
Abstract
Dendritic cells (DCs) play an integral role in regulating mucosal immunity and homeostasis, but the signaling network mediating this function of DCs is poorly defined. We identified the noncanonical NF-κB-inducing kinase (NIK) as a crucial mediator of mucosal DC function. DC-specific NIK deletion impaired intestinal immunoglobulin A (IgA) secretion and microbiota homeostasis, rendering mice sensitive to an intestinal pathogen, Citrobacter rodentium. DC-specific NIK was required for expression of the IgA transporter polymeric immunoglobulin receptor (pIgR) in intestinal epithelial cells, which in turn relied on the cytokine IL-17 produced by TH17 cells and innate lymphoid cells (ILCs). NIK-activated noncanonical NF-κB induced expression of IL-23 in DCs, contributing to the maintenance of TH17 cells and type 3 ILCs. Consistent with the dual functions of IL-23 and IL-17 in mucosal immunity and inflammation, NIK deficiency also ameliorated colitis induction. Thus, our data suggest a pivotal role for the NIK signaling axis in regulating DC functions in intestinal immunity and homeostasis.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Gutzeit, C., Magri, G. & Cerutti, A. Intestinal IgA production and its role in host–microbe interaction. Immunol. Rev. 260, 76–85 (2014).
Brandtzaeg, P. Secretory IgA: designed for anti-microbial defense. Front. Immunol. 4, 222 (2013).
Palm, N. W. et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 158, 1000–1010 (2014).
Coombes, J. L. & Powrie, F. Dendritic cells in intestinal immune regulation. Nat. Rev. Immunol. 8, 435–446 (2008).
Briseno, C. G., Murphy, T. L. & Murphy, K. M. Complementary diversification of dendritic cells and innate lymphoid cells. Curr. Opin. Immunol. 29, 69–78 (2014).
Walker, J. A., Barlow, J. L. & McKenzie, A. N. Innate lymphoid cells—How did we miss them? Nat. Rev. Immunol. 13, 75–87 (2013).
Buela, K. A., Omenetti, S. & Pizarro, T. T. Cross-talk between type 3 innate lymphoid cells and the gut microbiota in inflammatory bowel disease. Curr. Opin. Gastroenterol. 31, 449–455 (2015).
Cerutti, A. The regulation of IgA class switching. Nat. Rev. Immunol. 8, 421–434 (2008).
Massacand, J. C. et al. Intestinal bacteria condition dendritic cells to promote IgA production. PLoS One 3, e2588 (2008).
Ruane, D. et al. Microbiota regulate the ability of lung dendritic cells to induce IgA class-switch recombination and generate protective gastrointestinal immune responses. J. Exp. Med. 213, 53–73 (2016).
Oh, H. & Ghosh, S. NF-κB: roles and regulation in different CD4+ T-cell subsets. Immunol. Rev. 252, 41–51 (2013).
Sun, S. C. The non-canonical NF-κB pathway in immunity and inflammation. Nat. Rev. Immunol. 17, 545–558 (2017).
Sun, S. C. The noncanonical NF-κB pathway. Immunol. Rev. 246, 125–140 (2012).
Senftleben, U. et al. Activation of IKKa of a second, evolutionary conserved, NF-kB signaling pathway. Science 293, 1495–1499 (2001).
Xiao, G., Harhaj, E. W. & Sun, S. C. NF-κB-inducing kinase regulates the processing of NF-κB2 p100. Mol. Cell 7, 401–409 (2001).
Liao, G., Zhang, M., Harhaj, E. W. & Sun, S. C. Regulation of the NF-κB-inducing kinase by tumor necrosis factor receptor-associated factor 3-induced degradation. J. Biol. Chem. 279, 26243–26250 (2004).
Katakam, A. K. et al. Dendritic cells require NIK for CD40-dependent cross-priming of CD8+ T cells. Proc. Natl Acad. Sci. USA 112, 14664–14669 (2015).
Vedantam, G. & Viswanathan, V. K. Unlocking the gates to inflammatory bowel disease: the role of Enterococcus faecalis gelatinase. Gastroenterology 141, 795–798 (2011).
Zhou, Y. et al. Increased Enterococcus faecalis infection is associated with clinically active Crohn disease. Medicine (Baltimore) 95, e5019 (2016).
Jett, B. D., Huycke, M. M. & Gilmore, M. S. Virulence of enterococci. Clin. Microbiol. Rev. 7, 462–478 (1994).
Thompson, C. L., Vier, R., Mikaelyan, A., Wienemann, T. & Brune, A. ‘Candidatus Arthromitus’ revised: segmented filamentous bacteria in arthropod guts are members of Lachnospiraceae. Environ. Microbiol. 14, 1454–1465 (2012).
Ericsson, A. C., Hagan, C. E., Davis, D. J. & Franklin, C. L. Segmented filamentous bacteria: commensal microbes with potential effects on research. Comp. Med. 64, 90–98 (2014).
Talham, G. L., Jiang, H. Q., Bos, N. A. & Cebra, J. J. Segmented filamentous bacteria are potent stimuli of a physiologically normal state of the murine gut mucosal immune system. Infect. Immun. 67, 1992–2000 (1999).
Jaffar, Z., Ferrini, M. E., Herritt, L. A. & Roberts, K. Cutting edge: lung mucosal Th17-mediated responses induce polymeric Ig receptor expression by the airway epithelium and elevate secretory IgA levels. J. Immunol. 182, 4507–4511 (2009).
Cao, A. T., Yao, S., Gong, B., Elson, C. O. & Cong, Y. Th17 cells upregulate polymeric Ig receptor and intestinal IgA and contribute to intestinal homeostasis. J. Immunol. 189, 4666–4673 (2012).
Zheng, Y. et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nat. Med. 14, 282–289 (2008).
Pham, T. A. et al. Epithelial IL-22RA1-mediated fucosylation promotes intestinal colonization resistance to an opportunistic pathogen. Cell. Host. Microbe. 16, 504–516 (2014).
Eberl, G. RORgammat, a multitask nuclear receptor at mucosal surfaces. Mucosal Immunol. 10, 27–34 (2017).
Hue, S. et al. Interleukin-23 drives innate and T cell-mediated intestinal inflammation. J. Exp. Med. 203, 2473–2483 (2006).
Kullberg, M. C. et al. IL-23 plays a key role in Helicobacter hepaticus-induced T cell-dependent colitis. J. Exp. Med. 203, 2485–2494 (2006).
Yen, D. et al. IL-23 is essential for T cell-mediated colitis and promotes inflammation via IL-17 and IL-6. J. Clinic. Invest. 116, 1310–1316 (2006).
Maloy, K. J. & Kullberg, M. C. IL-23 and Th17 cytokines in intestinal homeostasis. Mucosal Immunol. 1, 339–349 (2008).
Shih, V. F. et al. Control of RelB during dendritic cell activation integrates canonical and noncanonical NF-κB pathways. Nat. Immunol. 13, 1162–1170 (2012).
Yu, J. et al. T cell–intrinsic function of the noncanonical NF-κB pathway in the regulation of GM-CSF expression and experimental autoimmune encephalomyelitis pathogenesis. J. Immunol. 193, 422–430 (2014).
Liang, C., Zhang, M. & Sun, S. C. β-TrCP binding and processing of NF-κB2/p100 involve its phosphorylation at serines 866 and 870. Cell. Signal. 18, 1309–1317 (2006).
Tucker, E. et al. A novel mutation in the Nfkb2 gene generates an NF-κB2 ‘super repressor’. J. Immunol. 179, 7514–7522 (2007).
Li, X., Yang, Y. & Ashwell, J. D. TNF-RII and c-IAP1 mediate ubiquitination and degradation of TRAF2. Nature 416, 345–347 (2002).
Mangan, P. R. et al. Transforming growth factor-beta induces development of the TH17 lineage. Nature 441, 231–234 (2006).
Basu, R. et al. Th22 cells are an important source of IL-22 for host protection against enteropathogenic bacteria. Immunity 37, 1061–1075 (2012).
Catana, C. S. et al. Contribution of the IL-17/IL-23 axis to the pathogenesis of inflammatory bowel disease. World J. Gastroenterol. 21, 5823–5830 (2015).
Neurath, M. F. Current and emerging therapeutic targets for IBD. Nat. Rev. Gastroenterol. Hepatol. 14, 269–278 (2017).
Kühn, R., Löhler, J., Rennick, D., Rajewsky, K. & Müller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 75, 263–274 (1993).
Specht, S., Arriens, S. & Hoerauf, A. Induction of chronic colitis in IL-10 deficient mice requires IL-4. Microbes Infect. 8, 694–703 (2006).
Davidson, N. J., Fort, M. M., Muller, W., Leach, M. W. & Rennick, D. M. Chronic colitis in IL-10−/− mice: insufficient counter regulation of a Th1 response. Int. Rev. Immunol. 19, 91–121 (2000).
Gomes-Santos, A. C. et al. New insights into the immunological changes in IL-10-deficient mice during the course of spontaneous inflammation in the gut mucosa. Clin. Dev. Immunol. 2012, 560817 (2012).
Suzuki, K. et al. Aberrant expansion of segmented filamentous bacteria in IgA-deficient gut. Proc. Natl Acad. Sci. USA 101, 1981–1986 (2004).
Tumanov, A. V. et al. Lymphotoxin controls the IL-22 protection pathway in gut innate lymphoid cells during mucosal pathogen challenge. Cell. Host. Microbe. 10, 44–53 (2011).
Brightbill, H. D. et al. Conditional deletion of NF-κB-inducing kinase (NIK) in adult mice disrupts mature B cell survival and activation. J. Immunol. 195, 953–964 (2015).
Jin, J. et al. Epigenetic regulation of the expression of Il12 and Il23 and autoimmune inflammation by the deubiquitinase Trabid. Nat. Immunol. 17, 259–268 (2016).
Ajami, N. J., Cope, J. L., Wong, M. C., Petrosino, J. F. & Chesnel, L. Impact of oral fidaxomicin administration on the intestinal microbiota and susceptibility to Clostridium difficile colonization in mice. Antimicrob. Agents Chemother. 62, e02112-17 (2018).
Caporaso, J. G. et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 6, 1621–1624 (2012).
Hu, H. et al. OTUD7B controls non-canonical NF-κB activation through deubiquitination of TRAF3. Nature 494, 371–374 (2013).
Qiu, J. et al. The aryl hydrocarbon receptor regulates gut immunity through modulation of innate lymphoid cells. Immunity 36, 92–104 (2012).
Nelson, J. D., Denisenko, O. & Bomsztyk, K. Protocol for the fast chromatin immunoprecipitation (ChIP) method. Nat. Protoc. 1, 179–185 (2006).
Acknowledgements
We thank Genentech, Inc. for providing the Map3k14-flox mice, Walter and Eliza Hall Institute of Medical Research for Nfkb2Lym1 mice, and National Cancer Institute Preclinical Repository for NF-κB p100/p52 antibody. This work was supported by grants from the National Institutes of Health (GM84459, AI057555, AI104519, and AI64639). This study also used the NIH/NCI-supported resources under award number P30CA016672 at The MD Anderson Cancer Center.
Author information
Authors and Affiliations
Contributions
Z.J. and J.-Y.Y. designed and performed the research, prepared the experiments, and wrote part of the manuscript; M.G., H.W., X.X., Y.L., T.L., L.Zhu, J.S., L.Zhang, X.Z., D.J., D.L., and X.C. contributed experiments; H.D.B. contributed critical reagents; Y.C. made supervising contributions to intestinal immune cells analyses; and S.-C.S. supervised the work and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
H.D.B. is an employee of Genentech, Inc. The other authors declare no competing interests. Genentech, Inc. provided the Map3k14-flox mice.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
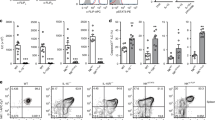
Supplementary Figure 1 DC-specific NIK deficiency does not affect the frequency or maturation state of DCs.
a, Genotyping PCR analysis of tail DNA from wild-type (WT; NIK+/+CD11c-Cre), heterozygous (Het; NIK+/flCD11c-Cre) and Map3k14-cKO (NIKfl/flCD11c-Cre) mice, showing the Map3k14 floxed and wild-type alleles. b, qRT–PCR analysis of Map3k14 mRNA encoded by the deleted exon in Map3k14-cKO (cKO) mice using sorted DCs and control T cells (n = 3). c, Immunoblot analysis of the indicated proteins in whole-cell lysates of wild-type and Map3k14-cKO BMDCs treated for 3 h with anti-CD40 in the presence ( + ) or absence (–) of the proteasome inhibitor MG132 (MG132 was used to block NIK degradation). d,e, Flow cytometric analysis of the CD11c+MHCII+ DC population (d) and surface expression of CD80 and CD86 in DCs (e) from spleen, mesenteric lymph nodes (MLN), Peyer’s patches (PP), and the other indicated organs. f,g, Flow cytometric analysis of total lamina propria (LP) DCs (CD11c+MHCII+) (f, n = 4 mice/group) or the indicated DC subsets, including CD103+ DCs (n = 4 mice/group), myeloid DCs (CD8a+CD11b–CD11c+) (n = 5 mice/group), lymphoid DCs (CD8a–CD11b+CD11c+) (n = 5 mice/group), TLR5+ DCs (n = 4 mice/group), and pDCs (n = 5 mice/group) (g). h, Flow cytometric analysis of the indicated surface markers in total LP DCs. Each symbol represents an individual mouse, and small horizontal lines indicate the mean ( ± s.e.m.) (b,f,g). Data were analyzed by unpaired two-tailed Student’s t test with P values shown above the plots (b). Data were representative of two independent experiments.
Supplementary Figure 2 16 S rRNA sequencing analysis of IgA+ and IgA– microbiota.
a,b, 16 S rRNA sequencing analysis of IgA+ and IgA– fecal bacteria, isolated from multiple mice (n = 6 per genotype) by magnetic-activated cell sorting (MACS), showing trends in beta diversity based on principal-coordinate analysis (PCoA; a) and taxa abundances (b). Each symbol represents an individual mouse (a,b), and b is presented as box plots (where the middle line indicates the mean value and all individual plots are shown). Data were analyzed by multiple comparisons using the Holm–Sidak method (b), with P values shown above the plots. Data are representative of three independent experiments.
Supplementary Figure 3 DC-specific NIK deficiency has no effect on serum IgA concentration, B cell frequency, or Peyer’s patch size and numbers.
a, ELISA of IgA concentration in the sera of wild-type (WT; n = 8) and Map3k14-cKO (cKO; n = 8) mice with the dilution factors indicated. b,c, Flow cytometric analysis of B cell frequency in the spleen, mesenteric lymph nodes (MLN) and Peyer’s patches (PP) of age-matched wild-type and Map3k14-cKO mice. Data are presented as a representative plot (b) and summary graph (c) based on multiple mice (each circle represents a mouse, n = 6–7 mice/group). d, Representative images of Peyer’s patches from age-matched wild-type and Map3k14-cKO mice. e–g, Summary graphs of the size (e, wild type, n = 68; cKO, n = 58), numbers (f, wild type, n = 27; cKO, n = 24) and cell numbers (g, n = 5 mice/group) of Peyer’s patches of age-matched wild-type and Map3k14-cKO mice. Each symbol represents an individual mouse (a,c,f,g) or an individual Peyer’s patch (e); small horizontal lines indicate the mean ( ± s.e.m.). Data were analyzed by unpaired two-tailed Student’s t test and are representative of three independent experiments.
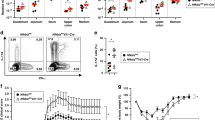
Supplementary Figure 4 Microbiota composition and fecal concentration of IgA and pIgR in co-housed wild-type and NIK-DKO mice.
a, 16 S rRNA sequencing analysis of the genus levels of microbiota composition from age-matched and co-housed wild-type (WT, n = 8) and Map3k14-cKO (n = 8) mice. b, Analysis of Enterococcus spp. in the fecal extracts of age-matched and co-housed wild-type (n = 5) and Map3k14-cKO (n = 5) mice using selective agar plates. c,d, ELISA of IgA (c) and pIgR (d) concentrations in fecal extracts of age-matched and co-housed wild-type (n = 7) and Map3k14-cKO (n = 7) mice. Each symbol (a–d) represents an individual mouse. Small horizontal lines (b–d) indicate the mean ( ± s.e.m.), and a is presented as box plots (where the middle line indicates the mean value and all individual plots are shown). Data were analyzed by unpaired two-tailed Student’s t test (b–d) or analyzed for multiple comparisons using the Holm–Sidak method (a), with P values shown above the plots. Data are representative of three independent experiments.
Supplementary Figure 5 Fecal IgA and pIgR concentrations and microbiota composition in wild-type and Rorc–/– mice.
a,b, ELISA analysis of IgA (a) and pIgR (b) concentrations in fecal extracts of age-matched wild-type (WT, n = 10) and Rorc–/– (n = 7) mice. c, Analysis of the commensal anaerobic bacteria Enterococcus spp. in the fecal extracts of age-matched wild-type (n = 10) and Rorc–/– (n = 7) mice by using selective agar plates. d, 16 s rRNA sequencing analysis of the genus levels of microbiota composition from age-matched wild-type (n = 5) and Rorc–/– (n = 3) mice. Each symbol (a–d) represents an individual mouse. Small horizontal lines (a–c) indicate the mean ( ± s.e.m.), and d is presented as box plots (where the middle line indicates the mean value and all individual plots are shown). Data were analyzed by unpaired two-tailed Student’s t test (a–c) or analyzed for multiple comparisons using the Holm–Sidak method (d), with P values shown above the plots. Data are representative of three independent experiments.
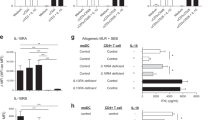
Supplementary Figure 6 NIK mediates TLR-stimulated IL-23 expression in lamina propria DCs.
a,b, qRT–PCR analysis of the indicated mRNAs in sorted wild-type (WT, n = 3) and Map3k14-cKO (n = 3) lamina propria DCs (CD45+CD11c+MHCII+) that were either incubated in medium (NT) or stimulated with the indicated TLR ligands for 6 h (a) or 12 h (b). Each symbol represents an individual mouse, and small horizontal lines indicate the mean ± s.e.m. (a,b). Data were analyzed by unpaired two-tailed Student’s t test, with P values shown above the plots. Data are representative of two independent experiments.
Supplementary Figure 7 Defective mucosal immunity and altered microbiota composition in Nfkb2Lym1/+ mice.
a, qRT–PCR analysis of Il23a mRNA in freshly sorted and untreated lamina propria DCs of wild-type (WT, n = 6) and Nfkb2Lym1/+ (n = 6) mice. b,c, ELISA of IgA (b) and pIgR (c) concentration in the feces of wild-type and Nfkb2Lym1/+ mice. d, Flow cytometric analysis of IL-17-producing CD4+ T (TH17) cells, presented as a representative FACS plot (left) and a summary graph (right) (n = 5 mice/group). e, Flow cytometric analysis of IL-17- and IL-22-producing ILC population, presented as a representative FACS plot (left) and summary graphs (right) (n = 3 mice/group). f, Analysis of commensal anaerobic bacteria Enterococcus spp. in the fecal extracts of age-matched wild-type (n = 7) and Nfkb2Lym1/+ (n = 7) mice. g, 16 S rRNA sequencing analysis of the genus levels of microbiota composition from age-matched wild-type and Nfkb2Lym1/+ mice (n = 3 mice/group). Each symbol (a–g) represents an individual mouse. Small horizontal lines (a–f) indicate the mean ( ± s.e.m.), and g is presented as box plots (where the middle line indicates the mean value and all individual plots are shown). Data were analyzed by unpaired two-tailed Student’s t test (a–f) or analyzed for multiple comparisons using the Holm–Sidak method (g), with P values shown above the plots. Data are representative of three independent experiments.
Supplementary Figure 8 Nfkb2Lym1/+ mice have impaired host defense against C. rodentium infection.
a–d, Body weight change (a), colon length (b), C. rodentium bacterial load in spleen, liver and feces (c), and histological analysis of H&E-stained colon tissue (d) of wild-type (WT) and Nfkb2Lym1/+ mice infected orally with 4 × 109 CFUs of C. rodentium (n = 4 per genotype). Scale bar, 200 μm. Each symbol represents an individual mouse, and small horizontal lines indicate the mean ( ± s.e.m.) (b,c). Data were analyzed by two-way ANOVA followed by Sidak’s multiple-comparisons test (a) or unpaired two-tailed Student’s t test (b,c). Data are representative of two independent experiments. P values are shown above plots. For a, **P = 0.006, ***P < 0.001 and ###P < 0.001.
Supplementary Figure 9 Gating strategy used in flow cytometry analysis experiments.
Live immune cell populations were first gated based on the FSC-A and SSC-A, and single cells were gated based on FSC-A and FSC-H. Immune cells were further gated based on CD45 expression. The subpopulations of the indicated immune cells were gated based on specific surface markers as indicated in the individual panels.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–9 and Supplementary Table 1
Rights and permissions
About this article

Cite this article
Jie, Z., Yang, JY., Gu, M. et al. NIK signaling axis regulates dendritic cell function in intestinal immunity and homeostasis. Nat Immunol 19, 1224–1235 (2018). https://doi.org/10.1038/s41590-018-0206-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41590-018-0206-z
This article is cited by
-
Etiopathogenesis of medication-related osteonecrosis of the jaws: a review
Journal of Molecular Medicine (2024)
-
Gut microbiota and acute kidney injury: immunological crosstalk link
International Urology and Nephrology (2023)
-
Association of Gut Microbiota with Inflammatory Bowel Disease and COVID-19 Severity: A Possible Outcome of the Altered Immune Response
Current Microbiology (2022)
-
A T cell-intrinsic function for NF-κB RelB in experimental autoimmune encephalomyelitis
Scientific Reports (2021)
-
Interaction between microbiota and immunity in health and disease
Cell Research (2020)
