research Program, NIAID, NIH. B.G. is supported by the Institut National de la Recherche Médicale. We are grateful to the women who participated in this study.
Author information
Authors and Affiliations
Contributions
M.T., J.M., and S.D. performed the experiments. J.E., and N.F. designed the experiments and analysed the data; M.T., J.M., S.D., D.D.S., R.M.F., P.D., N.T.N., and B.G. designed the experiments, analysed the data, and wrote the paper.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tétard, M., Milet, J., Dechavanne, S. et al. Heterozygous HbAC but not HbAS is associated with higher newborn birthweight among women with pregnancy-associated malaria. Sci Rep 7, 1414 (2017). https://doi.org/10.1038/s41598-017-01495-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-01495-9
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
