
Abstract
Urination (also called micturition) is thought to be regulated by a neural network that is distributed in both subcortical and cortical regions. Previously, urination-related neurons have been identified in subcortical structures such as the pontine micturition center (also known as Barrington’s nucleus). However, the origin of the descending cortical pathway and how it interfaces with this subcortical circuit to permit voluntary initiation of urination remain elusive. Here we identified a small cluster of layer 5 neurons in the primary motor cortex whose activities tightly correlate with the onset of urination in freely behaving mice and increase dramatically during territorial marking. Optogenetically activating these neurons elicits contraction of the bladder and initiates urination, through their projections to the pontine micturition center, while silencing or ablating them impairs urination and causes retention of urine. Together these results reveal a novel cortical component upstream of the pontine micturition center that is critically involved in urination.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Data and code availability
The data that support the findings of this study are available from the corresponding author upon reasonable request. All software code used to generate the results is also available from the corresponding author upon reasonable request.
References
Fowler, C. J., Griffiths, D. & de Groat, W. C. The neural control of micturition. Nat. Rev. Neurosci. 9, 453–466 (2008).
Benarroch, E. E. Neural control of the bladder: recent advances and neurologic implications. Neurology 75, 1839–1846 (2010).
Griffiths, D. Neural control of micturition in humans: a working model. Nat. Rev. Urol. 12, 695–705 (2015).
de Groat, W. C., Griffiths, D. & Yoshimura, N. Neural control of the lower urinary tract. Compr. Physiol. 5, 327–396 (2015).
Valentino, R. J., Wood, S. K., Wein, A. J. & Zderic, S. A. The bladder-brain connection: putative role of corticotropin-releasing factor. Nat. Rev. Urol. 8, 19–28 (2011).
Kitta, T. et al. Brain-bladder control network: the unsolved 21st century urological mystery. Int. J. Urol. 22, 342–348 (2015).
Andersson, K. E. & Arner, A. Urinary bladder contraction and relaxation: physiology and pathophysiology. Physiol. Rev. 84, 935–986 (2004).
Griffiths, D. J. & Fowler, C. J. The micturition switch and its forebrain influences. Acta Physiol. (Oxf). 207, 93–109 (2013).
Yoshiyama, M. et al. Functional roles of TRPV1 and TRPV4 in control of lower urinary tract activity: dual analysis of behavior and reflex during the micturition cycle. Am. J. Physiol. Renal. Physiol. 308, F1128–F1134 (2015).
Manohar, A., Curtis, A. L., Zderic, S. A. & Valentino, R. J. Brainstem network dynamics underlying the encoding of bladder information. eLife 6, e29917 (2017).
Griffiths, D. J. Cerebral control of bladder function. Curr. Urol. Rep. 5, 348–352 (2004).
de Groat, W. C. & Wickens, C. Organization of the neural switching circuitry underlying reflex micturition. Acta Physiol. (Oxf). 207, 66–84 (2013).
Hou, X. H. et al. Central control circuit for context-dependent micturition. Cell 167, 73–86. e12 (2016).
Keller, J. A. et al. Voluntary urination control by brainstem neurons that relax the urethral sphincter. Nat. Neurosci. 21, 1229–1238 (2018).
Fowler, C. J. & Griffiths, D. J. A decade of functional brain imaging applied to bladder control. Neurourol. Urodyn. 29, 49–55 (2010).
Nour, S., Svarer, C., Kristensen, J. K., Paulson, O. B. & Law, I. Cerebral activation during micturition in normal men. Brain 123, 781–789 (2000).
Michels, L. et al. Supraspinal control of urine storage and micturition in men–an fMRI study. Cereb. Cortex 25, 3369–3380 (2015).
Nadelhaft, I. & Vera, P. L. Neurons in the rat brain and spinal cord labeled after pseudorabies virus injected into the external urethral sphincter. J. Comp. Neurol. 375, 502–517 (1996).
Grill, W. M., Erokwu, B. O., Hadziefendic, S. & Haxhiu, M. A. Extended survival time following pseudorabies virus injection labels the suprapontine neural network controlling the bladder and urethra in the rat. Neurosci. Lett. 270, 63–66 (1999).
Stanley, S. et al. Identification of neuronal subpopulations that project from hypothalamus to both liver and adipose tissue polysynaptically. Proc. Natl Acad. Sci. USA 107, 7024–7029 (2010).
Smith, B. N. et al. Pseudorabies virus expressing enhanced green fluorescent protein: A tool for in vitro electrophysiological analysis of transsynaptically labeled neurons in identified central nervous system circuits. Proc. Natl Acad. Sci. USA 97, 9264–9269 (2000).
Zermann, D. H., Ishigooka, M., Doggweiler, R. & Schmidt, R. A. Central autonomic innervation of the lower urinary tract--a neuroanatomy study. World J. Urol. 16, 417–422 (1998).
Yoshimura, N. Bladder afferent pathway and spinal cord injury: possible mechanisms inducing hyperreflexia of the urinary bladder. Prog. Neurobiol. 57, 583–606 (1999).
Li, A. et al. Micro-optical sectioning tomography to obtain a high-resolution atlas of the mouse brain. Science 330, 1404–1408 (2010).
Gong, H. et al. High-throughput dual-colour precision imaging for brain-wide connectome with cytoarchitectonic landmarks at the cellular level. Nat. Commun. 7, 12142 (2016).
Stroh, A. et al. Making waves: initiation and propagation of corticothalamic Ca2+ waves in vivo. Neuron 77, 1136–1150 (2013).
Zhang, Q. et al. Locomotion-related population cortical Ca2+ transients in freely behaving mice. Front. Neural Circuits 11, 24 (2017).
Chen, T. W. et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 499, 295–300 (2013).
Granstedt, A. E., Szpara, M. L., Kuhn, B., Wang, S. S. & Enquist, L. W. Fluorescence-based monitoring of in vivo neural activity using a circuit-tracing pseudorabies virus. PLoS One 4, e6923 (2009).
Granstedt, A. E., Bosse, J. B., Thiberge, S. Y. & Enquist, L. W. In vivo imaging of alphaherpesvirus infection reveals synchronized activity dependent on axonal sorting of viral proteins. Proc. Natl Acad. Sci. USA 110, E3516–E3525 (2013).
Beier, K. T. et al. Anterograde or retrograde transsynaptic labeling of CNS neurons with vesicular stomatitis virus vectors. Proc. Natl Acad. Sci. USA 108, 15414–15419 (2011).
Zingg, B. et al. AAV-mediated anterograde transsynaptic tagging: mapping corticocollicular input-defined neural pathways for defense behaviors. Neuron 93, 33–47 (2017).
Atasoy, D., Betley, J. N., Su, H. H. & Sternson, S. M. Deconstruction of a neural circuit for hunger. Nature 488, 172–177 (2012).
Mahler, S. V. et al. Designer receptors show role for ventral pallidum input to ventral tegmental area in cocaine seeking. Nat. Neurosci. 17, 577–585 (2014).
Yang, C. F. et al. Sexually dimorphic neurons in the ventromedial hypothalamus govern mating in both sexes and aggression in males. Cell 153, 896–909 (2013).
Jennings, J. H. et al. Visualizing hypothalamic network dynamics for appetitive and consummatory behaviors. Cell 160, 516–527 (2015).
Holstege, G. Micturition and the soul. J. Comp. Neurol. 493, 15–20 (2005).
Blok, B. F., De Weerd, H. & Holstege, G. Ultrastructural evidence for a paucity of projections from the lumbosacral cord to the pontine micturition center or M-region in the cat: a new concept for the organization of the micturition reflex with the periaqueductal gray as central relay. J. Comp. Neurol. 359, 300–309 (1995).
Li, J. et al. Primary auditory cortex is required for anticipatory motor response. Cereb. Cortex 27, 3254–3271 (2017).
Stosiek, C., Garaschuk, O., Holthoff, K. & Konnerth, A. In vivo two-photon calcium imaging of neuronal networks. Proc. Natl Acad. Sci. USA 100, 7319–7324 (2003).
Adelsberger, H., Garaschuk, O. & Konnerth, A. Cortical calcium waves in resting newborn mice. Nat. Neurosci. 8, 988–990 (2005).
Grienberger, C. et al. Sound-evoked network calcium transients in mouse auditory cortex in vivo. J. Physiol. (Lond.) 590, 899–918 (2012).
Diehl, K. H. et al. A good practice guide to the administration of substances and removal of blood, including routes and volumes. J. Appl. Toxicol. 21, 15–23 (2001).
Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G. & Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 8, 1263–1268 (2005).
Zimmerman, C. A. et al. Thirst neurons anticipate the homeostatic consequences of eating and drinking. Nature 537, 680–684 (2016).
Kitamura, K., Judkewitz, B., Kano, M., Denk, W. & Häusser, M. Targeted patch-clamp recordings and single-cell electroporation of unlabeled neurons in vivo. Nat. Methods 5, 61–67 (2008).
Chen, X., Leischner, U., Rochefort, N. L., Nelken, I. & Konnerth, A. Functional mapping of single spines in cortical neurons in vivo. Nature 475, 501–505 (2011).
Ballaro, A., Mundy, A. R., Fry, C. H. & Craggs, M. D. Bladder electrical activity: the elusive electromyogram. BJU Int. 92, 78–84 (2003).
Scheepe, J. R. et al. Smooth muscle electromyography of the urinary bladder. Neurourol. Urodyn. 17, 71–80 discussion 80–83. (1998).
Li, S. T. et al. Effects of orexins on myoelectric activity of sphincter of Oddi in fasted rabbits. Acta Pharmacol. Sin. 27, 212–216 (2006).
Gevaert, T. et al. Deletion of the transient receptor potential cation channel TRPV4 impairs murine bladder voiding. J. Clin. Invest. 117, 3453–3462 (2007).
Mann-Gow, T. K. et al. Evaluating the procedure for performing awake cystometry in a mouse model. J. Vis. Exp. 117, 123 (2017).
Qin, H. et al. A visual-cue-dependent memory circuit for place navigation. Neuron 99, 47–55.e4 (2018).
Acknowledgements
We thank J. Lou for assisting in the composition of the figures and the members of the Neuroscience Pioneer Club for valuable discussions. This work was supported by grants from the 1000 Talents Program for Young Scholars, the National Natural Science Foundation of China (No. 81671106, 81771175, 31700933), the National Basic Research Program of China (“973 Program”: 2015CB759500), the National Key Research and Development Program of China (2018YFA0109600), the Science Fund for Creative Research Group of China (61721092), and ULabor R&D Program (2017ULab0001).
Author information
Authors and Affiliations
Contributions
X.C., J. Yan, and X.X. designed the experiments. J. Yao, Q.Z., and Xianping Li conducted PRV tracing experiments, fiber photometry recordings, optogenetics/loss-of-function experiments, bladder recordings, and cystometry. J. Yao, X. Liao, and S.L. analyzed the data. Y.Z., Xiangning Li, and H.G. analyzed the 3D reconstruction data. J. Yao and Q.L. performed muscimol injections. M.W., J.L., and W.Z. conducted the electrophysiological experiments. J. Yao, Q.Z., J.Z., and T.L. performed viral injections. H.W. and W.H. performed post hoc histology. X. Liao, H.Q., and H.J. developed methods for the analysis of fiber photometry and behavioral data. F.X. and H.G. contributed to the whole-brain and spinal cord morphological experiments. X.C., X.X., and J. Yan wrote the manuscript with the help of all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
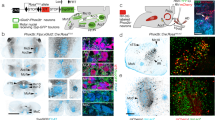
Supplementary Figure 1 EGFP+ regions in the brain and the spinal cord after PRV injection into the bladder wall.
a, Reconstructed images (top) and diagrams (bottom) of PRV-EGFP-labeled regions in the mouse brain on the sagittal (left) or coronal plane (right). The PRV-infected brain areas mainly include cerebral cortex (cortex), medial preoptic area (MPA), paraventricular nucleus (PVN), lateral hypothalamic area (LH), red nucleus (Red N), periaqueductal grey (PAG), locus coeruleus (LC), pontine micturition center (PMC), gigantocellular reticular nucleus (Gi), A5 noradrenaline cells (A5) and raphe nuclei. d: dorsal; v- ventral; r: rostral; c: caudal; l: left; ri: right. b, Left, schematic representation of the descending neural pathways infected by PRV. Right, confocal images showing the sections corresponding to the left panel. c-f, Coronal sections showing the lateral hypothalamic area (LH), PAG, red nucleus (Red N), A5 noradrenaline cells (A5), the gigantocellular reticular nucleus (Gi) and the raphe nuclei. All of these experiments were repeated independently in 3 male mice with similar results.
Supplementary Figure 2 Time-dependent infection of different brain areas after PRV-EGFP injection into the bladder wall.
Representative images showing PRV-infected neurons (green) in different brain areas at different time points after the viral injection. Left: 3 days; middle: 4 days; right: 4.5–5 days. These experiments were performed in 3 male mice for each group.
Supplementary Figure 3 PRV-infected neurons in different areas after separate PRV injections into left and right bladder walls or into the abdominal muscle.
Left and middle panels, representative images showing PRV-infected neurons (green and red) in different brain areas from one mouse with by PRV-EGFP injected into the right bladder wall and PRV-RFP into the left bladder wall. This experiment was repeated independently in 3 male mice with similar results. Right panel, no infected neurons were detected in these brain regions after PRV injection into the abdominal muscle. This experiment was repeated independently in 3 male mice with similar results.
Supplementary Figure 4 PRV-infected brain areas in female mice after PRV injection into the bladder wall.
a, Representative images showing PRV-infected neurons (green) in different brain areas in a female mouse. b, Serial sections showing PRV-infected neurons in the cerebral cortex of a female mouse. The same experiments were performed in 3 female mice.
Supplementary Figure 5 Histological confirmation of local viral injection into the L5 of M1 for optical-fiber-based Ca2+ signal recordings.
Top and Middle, serial sections showing GCaMP6f-labeled areas around the injection site in M1. Bottom, serial brain sections showing overlay of viral expression areas (in grey) from 5 male mice.
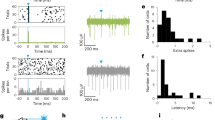
Supplementary Figure 6 GCaMP6 experiments confirming the correlation between neuronal activities of M1 L5 neurons and urination.
a, Schematics of fiber photometry recording of M1 L5 neurons infected with AAV-Syn-GCaMP6f. b, Post-hoc histological image showing the GCaMP6f labeled L5 neurons and fiber implantation. This experiment was repeated independently in 3 male mice with similar results. c, Left, an example of Ca2+ transients recorded in a freely moving mouse. Dashed line indicates the voiding onset. Right, averaged Ca2+ signals aligned to the onset of urination (red) or to the shuffled voiding onset (black). n = 14 trials from 3 male mice with GCaMP6f labeling. d, Quantification of amplitudes of urination-related Ca2+ transients (red) compared to the shuffled Ca2+ signals (black). Two-sided Wilcoxon signed-rank test, ***P = 0.00098 (z = 3.296, df = 13). n = 14 trials from 3 male mice. e, Representative traces showing blocking of the observed Ca2+ signals after muscimol injection into M1. Note that in the presence of muscimol, no Ca2+ signal was observed while the animal was moving. f, Comparison of the amplitudes of Ca2+ signals during and after muscimol application. One-sided Wilcoxon signed-rank test, ***P = 1.0 × 10–11 (z = −6.703; df = 59), n = 60 trials from 3 male mice with GCaMP6f labeling (20 trials from each mouse). g, An example of Ca2+ signals recorded in GCaMP6f-expressing M1 L5 neurons (black) during locomotion (purple) and urination. Light yellow box highlights the process of urination. Urine deposition area plotted in the bottom in red. h, Magnified view from panel g. Red arrows represent the urination onset, a time point during urination, and the urination offset. i, Screenshots showing the process of urine deposition at the three corresponding time points in panel h. All bar graphs and error bars represent mean ± s.e.m.
Supplementary Figure 7 Neural activities in M1 L5 neurons increase during territorial marking.
a, Heatmap of the Ca2+ signals was merged with spatial distribution of urine spots (ovals) and with positions when the mouse was still (triangles). The mouse was tested in a home cage lined with filter paper over 5 days (2 h for each day). The mouse was not given saline intraperitoneal (i.p.) injection before testing. b, Comparison of the amplitudes of Ca2+ signals during urinationand when the animals were still. One-sided Wilcoxon rank-sum test, ***P = 4.5 × 10–6 (z = 4.438; df = 28); n = 15 trials from 3 male mice. These experiments were performed with GCaMP6f labeling. All bar graphs and error bars represent mean ± s.e.m.
Supplementary Figure 8 Urination-associated Ca2+ transients in M1 L5 neurons of female mice.
a, An example of Ca2+ transients recorded in the M1 L5 of a female mouse. The red arrow indicates the urination onset. b, Overlay of all Ca2+ transients aligned to the onset of urination. n = 18 trials from 7 female mice. c, Averaged Ca2+ signal in M1 aligned to the urination onset (red) or to the shuffled urination onset (black). d, Quantification of amplitudes of urination-related Ca2+ transients (red) compared to the shuffled Ca2+ signals (black). Two-sided Wilcoxon signed-rank test, ***P = 0.0002 (z = 3.724, df = 17). n = 18 trials from 7 female mice. All bar graphs and error bars represent mean ± s.e.m.. e, Quantification of fraction of Ca2+ transients that correlated with urination compared to shuffled signals. Two-sided paired t-test, ***P = 0.00024 (t = 7.778; df = 6). n = 7 female mice. Box-and-whisker plot, center line (black bar), median; box, 25%–75% interquartile range; whiskers (grey bars), minimum and maximum. f, Onsets of M1 Ca2+ transients plotted relative to the onset of urination(dashed line). These experiments were performed with OGB-1AM labeling.
Supplementary Figure 9 The M1–PMC anatomical connection, as revealed by AAV- and VSV-based tracing.
a, Schematics of AAV-CaMKII-EGFP injection into M1 L5. b, Left and middle, images showing coronal slices of EGFP expression in L5 of the M1 (left) and EGFP-labeled axons in the PMC (middle). Right, An enlarged view around the area highlighted by the dashed circle from the middle panel. n = 5 male mice. c, Schematics of VSV-mCherry injection into M1. d, 6–7 days after VSV-mCherry injection into M1, anterograde-labeled neurons were observed in LC and PMC (top, middle). VSV-infected neurons were also detected in L1-L2 (top, right) and S6-S1 (bottom). The enlarged views of SPN are shown (bottom, middle and right). SPN, sacral parasympathetic neurons. L, left. R, right. 4 V, fourth ventricle. Tyrosine hydroxylase (TH) staining was used to label LC neurons. n = 3 male mice.
Supplementary Figure 10 Local injection of muscimol into M1 impairs urination; and histological confirmation of ablation of M1 neurons in the area corresponding to the region identified by PRV.
a, Schematics of the experimental protocol. b, Left, schematics showing the injection sites. Right, post-hoc histology showing the injection site. c, Quantification of the effects of muscimol or ACSF injection into different brain regions on urination(muscimol into M1: 12 male mice; ACSF into M1: 10 male mice; muscimol into Ma: 10 male mice; muscimol into the PMC: 10 male mice; ACSF into the PMC: 10 male mice; one-sided Wilcoxon signed-rank test for muscimol group compared with washout group or one-sided Wilcoxon rank-sum test for the other comparisons (***P = 0.000034, 0.000031, 0.000073, **P = 0.0012, 0.0028; z = −3.039, −3.986, −4.004, −2.772, −3.796; df = 11, 20, 20, 9, 18 from left to right). d, Quantification of the effects of muscimol or ACSF injection into the M1 of female mice on urination (muscimol into M1: 7 female mice; ACSF into M1: 7 female mice). One-sided Wilcoxon rank-sum test, ***P = 0.0008 (z = −3.154; df = 12). e, Serial coronal sections showing the NeuN staining in red and DAPI in blue in the M1 area 4 weeks after co-injection of AAV-CaMKII-Cre-GFP and AAV-DIO-taCasp3. This experiment was repeated independently in 11 male mice with similar results. All bar graphs and error bars represent mean ± s.e.m.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–10
Supplementary Video 1
Simultaneous recordings of Ca2+ signals in M1, locomotion and urination behavior. This movie corresponds to Supplementary Fig. 6
Supplementary Video 2
Simultaneous photostimulation of M1 L5 neurons and measurement of urination behavior
Rights and permissions
About this article

Cite this article
Yao, J., Zhang, Q., Liao, X. et al. A corticopontine circuit for initiation of urination. Nat Neurosci 21, 1541–1550 (2018). https://doi.org/10.1038/s41593-018-0256-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-018-0256-4
This article is cited by
-
Optogenetic stimulation of neurons in the anterior cingulate cortex induces changes in intravesical bladder pressure and the micturition reflex
Scientific Reports (2024)
-
BNST GABAergic neurons modulate wakefulness over sleep and anesthesia
Communications Biology (2024)
-
Electroacupuncture improves swallowing function in a post-stroke dysphagia mouse model by activating the motor cortex inputs to the nucleus tractus solitarii through the parabrachial nuclei
Nature Communications (2023)
-
A Novel Mouse Model for Polysynaptic Retrograde Tracing and Rabies Pathological Research
Cellular and Molecular Neurobiology (2023)
-
Neural circuit control of innate behaviors
Science China Life Sciences (2022)
