
Abstract
Vertebrates use the mannose 6-phosphate (M6P)-recognition system to deliver lysosomal hydrolases to lysosomes. Key to this pathway is N-acetylglucosamine (GlcNAc)-1-phosphotransferase (PTase) that selectively adds GlcNAc-phosphate (P) to mannose residues of hydrolases. Human PTase is an α2β2γ2 heterohexamer with a catalytic core and several peripheral domains that recognize and bind substrates. Here we report a cryo-EM structure of the catalytic core of human PTase and the identification of a hockey stick-like motif that controls activation of the enzyme. Movement of this motif out of the catalytic pocket is associated with a rearrangement of part of the peripheral domains that unblocks hydrolase glycan access to the catalytic site, thereby activating PTase. We propose that PTase fluctuates between inactive and active states in solution, and selective substrate binding of a lysosomal hydrolase through its protein-binding determinant to PTase locks the enzyme in the active state to permit glycan phosphorylation. This mechanism would help ensure that only N-linked glycans of lysosomal enzymes are phosphorylated.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
Data availability
The 3D cryo-EM maps of the human PTase catalytic core in the active (3.1 Å) and auto-inhibited (3.3 Å) state have been deposited in the EMDB under accession codes 24784 and 24785, respectively. The atomic model based on the two 3D maps has been deposited in the PDB under accession codes 7S05 and 7S06, respectively. Source data are provided with this paper.
References
Braulke, T. & Bonifacino, J. S. Sorting of lysosomal proteins. Biochim. Biophys. Acta 1793, 605–614 (2009).
Burda, P. & Aebi, M. The dolichol pathway of N-linked glycosylation. Biochim. Biophys. Acta 1426, 239–257 (1999).
Bai, L. & Li, H. Cryo-EM is uncovering the mechanism of eukaryotic protein N-glycosylation. FEBS J. 286, 1638–1644 (2019).
Kudo, M. et al. The α- and β-subunits of the human UDP-N-acetylglucosamine:lysosomal enzyme N-acetylglucosamine-1-phosphotransferase [corrected] are encoded by a single cDNA. J. Biol. Chem. 280, 36141–36149 (2005).
Raas-Rothschild, A. et al. Molecular basis of variant pseudo-Hurler polydystrophy (mucolipidosis IIIC). J. Clin. Invest. 105, 673–681 (2000).
Sperisen, P., Schmid, C. D., Bucher, P. & Zilian, O. Stealth proteins: in silico identification of a novel protein family rendering bacterial pathogens invisible to host immune defense. PLoS Comput. Biol. 1, e63 (2005).
Qian, Y. et al. Functions of the α, β, and γ subunits of UDP-GlcNAc:lysosomal enzyme N-acetylglucosamine-1-phosphotransferase. J. Biol. Chem. 285, 3360–3370 (2010).
Marschner, K., Kollmann, K., Schweizer, M., Braulke, T. & Pohl, S. A key enzyme in the biogenesis of lysosomes is a protease that regulates cholesterol metabolism. Science 333, 87–90 (2011).
Braulke, T., Pohl, S. & Storch, S. Molecular analysis of the GlcNac-1-phosphotransferase. J. Inherit. Metab. Dis. 31, 253–257 (2008).
Qian, Y. et al. Analysis of mucolipidosis II/III GNPTAB missense mutations identifies domains of UDP-GlcNAc:lysosomal enzyme GlcNAc-1-phosphotransferase involved in catalytic function and lysosomal enzyme recognition. J. Biol. Chem. 290, 3045–3056 (2015).
Liu, L., Lee, W. S., Doray, B. & Kornfeld, S. Role of spacer-1 in the maturation and function of GlcNAc-1-phosphotransferase. FEBS Lett. 591, 47–55 (2017).
Qian, Y., Flanagan-Steet, H., van Meel, E., Steet, R. & Kornfeld, S. A. The DMAP interaction domain of UDP-GlcNAc:lysosomal enzyme N-acetylglucosamine-1-phosphotransferase is a substrate recognition module. Proc. Natl Acad. Sci. USA 110, 10246–10251 (2013).
van Meel, E. et al. Multiple domains of GlcNAc-1-phosphotransferase mediate recognition of lysosomal enzymes. J. Biol. Chem. 291, 8295–8307 (2016).
De Pace, R. et al. Subunit interactions of the disease-related hexameric GlcNAc-1-phosphotransferase complex. Hum. Mol. Genet. 24, 6826–6835 (2015).
Velho, R. V., De Pace, R., Tidow, H., Braulke, T. & Pohl, S. Identification of the interaction domains between α- and γ-subunits of GlcNAc-1-phosphotransferase. FEBS Lett. 590, 4287–4295 (2016).
Liu, L., Lee, W. S., Doray, B. & Kornfeld, S. Engineering of GlcNAc-1-phosphotransferase for production of highly phosphorylated lysosomal enzymes for enzyme replacement therapy. Mol. Ther. Methods Clin. Dev. 5, 59–65 (2017).
Boustany, R. M. Lysosomal storage diseases—the horizon expands. Nat. Rev. Neurol. 9, 583–598 (2013).
Velho, R. V. et al. The lysosomal storage disorders mucolipidosis type II, type III α/β, and type III γ: update on GNPTAB and GNPTG mutations. Hum. Mutat. 40, 842–864 (2019).
Appelqvist, H., Waster, P., Kagedal, K. & Ollinger, K. The lysosome: from waste bag to potential therapeutic target. J. Mol. Cell Biol. 5, 214–226 (2013).
Marques, A. R. A. & Saftig, P. Lysosomal storage disorders—challenges, concepts and avenues for therapy: beyond rare diseases. J Cell Sci. 132, jcs221739 (2019).
Kudo, M. & Canfield, W. M. Structural requirements for efficient processing and activation of recombinant human UDP-N-acetylglucosamine:lysosomal-enzyme-N-acetylglucosamine-1-phosphotransferase. J. Biol. Chem. 281, 11761–11768 (2006).
Tiede, S. et al. Mucolipidosis II is caused by mutations in GNPTA encoding the α/β GlcNAc-1-phosphotransferase. Nat. Med. 11, 1109–1112 (2005).
Wang, Y. et al. Identification of predominant GNPTAB gene mutations in Eastern Chinese patients with mucolipidosis II/III and a prenatal diagnosis of mucolipidosis II. Acta Pharmacol. Sin. 40, 279–287 (2019).
Pedersen, L. C. et al. Crystal structure of an α1,4-N-acetylhexosaminyltransferase (EXTL2), a member of the exostosin gene family involved in heparan sulfate biosynthesis. J. Biol. Chem. 278, 14420–14428 (2003).
Kitagawa, H., Shimakawa, H. & Sugahara, K. The tumor suppressor EXT-like gene EXTL2 encodes an α1, 4-N-acetylhexosaminyltransferase that transfers N-acetylgalactosamine and N-acetylglucosamine to the common glycosaminoglycan–protein linkage region. The key enzyme for the chain initiation of heparan sulfate. J. Biol. Chem. 274, 13933–13937 (1999).
Tiede, S. et al. Missense mutations in N-acetylglucosamine-1-phosphotransferase α/β subunit gene in a patient with mucolipidosis III and a mild clinical phenotype. Am. J. Med. Genet. A 137A, 235–240 (2005).
Punjani, A. & Fleet, D. J. 3D variability analysis: resolving continuous flexibility and discrete heterogeneity from single particle cryo-EM. J. Struct. Biol. 213, 107702 (2021).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife 7, e42166 (2018).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486–501 (2010).
Buchan, D. W. A. & Jones, D. T. The PSIPRED Protein Analysis Workbench: 20 years on. Nucleic Acids Res. 47, W402–W407 (2019).
Liebschner, D. et al. Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Acta Crystallogr. D Struct. Biol. 75, 861–877 (2019).
Williams, C. J. et al. MolProbity: more and better reference data for improved all-atom structure validation. Protein Sci. 27, 293–315 (2018).
Robert, X. & Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 42, W320–W324 (2014).
Goddard, T. D. et al. UCSF ChimeraX: meeting modern challenges in visualization and analysis. Protein Sci. 27, 14–25 (2017).
Acknowledgements
Cryo-EM images were collected in the David Van Andel Advanced Cryo-Electron Microscopy Suite at the Van Andel Institute. We thank G. Zhao and X. Meng for facilitating data collection. This work was supported by the US National Institutes of Health (CA008759 to S.K. and CA231466 to Huilin Li) and grants from the Yash Gandhi Foundation (to S.K.) and the Van Andel Institute (to Huilin Li). Raw data and materials generated in this study are freely available upon request from the corresponding authors.
Author information
Authors and Affiliations
Contributions
S.K. and Huilin Li conceived and designed experiments. W.-S.L., B.D. and W.M.C. prepared samples. Hua Li and L.B. performed cryo-EM structural analysis. Hua Li and X.F. built the atomic model. W.-S.L. and B.D. generated mutant constructs, and W.-S.L. carried out enzyme activity assays. B.C.J. acquired confocal images, and W.-S.L. and L.L. performed co-immunoprecipitation experiments. Hua Li, S.K., B.D. and Huilin Li analyzed data and wrote the manuscript with input from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Structural and Molecular Biology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: Florian Ullrich, in collaboration with the Nature Structural and Molecular Biology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Workflow for cryo-EM structure determination.
a) A typical raw micrograph. A total of 13,320 micrographs were recorded. b) Workflow of cryo-EM data processing. c) Selected 2D averages out of a total of 34 class averages. d, g) Gold-standard Fourier shell correlations; e, h) Eulerian angle distribution of particles used for 3D reconstruction. f, i) Local resolution maps in the active and autoinhibited states.
Extended Data Fig. 2 Cryo-EM 3D map of the core region of the human PTase.
Top and bottom views of the density map with the NTE, CR1-4, EF hand, S4, CTE, and the two resolved helices in S1 and S3 in subunit A colored in blue, magenta, olive drab, yellow, dark orange, purple, wheat, pink, and gray, respectively. Subunit B is in cyan.
Extended Data Fig. 3 EM density superimposed with the atomic model.
a) EM density selected from each main region. b) EM densities for four of the five identified N-glycans. The density for other N-glycan on Asn-83 is shown in the first panel of (a). c) EM density of the autoinhibitory hockey-stick motif, which is the first α-helix of S3 domain (S3-H1).
Extended Data Fig. 4 Topology of PTase αβ monomer.
a) Monomer structure in cartoon with the three main regions highlighted in circles. b) Topological plot of the secondary structures. The three main regions are included in dashed shapes. Shown are the NTE, the large lobe, and the small lobe of a PTase monomer. NTE, CR1-4, EF hand, S4, and CTE are colored in blue, magenta, olive drab, yellow, dark orange, and purple, wheat, pink, respectively.
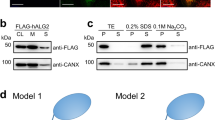
Extended Data Fig. 5 Effect of dimerization and interface mutations on PTase activity.
a) Immunofluorescence microscopy of transfected HeLa cells showing Golgi localization of WT PTase and the C70S mutant. b) Effect of C70S on the ability of PTase ΔS1-S3 to form a non-covalent dimer, as assessed by co-immunoprecipitation. Cell lysates from HEK 293 cells transfected with the indicated plasmids were immunoprecipitated with an anti-V5 rabbit monoclonal antibody, and the pellet fractions (10% loaded) were immunoblotted with an anti-FLAG mouse monoclonal antibody. For the input, only the immunoblot probed with the anti-FLAG antibody is shown here. The expression of ΔS1-S3-V5 in the input was ascertained by probing a separate blot with the anti-V5 rabbit monoclonal antibody (not shown). c) Effect of C70S on the ΔS1-S3 α-methylmannoside activity. For the activity assay, ΔS1-S3 and ΔS1-S3(C70S) were expressed in HEK 293 cells. The data shown are the mean of 2 independent experiments. d) Immunofluorescence microscopy of transfected HeLa cells shows the various interface mutants to be localized to the Golgi similar to WT PTase. e) Immunoblot of WT PTase and the interface mutants expressed in GNPTAB−/− HeLa cells, and probed with a monoclonal antibody against the V5 tag. 25 µg of each cell lysate was loaded. f) α-methylmannoside activity of WT PTase and the interphase mutants expressed in GNPTAB−/− HeLa cells. The data shown are the mean ± SD for 3 independent experiments. Uncropped images for b,e and data for graphs in c,f are available as source data.
Extended Data Fig. 6 Effects of mutations in the dimer interface at the EF hand and CR4 region.
a) Immunoblot of WT PTase, D1018V, L1025S and L1033P expressed in GNPTAB−/− HeLa cells and probed with a monoclonal antibody against the V5 tag. 25 µg of each cell lysate was loaded. b) Immunofluorescence microscopy of transfected HeLa cells shows the R1028A, R1031A, and R1028A/R1031A mutants to be localized to the Golgi similar to WT PTase. c) Immunoblot of WT PTase, R1028A, R1031A, and R1028A/R1031A expressed in GNPTAB−/− HeLa cells and probed with a monoclonal antibody against the V5 tag. 25 µg of each cell lysate was loaded. d) α-methylmannoside activity of WT PTase, R1028A, R1031A, and R1028A/R1031A expressed in GNPTAB−/− HeLa cells. The data shown are the mean ± SD for 3 independent experiments. Uncropped images for a,c and data for graph in d are available as source data.
Extended Data Fig. 7 Effects of catalytic site mutations.
a) Immunofluorescence microscopy of transfected HeLa cells shows the N406A and D408A mutants to be localized to the Golgi similar to WT PTase. b) Immunoblot of WT PTase, N406A and D408A expressed in GNPTAB−/− HeLa cells and probed with a monoclonal antibody against the V5 tag. 25 µg of each cell lysate was loaded. c) α-methylmannoside activity of WT PTase, N406A and D408A expressed in GNPTAB−/− HeLa cells. The data shown are the mean ± SD for 2 independent experiments. Uncropped images for b and data for graph in c are available as source data.
Extended Data Fig. 8 The S1 domain structure.
a) Sub domain organization of the human PTase S1 domain. S1-H1 is the first α-helix resolved in this study upstream of the S1 core. Structure in the dash rectangle, is currently unknown. b) Two orthogonal cartoon views of the S1 core structure solved previously by the Kurt Wuthrich lab and Joint Center for Structural Genomics using NMR method (PDB ID 2N6D). The S1 core has a saddle-like shape with a pseudo 2-fold symmetry. Each subdomain contains three α-helices and four β-strands and the two subdomains are connected by a bridge of two β-strands. c-d) The S1 core docked into the cryo-EM map of the S1 region in the autoinhibited (c) and active state (d).
Extended Data Fig. 9 Mutations mapped on the core structure of PTase.
a) Reported human disease related mutations (Velho et al. Hum Mutat 2019, 40, 842-864). b) Mutations examined in this study. Amino acid identity is labeled only for one of the two protomers.
Supplementary information
Supplementary Information
Supplementary Fig. 1 and Table 1
Supplementary Video 1
Dynamics in the PTase core region near the catalytic pocket. The video first shows a rotation of the 3D map of the core region in the auto-inhibited state superimposed with the atomic model and then zooms into the catalytic site. Next, the scene changes to 3DVA analysis results, which are composed of 20 3D frames (maps) revealing EM density changes as the structure transitions from the auto-inhibited state to the active state.
Supplementary Video 2
Dynamics in the PTase α2β2 complex. A low-resolution 3DVA analysis of the full structure, revealing large-scale movement in the S1 and N1–S3 regions as the structure transitions from the auto-inhibited state to the active state.
Source data
Source Data Fig. 2
Unprocessed western blots for Fig. 2c and statistical source data for Fig. 2d,g.
Source Data Fig. 3
Unprocessed western blots and statistical source data for Fig. 3e.
Source Data Extended Data Fig. 5
Unprocessed western blots for Fig. 5b,e and statistical source data for Fig. 5c,f.
Source Data Extended Data Fig. 6
Unprocessed western blots for Fig. 6a,c and statistical source data for Fig. 6d.
Source Data Extended Data Fig. 7
Unprocessed western blots for Fig. 7b and statistical source data for Fig. 7c.
Rights and permissions
About this article

Cite this article
Li, H., Lee, WS., Feng, X. et al. Structure of the human GlcNAc-1-phosphotransferase αβ subunits reveals regulatory mechanism for lysosomal enzyme glycan phosphorylation. Nat Struct Mol Biol 29, 348–356 (2022). https://doi.org/10.1038/s41594-022-00748-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41594-022-00748-0
This article is cited by
-
Lysosomes as coordinators of cellular catabolism, metabolic signalling and organ physiology
Nature Reviews Molecular Cell Biology (2024)
-
GCAF(TMEM251) regulates lysosome biogenesis by activating the mannose-6-phosphate pathway
Nature Communications (2022)
