
Abstract
The stability and shape of the erythrocyte membrane is provided by the ankyrin-1 complex, but how it tethers the spectrin–actin cytoskeleton to the lipid bilayer and the nature of its association with the band 3 anion exchanger and the Rhesus glycoproteins remains unknown. Here we present structures of ankyrin-1 complexes purified from human erythrocytes. We reveal the architecture of a core complex of ankyrin-1, the Rhesus proteins RhAG and RhCE, the band 3 anion exchanger, protein 4.2, glycophorin A and glycophorin B. The distinct T-shaped conformation of membrane-bound ankyrin-1 facilitates recognition of RhCE and, unexpectedly, the water channel aquaporin-1. Together, our results uncover the molecular details of ankyrin-1 association with the erythrocyte membrane, and illustrate the mechanism of ankyrin-mediated membrane protein clustering.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


Data availability
All raw video files, aligned micrographs and particle coordinates (in two STAR files, one for band 3–GPA and one for the ankyrin complexes) are deposited in the EMPIAR (EMPIAR-11043). The cryo-EM density maps and models have been deposited in the Electron Microscopy Data Bank and the PDB. PDB IDs are: 7UZ3, 7UZE, 7UZQ, 7UZS, 7UZU, 7UZV, 7V07, 7V0K, 7V0M, 7V0Q, 7V0S, 7V0T, 7V0U, 7V0X, 7V0Y, 7V19, 8CRQ, 8CRR, 8CRT, 8CS9, 8CSV, 8CSW, 8CSX, 8CSY, 8CT2, 8CT3, 8CSL and 8CTE. EMDB IDs are: EMD-26874, EMD-26886, EMD-26916, EMD-26917, EMD-26918, EMD-26919, EMD-26940, EMD-26943, EMD-26944, EMD-26948, EMD-26949, EMD-26950, EMD-26951, EMD-26952, EMD-26953, EMD-26954, EMD-26955, EMD-26956, EMD-26958, EMD-26960, EMD-26972, EMD-26973, EMD-26974, EMD-26975, EMD-26978, EMD-26979, EMD-26982, EMD-26965 and EMD-26988.
References
Marchesi, V. T. & Steers, E. Selective solubilization of a protein component of the red cell membrane. Science 159, 203–204 (1968).
Unsain, N., Stefani, F. D. & Cáceres, A. The actin/spectrin membrane-associated periodic skeleton in neurons. Front. Synaptic Neurosci. https://doi.org/10.3389/fnsyn.2018.00010 (2018).
Bennett, V. & Stenbuck, P. J. Identification and partial purification of ankyrin, the high affinity membrane attachment site for human erythrocyte spectrin. J. Biol. Chem. 254, 2533–2541 (1979).
Bennett, V. & Stenbuck, P. J. The membrane attachment protein for spectrin is associated with band 3 in human erythrocyte membranes. Nature 280, 468–473 (1979).
Stevens, S. R. et al. Ankyrin-R regulates fast-spiking interneuron excitability through perineuronal nets and Kv3.1b K+ channels. eLife 10, e66491 (2021).
Mohler, P. J., Davis, J. Q. & Bennett, V. Ankyrin-B coordinates the Na/K ATPase, Na/Ca exchanger, and InsP3 receptor in a cardiac T-tubule/SR microdomain. PLoS Biol. 3, e423 (2005).
Jenkins, P. M. et al. Giant ankyrin-G: a critical innovation in vertebrate evolution of fast and integrated neuronal signaling. Proc. Natl Acad. Sci. USA 112, 957–964 (2015).
Jenkins, S. M. & Bennett, V. Ankyrin-G coordinates assembly of the spectrin-based membrane skeleton, voltage-gated sodium channels, and L1 CAMs at Purkinje neuron initial segments. J. Cell Biol. 155, 739–746 (2001).
Bennett, V. Purification of an active proteolytic fragment of the membrane attachment site for human erythrocyte spectrin. J. Biol. Chem. 253, 2292–2299 (1978).
Perrotta, S., Gallagher, P. G. & Mohandas, N. Hereditary spherocytosis. Lancet 372, 1411–1426 (2008).
Bennett, V. & Healy, J. Organizing the fluid membrane bilayer: diseases linked to spectrin and ankyrin. Trends Mol. Med. 14, 28–36 (2008).
Nicolas, V. et al. Rh-RhAG/ankyrin-R, a new interaction site between the membrane bilayer and the red cell skeleton, is impaired by Rh(null)-associated mutation. J. Biol. Chem. 278, 25526–25533 (2003).
Jiang, W. et al. Interaction of glucose transporter 1 with anion exchanger 1 in vitro. Biochem. Biophys. Res. Commun. 339, 1255–1261 (2006).
Mankelow, T. J., Satchwell, T. J. & Burton, N. M. Refined views of multi-protein complexes in the erythrocyte membrane. Blood Cells Mol. Dis. 49, 1–10 (2012).
Young, G. et al. Quantitative mass imaging of single biological macromolecules. Science 360, 423–427 (2018).
Bepler, T. et al. Positive-unlabeled convolutional neural networks for particle picking in cryo-electron micrographs. Nat. Methods 16, 1153–1160 (2019).
Chen, K., Li, J., Wang, C., Wei, Z. & Zhang, M. Autoinhibition of ankyrin-B/G membrane target bindings by intrinsically disordered segments from the tail regions. eLife 6, e29150 (2017).
Wang, C. et al. Structural basis of diverse membrane target recognitions by ankyrins. eLife https://doi.org/10.7554/eLife.04353 (2014).
Toye, A. M. et al. Protein-4.2 association with band 3 (AE1, SLCA4) in Xenopus oocytes: effects of three natural protein-4.2 mutations associated with hemolytic anemia. Blood 105, 4088–4095 (2005).
Satchwell, T. J., Shoemark, D. K., Sessions, R. B. & Toye, A. M. Protein 4.2: a complex linker. Blood Cells Mol. Dis. 42, 201–210 (2009).
Risinger, M. A., Dotimas, E. M. & Cohen, C. M. Human erythrocyte protein 4.2, a high copy number membrane protein, is N-myristylated. J. Biol. Chem. 267, 5680–5685 (1992).
Beckmann, R., Smythe, J. S., Anstee, D. J. & Tanner, M. J. Coexpression of band 3 mutants and Rh polypeptides: differential effects of band 3 on the expression of the Rh complex containing D polypeptide and the Rh complex containing CcEe polypeptide. Blood 97, 2496–2505 (2001).
Hanicak, A. et al. Erythrocyte band 3 protein strongly interacts with phosphoinositides. FEBS Lett. 348, 169–172 (1994).
Bruce, L. J. et al. A band 3-based macrocomplex of integral and peripheral proteins in the RBC membrane. Blood 101, 4180–4188 (2003).
Perrotta, S. et al. The N-terminal 11 amino acids of human erythrocyte band 3 are critical for aldolase binding and protein phosphorylation: implications for band 3 function. Blood 106, 4359–4366 (2005).
Ferru, E. et al. Regulation of membrane-cytoskeletal interactions by tyrosine phosphorylation of erythrocyte band 3. Blood 117, 5998–6006 (2011).
Ding, Y., Casey, J. R. & Kopito, R. R. The major kidney AE1 isoform does not bind ankyrin (Ank1) in vitro. An essential role for the 79 NH2-terminal amino acid residues of band 3. J. Biol. Chem. 269, 32201–32208 (1994).
Kim, S. et al. Determination of structural models of the complex between the cytoplasmic domain of erythrocyte band 3 and ankyrin-R repeats 13-24. J. Biol. Chem. 286, 20746–20757 (2011).
Chang, S. H. & Low, P. S. Identification of a critical ankyrin-binding loop on the cytoplasmic domain of erythrocyte membrane band 3 by crystal structure analysis and site-directed mutagenesis. J. Biol. Chem. 278, 6879–6884 (2003).
Bruce, L. J. et al. Changes in the blood group Wright antigens are associated with a mutation at amino acid 658 in human erythrocyte band 3: a site of interaction between band 3 and glycophorin A under certain conditions. Blood 85, 541–547 (1995).
Williamson, R. C. & Toye, A. M. Glycophorin A: band 3 aid. Blood Cells Mol. Dis. 41, 35–43 (2008).
Young, M. T., Beckmann, R., Toye, A. M. & Tanner, M. J. Red-cell glycophorin A-band 3 interactions associated with the movement of band 3 to the cell surface. Biochem. J. 350 Pt 1, 53–60 (2000).
Trenker, R., Call, M. E. & Call, M. J. Crystal structure of the glycophorin A transmembrane dimer in lipidic cubic phase. J. Am. Chem. Soc. 137, 15676–15679 (2015).
Kalli, A. C. & Reithmeier, R. A. F. Interaction of the human erythrocyte Band 3 anion exchanger 1 (AE1, SLC4A1) with lipids and glycophorin A: molecular organization of the Wright (Wr) blood group antigen. PLoS Comput. Biol. 14, e1006284 (2018).
Taylor, A. M., Boulter, J., Harding, S. E., Cölfen, H. & Watts, A. Hydrodynamic properties of human erythrocyte band 3 solubilized in reduced Triton X-100. Biophys. J. 76, 2043–2055 (1999).
Su, Y. et al. Associations of protein 4.2 with band 3 and ankyrin. Mol. Cell Biochem. 289, 159–166 (2006).
Hsu, K. et al. Adaptable interaction between aquaporin-1 and band 3 reveals a potential role of water channel in blood CO2 transport. FASEB J. 31, 4256–4264 (2017).
Reithmeier, R. A. A membrane metabolon linking carbonic anhydrase with chloride/bicarbonate anion exchangers. Blood Cells Mol. Dis. 27, 85–89 (2001).
Vince, J. W. & Reithmeier, R. A. Carbonic anhydrase II binds to the carboxyl terminus of human band 3, the erythrocyte C1-/HCO3- exchanger. J. Biol. Chem. 273, 28430–28437 (1998).
Endeward, V. et al. Evidence that aquaporin 1 is a major pathway for CO2 transport across the human erythrocyte membrane. FASEB J. 20, 1974–1981 (2006).
Planelles, G. Ammonium homeostasis and human Rhesus glycoproteins. Nephron Physiol. 105, p11–p17 (2007).
Gruswitz, F. et al. Function of human Rh based on structure of RhCG at 2.1 A. Proc. Natl Acad. Sci. USA 107, 9638–9643 (2010).
Geyer, R. R., Parker, M. D., Toye, A. M., Boron, W. F. & Musa-Aziz, R. Relative CO2/NH3 permeabilities of human RhAG, RhBG and RhCG. J. Membr. Biol. 246, 915–926 (2013).
Endeward, V., Cartron, J. & Ripoche, P., Gros, and G. RhAG protein of the Rhesus complex is a CO2 channel in the human red cell membrane. FASEB J. 22, 64–73 (2008).
Burton, N. M. & Anstee, D. J. Structure, function and significance of Rh proteins in red cells. Curr. Opin. Hematol. 15, 625–630 (2008).
Conroy, M. J., Bullough, P. A., Merrick, M. & Avent, N. D. Modelling the human rhesus proteins: implications for structure and function. Br. J. Haematol. 131, 543–551 (2005).
Niggli, V. & Carafoli, E. The plasma membrane Ca(2+) ATPase: purification by calmodulin affinity chromatography, and reconstitution of the purified protein. Methods Mol. Biol. 1377, 57–70 (2016).
Suloway, C. et al. Fully automated, sequential tilt-series acquisition with Leginon. J. Struct. Biol. 167, 11–18 (2009).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Pettersen, E. F. et al. UCSF Chimera–a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife 7, e42166 (2018).
Yang, J. et al. Improved protein structure prediction using predicted interresidue orientations. Proc. Natl Acad. Sci. USA 117, 1496–1503 (2020).
Michaely, P., Tomchick, D. R., Machius, M. & Anderson, R. G. W. Crystal structure of a 12 ANK repeat stack from human ankyrinR. EMBO J. 21, 6387–6396 (2002).
Arakawa, T. et al. Crystal structure of the anion exchanger domain of human erythrocyte band 3. Science 350, 680–684 (2015).
Zhang, D., Kiyatkin, A., Bolin, J. T. & Low, P. S. Crystallographic structure and functional interpretation of the cytoplasmic domain of erythrocyte membrane band 3. Blood 96, 2925–2933 (2000).
Ruiz Carrillo, D. et al. Crystallization and preliminary crystallographic analysis of human aquaporin 1 at a resolution of 3.28Å. Acta Crystallogr. F. Struct. Biol. Commun. 70, 1657–1663 (2014).
Stein, N. CHAINSAW: a program for mutating PDB files used as templates in molecular replacement. J. Appl. Crystallogr. 41, 641–643 (2008).
Winn, M. D. et al. Overview of the CCP 4 suite and current developments. Acta Crystallogr. D. Biol. Crystallogr. 67, 235–242 (2011).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D. Biol. Crystallogr. 66, 486–501 (2010).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D. Biol. Crystallogr. 60, 2126–2132 (2004).
Casañal, A., Lohkamp, B. & Emsley, P. Current developments in Coot for macromolecular model building of electron cryo-microscopy and crystallographic data. Protein Sci. 29, 1069–1078 (2020).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D. Struct. Biol. 74, 531–544 (2018).
Pettersen, E. F. et al. UCSF ChimeraX: structure visualization for researchers, educators, and developers. Protein Sci. 30, 70–82 (2021).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Noble, A. J. & Stagg, S. M. Automated batch fiducial-less tilt-series alignment in Appion using Protomo. J. Struct. Biol. 192, 270–278 (2015).
Winkler, H. & Taylor, K. A. Accurate marker-free alignment with simultaneous geometry determination and reconstruction of tilt series in electron tomography. Ultramicroscopy 106, 240–254 (2006).
Lander, G. C. et al. Appion: an integrated, database-driven pipeline to facilitate EM image processing. J. Struct. Biol. 166, 95–102 (2009).
Grant, T. & Grigorieff, N. Measuring the optimal exposure for single particle cryo-EM using a 2.6Å reconstruction of rotavirus VP6. eLife 4, e06980 (2015).
Agulleiro, J. I. & Fernandez, J. J. Fast tomographic reconstruction on multicore computers. Bioinformatics 27, 582–583 (2011).
Agulleiro, J.-I. & Fernandez, J.-J. Tomo3D 2.0 – exploitation of advanced vector extensions (AVX) for 3D reconstruction. J. Struct. Biol. 189, 147–152 (2015).
Tegunov, D. & Cramer, P. Real-time cryo-electron microscopy data preprocessing with Warp. Nat. Methods 16, 1146–1152 (2019).
Liu, Y.-T. et al. Isotropic reconstruction of electron tomograms with deep learning. Preprint at bioRxiv https://doi.org/10.1101/2021.07.17.452128 (2021).
Kremer, J. R., Mastronarde, D. N. & McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. J. Struct. Biol. 116, 71–76 (1996).
Zheng, S. et al. AreTomo: an integrated software package for automated marker-free, motion-corrected cryo-electron tomographic alignment and reconstruction. J. Struct. Biol. X 6, 100068 (2022).
Wagner, T. et al. SPHIRE-crYOLO is a fast and accurate fully automated particle picker for cryo-EM. Commun. Biol. 2, 218 (2019).
Zivanov, J. et al. A Bayesian approach to single-particle electron cryo-tomography in RELION-4.0. Preprint at bioRxiv https://doi.org/10.1101/2022.02.28.482229 (2022).
Kimanius, D., Dong, L., Sharov, G., Nakane, T. & Scheres, S. H. W. New tools for automated cryo-EM single-particle analysis in RELION-4.0. Biochem. J. 478, 4169–4185 (2021).
Acknowledgements
Cryo-EM data were collected at Columbia Cryo-EM facility and at the Simons Electron Microscopy Center (SEMC), directed by B. Carragher and C. Potter, with the assistance of staff from both SEMC and the Columbia University Cryo-Electron Microscopy Center. R. Grassucci and Z. Zhang from the Columbia Cryo-EM Center assisted with data collection. Some of the work was performed using equipment from the Center for Membrane Protein Production and Analysis (COMPPÅ; grant no. NIH P41 GM116799 to W.A. Hendrickson). We thank L. Maso for generating the schematic representation of the ankyrin-1 complex (Fig. 1c) and F. Mancia and his laboratory members for support and critical reading of the manuscript. We thank G. Gillis for assisting during protein purification. We are grateful to P. Berto and L. Barazzuol, Department of Biomedical Sciences, University of Padova, for their support in the preparation of the ghost membranes. We thank the ‘Centro Transfusionale dell’Azienda Ospedaliera Università di Padova’ and the individuals who donated their blood from which the ankyrin complex was purified. T.C. is supported by grants from the Italian Ministry of University and Research (Bando SIR 2014 grant nos. RBSI14C65Z and PRIN2017) and from the Universita’ degli Studi di Padova (Progetto Giovani 2012 grant no. GRIC128SP0, Progetto di Ateneo 2016 no. CALI_SID16_01 and STARS consolidator grant no. 2019). Some of this work was performed at the SEMC, National Resource for Automated Molecular Microscopy and the National Center for In situ Tomographic Ultramicroscopy located at the New York Structural Biology Center, supported by grants from the Simons Foundation (no. SF349247) and NIH NIGMS (no. GM103310) and NIH (no. U24GM139171).
Author information
Authors and Affiliations
Contributions
F.V., with the assistance of L.Y.Y., performed protein preparation for structural analysis. F.V. and T.C. prepared membranes from human erythrocytes. F.V. screened and optimized sample vitrification, and generated cryo-EM data. J.D.J. and A.J.N. carried out tomography data analysis. Single-particle cryo-EM data analysis and model building was conducted by O.B.C. The manuscript was written by O.B.C. and F.V. with input from T.C. Figures were prepared by K.K and F.V.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Structural & Molecular Biology thanks Yifan Cheng, Werner Kühlbrandt and Ashley Toye for their contribution to the peer review of this work. Primary Handling editor: Florian Ullrich, in collaboration with the Nature Structural & Molecular Biology team. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Single particle cryo-EM structure determination.
a, A representative micrograph out of 14.464 micrographs. b, 2D class averages of particles initially picked with Topaz (~1.8 M), ordered by population. 2D class averages of band 3–GPA (red box) and ankyrin-1 complexes (green box) are highlighted. c, Fourier shell correlation (FSC) curves. Maps shown as inset, with refinement mask in the case of local refinements.
Extended Data Fig. 2 Cryo-EM workflow and analysis of the ankyrin-1 complex.
Flowchart outlining cryo-EM image acquisition and processing performed to obtain the structure of ankyrin-1 complex. All processing was performed using CryoSPARC v.3.3 (see Methods for details).
Extended Data Fig. 3 Model/map fit.
Cryo-EM densities (transparent gray surface) are shown with corresponding segments of the atomic model; sidechains are rendered in stick representation and colored as in Fig. 1. The densities for GPA and free band 3 are derived from the free band3 class. Density for band 3-I TM derives from local refinement of Class 3. For all the other images we used the density from local refinement of the consensus refinement.
Extended Data Fig. 4 Ankyrin-1 complex classes.
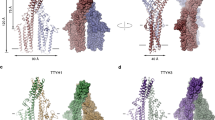
a, Different views of six main classes of the ankyrin-1 complex. Class 1 contains an (RhAG)2(RhCE) heterotrimer, ankyrin-1, protein 4.2, and three band 3 dimers (I, II, III). Class 2 exhibits a smaller micelle, retaining the core (RhAG)2(RhCE)(Ank1)(P4.2) architecture, but including only one band 3 dimer (B3-I), bound to protein 4.2, and aquaporin. Class 3 contains an (RhAG)2(RhCE) heterotrimer, and a single band 3 (B3-I). Class 4 has a bigger micelle and contains an (RhAG)2(RhCE) heterotrimer, ankyrin-1, protein 4.2, three band 3 dimers and an unidentified protein “X’. Class 5 contains an (RhAG)2(RhCE) heterotrimer, ankyrin-1, protein 4.2, aquaporin and Band 3-I and Band 3-III. Class 6 contains an (RhAG)2(RhCE) heterotrimer, ankyrin-1, protein 4.2, and Band 3-I and Band 3-III. In all the six classes the cytosolic domain of Band 3-II is present. The transmembrane domains of B3-II in classes 2, 3 and 6 appear to be associated with a separate micelle, likely due to either partial dissociation during extraction and/or re-association with free band 3 dimers post-extraction. b, Comparison of Class 1 & Class 2. Rh is colored in blue, Band 3-I in orange, Band 3-II in lilac, Band 3-III in yellow and aquaporin in pink. Superposition of Class 1 and Class 2 shows that presence of the additional two band 3 dimers does not exclude the presence of aquaporin.
Extended Data Fig. 5 Cryo-ET of native membrane vesicles.
a, A composite slice from the tomogram shown in Supplementary Video 2 with several densities resembling ankyrin-1 complex side-views boxed out. b, SPA 2D class side-views for comparison with the tomogram slices. c, Magnified tomogram slices from the boxed-out regions in A. The multi-complex 2D slice on the right shows one of many visible strings of ankyrin-1 complex-like densities suggesting potential higher order assemblies. Tomogram slices are 8.3 Å thick. During the process of vesicle formation, vesicles can form in either the native or inverted configurations, with the cytosolic side of the membrane oriented in either direction.
Extended Data Fig. 6 Cryo-EM workflow for sub-classification of Band 3-I from the consensus refinement.
Flowchart outlining cryo-EM processing performed to improve the structure of Band 3-I (orange) and the bound GPA proteins (magenta). All processing was performed using CryoSPARC v.3.3. A similar procedure was used to improve the density of AQP1 starting from Class 2.
Extended Data Fig. 7 Conformation of membrane-associated ankyrin-1.
a, Ankyrin structure shown as a molecular surface with the inner groove depicted in pink, the convex outer surface in red, in yellow the N-terminal peptide of Band 3-III and in magenta the C-terminal linker of ankyrin-1. b, Structural alignment between ankyrin-1 structure (colored by RMSD) and ankyrin-2 (in gray). The first 5 repeats of Ank1 are dramatically rearranged compared to their position in the Ank2 structure. Arrows indicate AR5 and 6. c, Structural alignment between ankyrin-2 (gray) and ankyrin-2 bound to Nav1.2 peptide (colored by RMSD), shows that flexibility at this interface also occurs in ankyrin-2. d, Hydrophobic surface calculated for the first 5 ankyrin repeats (AR1-5) of Ank1 (top) and Ank2 (bottom). RhCE N-terminal (magenta) and C-terminal fragments (purple) are displayed in stick representation. In the lower figure Ank2 is bound to Nav1.2 peptide (blue). e, Sequence alignment of all the 24 ankyrin repeats of Ank1 using MUSCLE82 and visualized and colored using Jalview with the ClustalX color scheme. The sequences for AR5 and 6 are outlined in red. f, Sequence of AR5 and AR6 from Ank1, Ank2 and Ank3, aligned using MUSCLE 82 and visualized and colored using Jalview with the ClustalX colour scheme. The three sequences are well conserved in AR5 and 6. Arrows indicate the region that rearranges in Ank1. AR5 is shorter than the other repeats by 4 residues, and AR6 lacks both the “S/TP’ motif at positions 4–5, and the consensus “GH’ motif at positions 13–14 is replaced by two aspartates.
Extended Data Fig. 8 Ankyrin interaction with Band 3-III N-terminal peptide.
a, Map at low density threshold allows tracing of the complete Band 3-III N-terminal peptide (dotted yellow line), that runs back along the inner ankyrin groove. b-c, Molecular surface of the ankyrin repeats, colored by hydrophobicity, bound to the N-terminal peptide of Band 3-III (yellow). Residues 2–24 of Band 3-III form an ordered interaction with AR6-10 and the AR5-6 linker. Key residues are displayed as sticks. Band 3-III is shown as yellow ribbons. d, Structural comparison between ankyrin-1 (top) and ankyrin-2 structures (PDB: 4Y4D) (bottom). Structure of ankyrin-1 (dark red AR1-5 and red AR6-12) is associated with N-terminal peptide of Band 3-I (yellow) and ankyrin-2 (dark blue AR1-6 and light blue AR6-12) is associated with the autoinhibitory domain (AI) of ankyrin-1. e, Close up of the AR5-6 of ankyrin-1 in panel S8d. The Band 3-I part colored in green corresponds to the first 11 N-terminal amino acids. f, Structural alignment between the first five ankyrin repeats (AR1-5) of ankyrin-1 (dark red) bound to RhCE peptide and ankyrin-2 (dark blue) bound to the autoinhibitory peptide of ankyrin-1. The key conserved residues that mediate the interactions between ANK1 and B3-III and ANK2 and the autoinhibitory region are shown as sticks.
Extended Data Fig. 9 Flexibility of band 3 cytosolic domain.
a, Models of the three Band 3 dimers (I, II, III). The three cytosolic domains of Band 3 proteins in the complex have different orientations. The Band 3-I cytoplasmic domain is inverted with respect to those of Band 3-II and Band 3-III. The TM part of Band 3 is colored gray. b, Two different views of band 3 cytosolic domain represented as molecular surface. Interacting surfaces of band 3 are distinct and non-overlapping. Regions of the band 3 cytosolic domain that interact with proteins in the complex are shown in different colors: green for the region that interact with protein 4.2, red for ankyrin repeats 22–24 and purple for ankyrin repeats 17–19. c, Comparison between Band 3-I (yellow) associated with protein 4.2 (green) and Band 3-II (lilac). The model is displayed as ribbon and the surface is shown in transparency. In the right panel a close view of the interactions between Band 3-II cytosolic domain (lilac) and protein 4.2 (green); the two share the same interaction site with the TM part. Key residues for the interaction are displayed as sticks.
Extended Data Fig. 10 (RhAG)2(RhCE) trimer interactions.
a, Model of AQP1 with waters (red dots fit inside the map). In the bottom inset a close view of the waters in the central cavity. b, Unmodeled density at the interface of aquaporin-1 (salmon) with RhCE (purple) and RhAG (light blue). Potential interacting sidechains are depicted in stick representation. c, The structure of (RhAG)2(RhCE) trimer as viewed from the cytoplasm, RhCE is colored in purple, RhAG1 in light blue and to RhAG2 in dark blue. The cryo-EM density map of the (RhAG)2(RhCE) trimer is shown in transparency. The 12 TM helixes are numbered (0–11) for each Rh molecule. Gray circles shown the pore position in each subunit. d, Interfaces between the three different subunits, with key residues represented as sticks. e, Interface between the three Rh protomers. The key residues that mediate the interactions in the interfaces are shown as sticks. Three different parts of the trimerization interface are show in the right figure viewed from the top. f, Analysis of the pores of RhCE and RhAG. Figure of the pore in green generated by HOLE. 2D graph of pore radius for RhCE (top graph) and RhAG (bottom graph). h, Close up of RhAG2, the map shows an undefine density. The key residues that interact with the undefined density are shown as sticks. g, Structure of (RhAG)2(RhCE) complex and the first five repeats of ankyrin. The amino acids depicted in yellow sticks highlight the sites of variation between RhCE and RhD. Three sites of variation between RhD and RhCE are shown in the right panels, with the density map overlayed. The density map is consistent with the presence of RhCE, not RhD.
Supplementary information
Supplementary Materials
Supplementary Figs. 1–4, Table 1 and captions for Videos 1–6.
Supplementary Video 1
Class 1 overview. An overview of the assembly of the ankyrin-1 complex (Class 1) is shown, with both atomic model and map depicted.
Supplementary Video 2
Cryo-ET of native membrane vesicles. A composite slice from the tomogram with several densities resembling ankyrin-1 complex side-views boxed out. The multi-complex 2D slice on the right shows one of many visible strings of ankyrin-1 complex-like densities suggesting potential higher order assemblies. Tomogram slices are 8.3 Å thick.
Supplementary Video 3
Sub-tomogram average of particles extracted from native membrane vesicles. The map generated from 1500 sub-tomograms of the ankyrin complex is shown, together with an atomic model generated by aligning and combining Classes 1 & 2.
Supplementary Video 4
Class 2 overview. An overview of the assembly of Class 2 is shown, with both atomic model and map depicted.
Supplementary Video 5
N-terminal peptide of B3-III. The N-terminal peptide of Band 3-III (yellow cartoon) is shown in the groove of ankyrin-1 (red surface), followed by a zoom in on the fit of the N-terminal peptide of Band 3-III to the density map.
Supplementary Video 6
PIP2 model/map fit. The fit of the PIP2 model to the band 3-GPA dimer map is shown.
Rights and permissions
About this article

Cite this article
Vallese, F., Kim, K., Yen, L.Y. et al. Architecture of the human erythrocyte ankyrin-1 complex. Nat Struct Mol Biol 29, 706–718 (2022). https://doi.org/10.1038/s41594-022-00792-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41594-022-00792-w
This article is cited by
-
Bridging structural and cell biology with cryo-electron microscopy
Nature (2024)
-
Structural and functional insights into the lipid regulation of human anion exchanger 2
Nature Communications (2024)
-
Mechanism of anion exchange and small-molecule inhibition of pendrin
Nature Communications (2024)
-
Future opportunities in solute carrier structural biology
Nature Structural & Molecular Biology (2024)
-
Substrate binding and inhibition of the anion exchanger 1 transporter
Nature Structural & Molecular Biology (2023)
