
Abstract
Macrophages promote both injury and repair after myocardial infarction, but discriminating functions within mixed populations remains challenging. Here we used fate mapping, parabiosis and single-cell transcriptomics to demonstrate that at steady state, TIMD4+LYVE1+MHC-IIloCCR2− resident cardiac macrophages self-renew with negligible blood monocyte input. Monocytes partially replaced resident TIMD4–LYVE1–MHC-IIhiCCR2− macrophages and fully replaced TIMD4−LYVE1−MHC-IIhiCCR2+ macrophages, revealing a hierarchy of monocyte contribution to functionally distinct macrophage subsets. Ischemic injury reduced TIMD4+ and TIMD4– resident macrophage abundance, whereas CCR2+ monocyte-derived macrophages adopted multiple cell fates within infarcted tissue, including those nearly indistinguishable from resident macrophages. Recruited macrophages did not express TIMD4, highlighting the ability of TIMD4 to track a subset of resident macrophages in the absence of fate mapping. Despite this similarity, inducible depletion of resident macrophages using a Cx3cr1-based system led to impaired cardiac function and promoted adverse remodeling primarily within the peri-infarct zone, revealing a nonredundant, cardioprotective role of resident cardiac macrophages.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


Data availability
Single-cell sequence data that support the findings of this study are available through the National Center for Biotechnology Information Gene Expression Omnibus (NCBI GEO) (GSE119355). The Agilent gene array data that support the findings of this study are available through the NCBI GEO (GSE119515).
Change history
12 March 2019
In the version of this article initially published, the equal contribution of the third author was omitted. The footnote links for that author should be “Sara Nejat1,11” and the correct statement is as follows: “11These authors contributed equally: Sarah A. Dick, Jillian A. Macklin, Sara Nejat.” The error has been corrected in the HTML and PDF versions of the article.
References
Lozano, R. et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380, 2095–2128 (2012).
Ismahil, M. A. et al. Remodeling of the mononuclear phagocyte network underlies chronic inflammation and disease progression in heart failure: critical importance of the cardiosplenic axis. Circ. Res. 114, 266–282 (2014).
Epelman, S., Lavine, K. J. & Randolph, G. J. Origin and functions of tissue macrophages. Immunity 41, 21–35 (2014).
Schulz, C. et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 336, 86–90 (2012).
Epelman, S. et al. Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation. Immunity 40, 91–104 (2014).
Ginhoux, F. & Guilliams, M. Tissue-resident macrophage ontogeny and homeostasis. Immunity 44, 439–449 (2016).
Porrello, E. R. et al. Transient regenerative potential of the neonatal mouse heart. Science 331, 1078–1080 (2011).
Lavine, K. et al. Distinct macrophage lineages contribute to disparate patterns of cardiac recovery and remodeling in the neonatal and adult heart. Proc. Natl Acad. Sci. USA 111, 16029–16034 (2014).
Aurora, A. B. et al. Macrophages are required for neonatal heart regeneration. J. Clin. Invest. 124, 1382–1392 (2014).
Leid, J. M. et al. Primitive embryonic macrophages are required for coronary development and maturation. Circ. Res. 118, 1498–1511 (2016).
Panizzi, P. et al. Impaired infarct healing in atherosclerotic mice with Ly-6Chi monocytosis. J. Am. Coll. Cardiol. 55, 1629–1638 (2010).
van Amerongen, M. J., Harmsen, M. C., van Rooijen, N., Petersen, A. H. & van Luyn, M. J. Macrophage depletion impairs wound healing and increases left ventricular remodeling after myocardial injury in mice. Am. J. Pathol. 170, 818–829 (2007).
Molawi, K. et al. Progressive replacement of embryo-derived cardiac macrophages with age. J. Exp. Med. 211, 2151–2158 (2014).
Serbina, N. V. & Pamer, E. G. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat. Immunol. 7, 311–317 (2006).
Ridker, P. M. et al. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N. Engl. J. Med. 377, 1119–1131 (2017).
Hulsmans, M. et al. Macrophages facilitate electrical conduction in the heart. Cell 169, 510–522 e520 (2017).
Miyanishi, M. et al. Identification of Tim4 as a phosphatidylserine receptor. Nature 450, 435–439 (2007).
Clemente-Casares, X. et al. A CD103+ conventional dendritic cell surveillance system prevents development of overt heart failure during subclinical viral myocarditis. Immunity 47, 974–989 (2017).
Van der Borght, K. et al. Myocardial infarction primes autoreactive T cells through activation of dendritic cells. Cell Rep. 18, 3005–3017 (2017).
Gautier, E. L. et al. Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat. Immunol. 13, 1118–1128 (2012).
Tirosh, I. et al. Dissecting the multicellular ecosystem of metastatic melanoma by single-cell RNA-seq. Science 352, 189–196 (2016).
Guilliams, M. et al. Unsupervised high-dimensional analysis aligns dendritic cells across tissues and species. Immunity 45, 669–684 (2016).
Satpathy, A. T. et al. Zbtb46 expression distinguishes classical dendritic cells and their committed progenitors from other immune lineages. J. Exp. Med. 209, 1135–1152 (2012).
Trapnell, C. et al. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 32, 381–386 (2014).
Chen, J., Schlitzer, A., Chakarov, S., Ginhoux, F. & Poidinger, M. Mpath maps multi-branching single-cell trajectories revealing progenitor cell progression during development. Nat. Commun. 7, 11988 (2016).
Parkhurst, C. N. et al. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell 155, 1596–1609 (2013).
Yona, S. et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity 38, 79–91 (2013).
Leuschner, F. et al. Rapid monocyte kinetics in acute myocardial infarction are sustained by extramedullary monocytopoiesis. J. Exp. Med. 209, 123–137 (2012).
Leuschner, F. et al. Angiotensin-converting enzyme inhibition prevents the release of monocytes from their splenic reservoir in mice with myocardial infarction. Circ. Res. 107, 1364–1373 (2010).
Nahrendorf, M. et al. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J. Exp. Med. 204, 3037–3047 (2007).
Wan, E. et al. Enhanced efferocytosis of apoptotic cardiomyocytes through myeloid-epithelial-reproductive tyrosine kinase links acute inflammation resolution to cardiac repair after infarction. Circ. Res. 113, 1004–1012 (2013).
Lim, H. Y. et al. Hyaluronan receptor LYVE-1-expressing macrophages maintain arterial tone through hyaluronan-mediated regulation of smooth muscle cell collagen. Immunity 49, 326–341 e327 (2018).
Majmudar, M. D. et al. Monocyte-directed RNAi targeting CCR2 improves infarct healing in atherosclerosis-prone mice. Circulation 127, 2038–2046 (2013).
Bajpai, G. et al. The human heart contains distinct macrophage subsets with divergent origins and functions. Nat. Med. 24, 1234–1245 (2018).
Satoh, T. et al. Identification of an atypical monocyte and committed progenitor involved in fibrosis. Nature 541, 96–101 (2017).
Menezes, S. et al. The heterogeneity of Ly6Chi monocytes controls their differentiation into iNOS+ macrophages or monocyte-derived dendritic cells. Immunity 45, 1205–1218 (2016).
Ikeda, N. et al. Emergence of immunoregulatory Ym1(+)Ly6C(hi) monocytes during recovery phase of tissue injury. Sci. Immunol. 3, eaat0207 (2018).
Mrdjen, D. et al. High-dimensional single-cell mapping of central nervous system immune cells reveals distinct myeloid subsets in health, aging, and disease. Immunity 48, 599 (2018).
Han, X. et al. Mapping the mouse cell atlas by microwell-seq. Cell 172, 1091–1107 e1017 (2018).
Mass, E. et al. Specification of tissue-resident macrophages during organogenesis. Science 353, aaf4238 (2016).
van de Laar, L. et al. Yolk sac macrophages, fetal liver, and adult monocytes can colonize an empty niche and develop into functional tissue-resident macrophages. Immunity 44, 755–768 (2016).
Gosselin, D. et al. An environment-dependent transcriptional network specifies human microglia identity. Science 356, eaal3222 (2017).
Aronoff, L., Epelman, S. & X, C.-C. Isolation and identification of extravascular immune cells of the heart. J. Vis. Exp. 138, 7 (2018).
Noyan-Ashraf, M. H. et al. GLP-1R agonist liraglutide activates cytoprotective pathways and improves outcomes after experimental myocardial infarction in mice. Diabetes 58, 975–983 (2009).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420 (2018).
Lun, A., McCarthy, D. & Marioni, J. A step-by-step workflow for low-level analysis of single-cell RNA-seq data with Bioconductor. F1000 Res. 5, 2122 (2016).
McCarthy, D., Campbell, K., Lun, A. & Wills, Q. Scater: pre-processing, quality control, normalization and visualization of single-cell RNA-seq data in R. Bioinformatics 33, 1179–1186 (2017).
Blondel, V. D., Guillaume, J. L., Lambiotte, R. & Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. 10, 10008 (2008).
Finak, G. et al. MAST: a flexible statistical framework for assessing transcriptional changes and characterizing heterogeneity in single-cell RNA sequencing data. Genome Biol. 16, 278 (2015).
Reimand, J. et al. g:Profiler—a web server for functional interpretation of gene lists (2016update). Nucleic Acids Res. 44, W83–W89 (2016).
Qiu, X. et al. Reversed graph embedding resolves complex single-cell trajectories. Nat. Meth. 14, 979–982 (2017).
Acknowledgements
This work was supported by the Canadian Institutes of Health Research (S.E., PJT364831; J.A.M. and S.A.D.), Heart and Stroke Foundation (S.E.), March of Dimes (S.E.), Ted Rogers Centre for Heart Research (S.E., S.A.D. and J.A.M.), the Peter Munk Cardiac Centre (S.E.) and the National Institutes of Health (S.E. K08HL112826). We thank D. Mann for his insight and advice, and S. Wilson for editorial assistance. Thanks to N. Winegarden and N. Khuu for help with processing single-cell RNA sequencing samples.
Author information
Authors and Affiliations
Contributions
S.A.D. and J.A.M. designed and performed experiments with the help of X.C-C., S.H., C.K., M.G.A., A.W., L.A., R.Z., R.B. and I.B. A.M. performed all surgeries. M.H., K.J.L., B.R., F.G., M.I.C. and C.S.R. provided expertise and feedback. S.N. performed the bioinformatics analyses. J.C. performed the Mpath analysis. S.E. conceived the study, obtained funding and wrote the manuscript with S.A.D., S.N. and J.A.M.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 Gene-expression analysis of cardiac macrophage clusters defined by single-cell RNA sequencing reveals five distinct populations.
a) Cardiac mononuclear phagocytes were sorted from adult c57bl/6j mice (~20 weeks old). Total macrophages (MFs) (R2; CD45+CD64lo–hiCD11b+) and total dendritic cells (DCs) (R3; CD45+CD64–CD11c+MHC-IIint–hi). The two sorted populations were pooled in a 1:1 ratio before processing for scRNA-seq on the 10x Genomics Chromium platform. N=8 (4 male; 4 female); one experiment. b) The top ~65 differentially expressed genes in each cluster were input into the ImmGEN online portal for "My Geneset" http://rstats.immgen.org/MyGeneSet_New/index.html to generate a similarity score. We included all MF, monocyte and dendritic cell populations in the V1 array database for the assessment to generate a means-normalized expression scatter plot. Colored boxes highlight similarity in dendritic cell (light green), macrophage (dark green) and monocyte (red) populations to the sorted populations contributed to ImmGen. c) Violin plots of cluster defining genes in the 11 clusters defined in Fig. 1c. *p<0.01 (adjusted P value). Statistical analysis of scRNA-seq is defined in the material and methods, and was performed using the MAST method.
Supplementary Figure 2 Cx3cr1CreER:R26Td mice retain the ability to distinguish resident macrophages from recruited macrophages following irradiation and adoptive transplant.
a) Cx3cr1CreER–YFP:R26Td mice were used to determine Td reporter expression in cardiac macrophages (CD45+CD64+CD11b+), brain microglia (CD45lowCD64+CD11b+), liver Kupffer cells (CD45+CD64+CD11b+) and blood monocytes (CD115+CD11b+) in the absence of tamoxifen administration (No TAM) at the indicated times (from E17.5 to 77 days post-natal). Recombination in tissue macrophages without TAM was compared to pregnant mice that were given a TAM pulse at E18.5 in utero (via i.p. injection of 2mg TAM). N=3; center value, mean; error bars, SEM; one representative experiment, repeated twice. b) A second fate-mapping system was also used (Cx3cr1CreERT2)30 and bred to the same reporter R26Td line to assess for Td expression in cardiac macrophages (heart) and microglia (brain) at the indicated times either without TAM (No TAM) or following TAM administration at E18.5 (+TAM). N=3; center value, mean; one experiment. c) Bone marrow from adult Cx3cr1CreER–YFP:R26Td mice (Donor BM, CD45.2), not previously pulsed with tamoxifen, was isolated and the %Td+ macrophages was determined in blood CD115+CD11b+ monocytes and total CD45+ bone marrow. Donor Bone marrow was transplanted into non-lethal irradiated CD45.1 mice (Recipients). After 6 weeks, chimerism rates in the recipient were ~95%. Blood and cardiac tissue was isolated from the CD45.1 recipient. A subset of recipient mice was given a pulse of tamoxifen (TAM) (i.p. 2mg) 2 days prior to isolation to confirm the responsiveness of chimeric CD45.2 cells in the recipient compared to without TAM administration (No TAM). The %Td+ cells was determined in total CD45.2+ blood monocytes (CD115+CD11b+) and total CD45.2+ cardiac macrophages (MFs) (CD64+CD11b+). N=3,2; center value, mean; error bars, SEM; one experiment.
Supplementary Figure 3 Tracking resident cardiac macrophages post myocardial infarction.
a-f) 3-week-old Cx3cr1CreER–YFP:R26Td mice were fed tamoxifen (TAM)- containing chow for 10 days, and then mice were either left as non-infarcted controls for an additional 6 weeks before a myocardial infarction was performed (MI). a) Ischemic tissue was isolated at Day 2 and Day 28 post-MI and total resident cardiac macrophages (CD45+CD64+CD11b+Td+) were analyzed as TIMD4+ and TIMD4– subsets per mg/tissue compared to non-infarcted controls (No MI). N=3; repeated independently 2 times with similar results. b) Total CD45+CD64+CD11b+ cardiac macrophages (MFs) were partitioned into TIMD4+, CCR2+ and TIMD4–CCR2– MF subsets. LYVE1 was detected by flow cytometry. N=3; repeated independently 2 times with similar results. c) Representative images of hearts isolated, fixed and sectioned longitudinally for immunofluorescence from Cx3cr1CreER–YFP:R26Td mice post-MI. Td expression was induced as in panel (a) above. Cardiac macrophages (CD68+, white) were counted based on their coexpression for the residency marker Td (red) and LYVE1 (green) in the infarct and peri-infarct regions, defined in Fig. 4a, from 3 individual images/region/mouse. N=3; one experiment. d) Hearts were isolated from the ischemic zone of Cx3cr1CreER–YFP:R26Td/DTR mice day 4 post-MI after a 2 hr-BrdU pulse. Total Td+ cardiac macrophages were subdivided into TIMD4+ and TIMD4– MF populations and %BrdU+ macrophages was quantified. N=3,3,4,4; one experiment. e) Heat map of the 20 most differentially expressed genes from clusters from Fig. 5b. Overlapping clusters (#1 to #6a), represent macrophages from control and post-MI samples that clustered together. For the overlapping clusters both the control and MI cells from each cluster are aligned next to each other in the heatmap. f) Means-normalized expression scatter plot produced by comparison of the differentially expressed genes from each unique cluster (7, 8, 9, 10 and 11) in Fig. 5b to the ImmGen data set. For all: *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001; two-tailed Student’s T test; center value, mean; error bars, SEM.
Supplementary Figure 4 Chimeric Cx3cr1CreER–YFP:R26Td monocytes retain the ability to distinguish resident macrophages from recruited macrophages post myocardial infarction in the setting of parabiosis and shielded irradiation.
a) Cx3cr1CreER–YFP:R26Td mice (Donor CD45.2) were fed tamoxifen (TAM)-containing chow at 3 weeks of age for 1 week, TAM was then discontinued for additional 10 weeks. At this time, Cx3cr1CreER–YFP:R26Td mice were paired with recipient CD45.1 mice. Two weeks after pairing, recipient CD45.1 mice were infarcted by surgical ligation of the left anterior descending artery (MI). At day 5 post-MI, blood and cardiac tissue (ischemic zone) was quantified for %Td+ in chimeric CD45.2 macrophages (MFs) (CD45+CD64+CD11b+) and blood monocytes (CD115+CD11b+) in the CD45.1 recipient mouse. N=3. One experiment. b) Bone marrow was isolated from adult Cx3cr1CreER–YFP:R26Td mice (Donor BM, CD45.2) treated as in panel (a). The bone marrow cells were transplanted into non-lethally irradiated CD45.1 mice (Recipient) that had a lead shield covering the heart and thoracic cavity to minimize loss of resident cardiac macrophages. Following 6 weeks of reconstitution, surgical MI was performed on the recipient CD45.1 mouse and it recovered for an additional 4 weeks. Blood and cardiac tissue (separated into ischemic and remote zone, see Fig. 4a) was taken for flow cytometric analysis. A subset of mice was given a pulse of TAM (i.p.) 2 days prior to isolation to confirm the responsiveness of the CD45.2 cells to activate the TAM-inducible Td reporter compared to No TAM controls. Gated on total CD45.2+ blood monocytes (CD115+CD11b+) and total CD45.2+ cardiac macrophages (CD64+CD11b+) in the CD45.1 recipient and %Td+ was quantified. N=3; one experiment. For all: center value, mean; error bars, SEM.
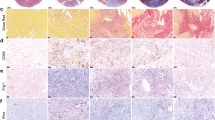
Supplementary Figure 5 Similar cardiac macrophage subsets exist in human cardiomyopathy.
Left ventricular cardiac tissue was obtained from human patients with end-stage cardiomyopathy during implantation of left ventricular assist devices. a) Tissue was digested and analyzed by flow cytometry [gated similar to mouse (CD45+CD64+MerTK+CD14+)] or b) sorted for Hema-3 staining analysis and visualization. c) Cardiac macrophage (MF) populations were sorted from human tissue based on MHC-II and CCR2 expression. RNA was extracted, amplified, and gene expression was determined by gene array (Agilent). Genes that were most differentially expressed in the 5 Seurat MF clusters identified by scRNA-seq in the mouse (Fig. 1d) were compared to the gene expression across the three human MF subsets isolated to generate heatmaps. N=6 patient samples; one experiment.
Supplementary Figure 6 Resident cardiac macrophages proliferate post myocardial infarction in the remote myocardium, with resident macrophage depletion leading to increased cardiomyocyte hypertrophy without cardiac fibrosis.
3-week-old Cx3cr1CreER–YFP:R26Td mice were fed tamoxifen (TAM)- containing chow for 10 days, and then mice were either left as non-infarcted controls for an additional 6 weeks. a) Diphtheria toxin (DT) was then administered to Cx3cr1CreER–YFP:R26Td/DTR mice for 2 weeks at steady state. Blood monocytes (CD115+CD11b+Ly6C+), neutrophils (CD45+CD11b+CD64–Ly6G+), lymphocytes (CD45+CD11b+CD64–), Td–CCR2+ cardiac macrophages, and Td+ cardiac macrophages were quantified by flow cytometry, normalized to DT-injected Cx3cr1+/+:R26Td/DTR control mice. N=5; repeated independently 3 times with similar results. b) Representative flow plots of resident macrophage depletion during steady state (Fig. 6a) from blood and cardiac tissue following a myocardial infarction demonstrating effective depletion of resident macrophages. c) Naive wild-type mice were given chronic PBS or 250ng DT injections (i.p.) for 35 days. Whole hearts were either cut longitudinally and sectioned to be stained with Fast Green for interstitial fibrosis, quantified as % Fibrosis using ImageJ software, or quantified by flow cytometry for the number of neutrophils or cardiac macrophages normalized per mg tissue. N=3, 7; one experiment. d) Hearts were isolated from adult TAM-fed Cx3cr1CreER–YFP:R26Td/DTR mice (as above) day 5 post-MI, comparing non-depleted or depleted mice (DT treated daily). Recruited macrophages (MF), monocytes, and neutrophils were quantified and normalized per mg tissue in the ischemic and remote cardiac zones. N=6,5,4; repeated independently 2 times with similar results. e-f) Hearts were isolated from adult Cx3cr1CreER–YFP:R26Td mice following TAM-induction (as above), and mice received BrdU injection 2 hours prior to harvest. Resident MFs (Td+), recruited MFs (Td–) and monocytes were normalized per mg tissue in the remote zone at various time points (e), or %BrdU+ was determined (f), compared to an uninfarcted control (No MI). N(Td+)=13, 13, 6, 3, 10. N(Td–, mono)= 12, 10, 7, 3, 6; Repeated 3 times for day 2, 2 times for day 4 and 28, 1 time for day 7, 6 times for control with similar results. g) Change in fractional shortening (contractile function) from day 7 to day 28 post-MI in the remote zone comparing mice with or without chronic resident macrophage depletion (Paired T-test) through the use of M-mode echocardiography. N=11; repeated 2 times independently with similar results. h-i) Longitudinally cut hearts from Cx3cr1CreER–YFP:R26Td/DTR mice (as above) isolated day 35 post-MI to compare post-MI control and depleted groups. Cardiomyocyte hypertrophy was detected using Wheat Germ Agglutinin staining in the remote zone. Hearts imaged at 20x magnification and cardiomyocytes measured in ImageJ, 8 images/zone/heart at 2 cutting levels. N=4, 6, 6; repeated once with similar results (h); Cardiac tissue fibrosis was detected using Fast Green staining. Percentage fibrotic area was quantified with ImageJ analysis, 4 images/zone/heart at 2 cutting levels. N = 9; repeated once with similar results. For all: *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001; Two-tailed Student’s T test; center value, mean; error bars, SEM.
Supplementary information
Supplementary Figures 1-6
Supplementary Figures 1-6 Supplementary Table 1
Supplementary Table 2
DEgenes For Supplementary Figure 3e
Rights and permissions
About this article

Cite this article
Dick, S.A., Macklin, J.A., Nejat, S. et al. Self-renewing resident cardiac macrophages limit adverse remodeling following myocardial infarction. Nat Immunol 20, 29–39 (2019). https://doi.org/10.1038/s41590-018-0272-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-018-0272-2
This article is cited by
-
The roles of tissue resident macrophages in health and cancer
Experimental Hematology & Oncology (2024)
-
The PD-1–PD-L1 pathway maintains an immunosuppressive environment essential for neonatal heart regeneration
Nature Cardiovascular Research (2024)
-
ALKBH5-mediated m6A modification of IL-11 drives macrophage-to-myofibroblast transition and pathological cardiac fibrosis in mice
Nature Communications (2024)
-
Animal models to study cardiac regeneration
Nature Reviews Cardiology (2024)
-
ATF3’s shielding hand to counter ischemia-reperfusion injury in cardiac macrophages
Nature Cardiovascular Research (2024)
