
Abstract
The paradigm of cancer-targeted therapies has focused largely on inhibition of critical pathways in cancer. Conversely, conditional activation of signaling pathways as a new source of selective cancer vulnerabilities has not been deeply characterized. In this study, we sought to systematically identify context-specific gene-activation-induced lethalities in cancer. To this end, we developed a method for gain-of-function genetic perturbations simultaneously across ~500 barcoded cancer cell lines. Using this approach, we queried the pan-cancer vulnerability landscape upon activating ten key pathway nodes, revealing selective activation dependencies of MAPK and PI3K pathways associated with specific biomarkers. Notably, we discovered new pathway hyperactivation dependencies in subsets of APC-mutant colorectal cancers where further activation of the WNT pathway by APC knockdown or direct β-catenin overexpression led to robust antitumor effects in xenograft and patient-derived organoid models. Together, this study reveals a new class of conditional gene-activation dependencies in cancer.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


Data availability
Datasets for mutual exclusivity analysis: pan-can TCGA (MC3 Public version, accessed from UCSC Xena), AACR-GENIE (DFCI and MSKCC cohorts, Public v.7.0, accessed from https://www.synapse.org/#!Synapse:syn7222066/wiki/410924), Foundation One (accessed from https://gdc.cancer.gov/about-gdc/contributed-genomic-data-cancer-research/foundation-medicine/foundation-medicine) and COSMIC (https://cancer.sanger.ac.uk/cosmic/file_download_info?data=GRCh38%2Fcosmic%2Fv89%2FCosmicMutantExportCensus.tsv.gz). Please note that accessing Foundation Medicine data requires an application to dbGaP for access to the Foundation Medicine Adult Cancer Clinical Dataset (FM-AD) (study accession phs001179). Processed mutual exclusivity analysis is provided in Supplementary Table. PRISM screen data has been uploaded to GEO (raw sequencing data, GSE238126) and provided as Supplementary Table 2 (processed data). RNA-seq analysis has been uploaded to GEO (GSE232944). All genomic data from CCLE is available at https://portals.broadinstitute.org/ccle/data. DepMap 20Q4 was used for all analyses except for mutation analyses (DepMap 22Q1), Extended Data Fig. 2 (DepMap 23Q2) and Extended Data Fig. 3 (Sanger Project Score and GDSC1). Source data are provided with this paper.
Code availability
All computational analyses were performed in R (v.3.5.2). Source codes are available at https://github.com/sellerslab/SystematicPathwayActivationInCancer.
Change history
23 February 2024
A Correction to this paper has been published: https://doi.org/10.1038/s41588-024-01680-3
References
Chang, L., Ruiz, P., Ito, T. & Sellers, W. R. Targeting pan-essential genes in cancer: challenges and opportunities. Cancer Cell 39, 466–479 (2021).
Mauro, M. J. & Druker, B. J. STI571: targeting BCR-ABL as therapy for CML. Oncologist 6, 233–238 (2001).
Paez, J. G. et al. EGFR mutations in lung cancer: correlation with clinical response to Gefitinib therapy. Science 304, 1497–1500 (2004).
Flaherty, K. T. et al. Combined BRAF and MEK inhibition in melanoma with BRAF V600 mutations. N. Engl. J. Med. 367, 1694–1703 (2012).
Canon, J. et al. The clinical KRAS(G12C) inhibitor AMG 510 drives anti-tumour immunity. Nature 575, 217–223 (2019).
Mavrakis, K. J. et al. Disordered methionine metabolism in MTAP/CDKN2A-deleted cancers leads to dependence on PRMT5. Science 351, 1208–1213 (2016).
Chan, E. M. et al. WRN helicase is a synthetic lethal target in microsatellite unstable cancers. Nature 568, 551–556 (2019).
Neggers, J. E. et al. Synthetic lethal interaction between the ESCRT paralog enzymes VPS4A and VPS4B in cancers harboring loss of chromosome 18q or 16q. Cell Rep. 33, 108493 (2020).
Rouleau, M., Patel, A., Hendzel, M. J., Kaufmann, S. H. & Poirier, G. G. PARP inhibition: PARP1 and beyond. Nat. Rev. Cancer 10, 293–301 (2010).
Unni, A. M., Lockwood, W. W., Zejnullahu, K., Lee-Lin, S. Q. & Varmus, H. Evidence that synthetic lethality underlies the mutual exclusivity of oncogenic KRAS and EGFR mutations in lung adenocarcinoma. eLife 4, e06907 (2015).
Ito, T. et al. Paralog knockout profiling identifies DUSP4 and DUSP6 as a digenic dependence in MAPK pathway-driven cancers. Nat. Genet. 53, 1664–1672 (2021).
Leung, G. P. et al. Hyperactivation of MAPK signaling is deleterious to RAS/RAF-mutant melanoma. Mol. Cancer Res. 17, 199–211 (2019).
Das Thakur, M. et al. Modelling vemurafenib resistance in melanoma reveals a strategy to forestall drug resistance. Nature 494, 251–255 (2013).
Seong, B. K. A. et al. TRIM8 modulates the EWS/FLI oncoprotein to promote survival in Ewing sarcoma. Cancer Cell 39, 1262–1278.e7 (2021).
Gallo, D. et al. CCNE1 amplification is synthetic lethal with PKMYT1 kinase inhibition. Nature 604, 749–756 (2022).
The AACR Project GENIE Consortium. et al. AACR Project GENIE: powering precision medicine through an international consortium. Cancer Discov. 7, 818–831 (2017).
Sanchez-Vega, F. et al. Oncogenic signaling pathways in the cancer genome atlas. Cell 173, 321–337.e10 (2018).
Hartmaier, R. J. et al. High-throughput genomic profiling of adult solid tumors reveals novel insights into cancer pathogenesis. Cancer Res. 77, 2464–2475 (2017).
Mina, M. et al. Conditional selection of genomic alterations dictates cancer evolution and oncogenic dependencies. Cancer Cell 32, 155–168.e6 (2017).
Yu, C. et al. High-throughput identification of genotype-specific cancer vulnerabilities in mixtures of barcoded tumor cell lines. Nat. Biotechnol. 34, 419–423 (2016).
Corsello, S. M. et al. The Drug Repurposing Hub: a next-generation drug library and information resource. Nat. Med. 23, 405–408 (2017).
Tsherniak, A. et al. Defining a cancer dependency map. Cell 170, 564–576.e16 (2017).
Basu, A. et al. An interactive resource to identify cancer genetic and lineage dependencies targeted by small molecules. Cell 154, 1151–1161 (2013).
Ghandi, M. et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature 569, 503–508 (2019).
Duffy, M. J., Tang, M., Rajaram, S., O’Grady, S. & Crown, J. Targeting mutant p53 for cancer treatment: moving closer to clinical use? Cancers 14, 4499 (2022).
Chen, Y.-N. P. et al. Allosteric inhibition of SHP2 phosphatase inhibits cancers driven by receptor tyrosine kinases. Nature 535, 148–152 (2016).
Lito, P. et al. Relief of profound feedback inhibition of mitogenic signaling by RAF inhibitors attenuates their activity in BRAFV600E melanomas. Cancer Cell 22, 668–682 (2012).
Yang, W. et al. Genomics of Drug Sensitivity in Cancer (GDSC): a resource for therapeutic biomarker discovery in cancer cells. Nucleic Acids Res. 41, D955–D961 (2013).
Behan, F. M. et al. Prioritization of cancer therapeutic targets using CRISPR–Cas9 screens. Nature 568, 511–516 (2019).
Juric, D. et al. Convergent loss of PTEN leads to clinical resistance to a PI(3)Kα inhibitor. Nature 518, 240–244 (2015).
Oda, K., Stokoe, D., Taketani, Y. & McCormick, F. High frequency of coexistent mutations of PIK3CA and PTEN genes in endometrial carcinoma. Cancer Res. 65, 10669–10673 (2005).
Matulonis, U. et al. Phase II study of the PI3K inhibitor pilaralisib (SAR245408; XL147) in patients with advanced or recurrent endometrial carcinoma. Gynecol. Oncol. 136, 246–253 (2015).
Myers, A. P. et al. Phase II, two-stage, two-arm, PIK3CA mutation stratified trial of MK-2206 in recurrent endometrial cancer (EC). J. Clin. Oncol. 31, 5524–5524 (2013).
Yaeger, R. et al. Clinical sequencing defines the genomic landscape of metastatic colorectal cancer. Cancer Cell 33, 125–136.e3 (2018).
Lepourcelet, M. et al. Small-molecule antagonists of the oncogenic Tcf/β-catenin protein complex. Cancer Cell 5, 91–102 (2004).
Liao, H. et al. A PROTAC peptide induces durable β-catenin degradation and suppresses Wnt-dependent intestinal cancer. Cell Discov. 6, 35 (2020).
Grossmann, T. N. et al. Inhibition of oncogenic Wnt signaling through direct targeting of β-catenin. Proc. Natl Acad. Sci. USA 109, 17942–17947 (2012).
Levine, A. J., Jenkins, N. A. & Copeland, N. G. The roles of initiating truncal mutations in human cancers: the order of mutations and tumor cell type matters. Cancer Cell 35, 10–15 (2019).
Christie, M. et al. Different APC genotypes in proximal and distal sporadic colorectal cancers suggest distinct WNT/β-catenin signalling thresholds for tumourigenesis. Oncogene 32, 4675–4682 (2013).
Albuquerque, C. et al. The ‘just-right’ signaling model: APC somatic mutations are selected based on a specific level of activation of the β-catenin signaling cascade. Hum. Mol. Genet. 11, 1549–1560 (2002).
Ranes, M., Zaleska, M., Sakalas, S., Knight, R. & Guettler, S. Reconstitution of the destruction complex defines roles of AXIN polymers and APC in β-catenin capture, phosphorylation, and ubiquitylation. Mol. Cell 81, 3246–3261.e11 (2021).
Schneikert, J., Grohmann, A. & Behrens, J. Truncated APC regulates the transcriptional activity of β-catenin in a cell cycle dependent manner. Hum. Mol. Genet. 16, 199–209 (2007).
The Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 487, 330–337 (2012).
Golshani, G. & Zhang, Y. Advances in immunotherapy for colorectal cancer: a review. Ther. Adv. Gastroenterol. 13, 1756284820917527 (2020).
Nusse, R. & Clevers, H. Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell 169, 985–999 (2017).
Krishnamurthy, N. & Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: update on effectors and inhibitors. Cancer Treat. Rev. 62, 50–60 (2018).
Jung, Y.-S. & Park, J.-I. Wnt signaling in cancer: therapeutic targeting of Wnt signaling beyond β-catenin and the destruction complex. Exp. Mol. Med. 52, 183–191 (2020).
Sun, C. et al. Reversible and adaptive resistance to BRAF(V600E) inhibition in melanoma. Nature 508, 118–122 (2014).
Unni, A. M. et al. Hyperactivation of ERK by multiple mechanisms is toxic to RTK-RAS mutation-driven lung adenocarcinoma cells. eLife 7, e33718 (2018).
Saghafinia, S. et al. Conditional selection of genomic alterations dictates cancer evolution and oncogenic dependencies. Cancer Cell 32, 155–168.e6 (2017).
Soumerai, T. E. et al. Clinical utility of prospective molecular characterization in advanced endometrial cancer. Clin. Cancer Res. 24, 5939–5947 (2018).
Bailey, M. H. et al. Comprehensive characterization of cancer driver genes and mutations. Cell 173, 371–385.e18 (2018).
Dou, Y. et al. Proteogenomic characterization of endometrial carcinoma. Cell 180, 729–748.e26 (2020).
Cerami, E. et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2, 401–404 (2012).
Sweeney, S. M. et al. AACR project genie: powering precision medicine through an international consortium. Cancer Discov. 7, 818–831 (2017).
Corsello, S. M. et al. Discovering the anticancer potential of non-oncology drugs by systematic viability profiling. Nat. Cancer 1, 235–248 (2020).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015).
Cornwell, M. et al. VIPER: visualization Pipeline for RNA-seq, a Snakemake workflow for efficient and complete RNA-seq analysis. BMC Bioinf. 19, 135 (2018).
Sato, T. et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 141, 1762–1772 (2011).
Acknowledgements
We thank K. Polyak, S. Elledge, M. Meyerson and the members of the Sellers laboratory for helpful discussions. We thank Broad Institute Genomics Platform, Broad Institute PRISM Laboratory, Broad Institute Comparative Medicine and DFCI Molecular Biology Core Facilities for their technical support.
Author information
Authors and Affiliations
Contributions
L.C. and W.R.S. conceived the studies. L.C., M.G.R., M.R. and J.A.R. designed and executed the PRISM GOF screen. L.C., D.K. and R.L. performed bioinformatic and statistical analysis. L.C., N.Y.J. and P.R. performed in vitro validation experiments and analysis. D.J.R., L.C., N.Y.J., S.C. and T.-Y.S. performed mouse xenograft experiments. A.A. and Y.-Y.T. performed patient-derived organoid validation experiments. T.I. and L.M.v.T. provided critical insights into experimental design and interpretation. L.C. and W.R.S. wrote the paper, with critical reading and feedback from other coauthors.
Corresponding author
Ethics declarations
Competing interests
W.R.S. is a Board or SAB member and holds equity in Ideaya Biosciences, Civetta Therapeutics, Red Ridge Bio, Delphia Therapeutics, Scorpion Therapeutics and 2Seventy Bio. Delphia Therapeutics is exploring GOF therapeutics in cancer. W.R.S. has consulted for Array, Astex, Epidarex Capital, Ipsen, PearlRiver Therapeutics, Merck Pharmaceuticals, Sanofi, Servier and Syndax Pharmaceuticals and receives research funding from Pfizer Pharmaceuticals, Merck Pharmaceuticals, Ideaya Biosciences, Calico, Boehringer-Ingelheim, Bristol Myers Squibb, Bayer Pharmaceuticals, Novartis Institutes for Biomedical Research and Ridgeline Discovery. W.R.S. is a copatent holder on EGFR mutation diagnostic patents. L.C. is an employee of Cure Ventures and was a previous employee of 5AM Ventures and Flagship Pioneering. T.I. is an equity holder of Scorpion Therapeutics and Odyssey Therapeutics. All other authors declare no competing interests.
Peer review
Peer review information
Nature Genetics thanks Kris Wood and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Assay development and quality control of the Gain-of-Function ORF screen in 488 cell lines.
Correlation of cell line barcode reads between barcode PCR technical replicates (n = 2) across all ORF conditions (n = 11). Y-axis represents the Pearson correlation coefficients, and each bar represents a biological condition.
Extended Data Fig. 2 Relationship between the viability effects of TSG overexpression and corresponding mutational and functional biomarkers in 488 cancer cell lines.
The Y-axis represents the Chronos score of gene CRISPR knockout dependencies in each cell line. The X-axis represents the log-fold change (LogFC) of each cell line resulting from ORF overexpression compared to GFP overexpression. a, Relationship between the viability effects of p53 ORF overexpression (X-axis) and the viability effects of TP53 knockout (Y-axis) in 488 cancer cell lines. Each dot represents a cancer cell line colored by TP53 hotspot mutation status as indicated. b, Relationship between the viability effects of p16 ORF overexpression (X-axis) and the viability effects of CDK6 knockout (Y-axis) in 488 cancer cell lines. Each dot represents a cancer cell line colored by RB1 hotspot mutation status as indicated. c, Relationship between the viability effects of PTEN ORF overexpression (X-axis) and the viability effects of PIK3CA knockout (Y-axis) in 488 cancer cell lines. Each dot represents a cancer cell line colored by PIK3CA hotspot mutation status as indicated.
Extended Data Fig. 3 Vulnerabilities of ERK2 overexpression in NRAS-mutant cells.
Box plot showing differential ERK2 overexpression according to BRAF or NRAS hotspot/recurrence mutation status in 488 cell lines. The Y-axis represents the log-fold change (LogFC) of each cell line upon ERK2 overexpression compared with GFP overexpression. P-values by unpaired two-sided t-test. The box in this figure shows the median (the middle line) and the 25–75th percentiles (the box); the whiskers show 1.5× the interquartile range from the lower and upper quartile.
Extended Data Fig. 4 Cross-validation of context-specific vulnerabilities upon p53, p16, PTEN, and ERK2 overexpression across 488 cell lines using Project Score and Sanger GDSC datasets.
a, b, Genetic and drug perturbation features associated with differential viability effects resulting from p53 overexpression. (a) Pairwise correlation of the viability effects of p53 overexpression with ~20,000 CRISPR dependency features (Project Score) in 152 overlapping cell lines. The Y-axis represents the statistical significance (-log10_pvalue) of each pair of associations from the Pearson correlation analysis, and the X-axis represents the Pearson correlation coefficient for each association pair. (b) Correlation between ~300 compound sensitivity (Sanger GDSC1 dataset) features and p53 overexpression viability effects across ~300 cell lines. Axes are as shown in a. c, d, Genetic and drug perturbation features associated with differential viability effects resulting from p16 overexpression. (c) Pairwise correlation of the viability effects of p16 overexpression with ~20,000 CRISPR dependency features (Project Score) in 152 cell lines. The Y-axis represents the statistical significance (-log10_pvalue) of each pair of associations from the Pearson correlation analysis, and the X-axis represents the Pearson correlation coefficient for each association pair. (d) Correlation between ~300 compound sensitivity (Sanger GDSC1 dataset) features and p16 overexpression viability effects across ~300 cell lines. Axes are as shown in a. e, f, Genetic and drug perturbation features associated with differential viability effects resulting from (e) PTEN and (f) overexpression. Pairwise correlation between ~300 compound sensitivity (Sanger GDSC1 dataset) features and PTEN overexpression viability effects across ~300 cell lines. The Y-axis represents the statistical significance (-log10_pvalue) of each pair of associations from the Pearson correlation analysis, and the X-axis represents the Pearson correlation coefficient for each association pair. Colors of points represent positive (brown) and negative (purple) Pearson correlation coefficients. ‘Inhibitor’ is denoted by ‘i’ in point labels.
Extended Data Fig. 5 Immunoblot detection of ectopically expressed AKT1 in a panel of endometrial cancer cell lines.
Immunoblot of indicated proteins from extracts prepared from the indicated cell lines ectopically expressing GFP, AKT1E17K and AKT1E17K,K179A (all with V5 tag). Each experiment was repeated independently two times. Row gel scanning images are provided in Source Data Extended Data Fig. 1.
Extended Data Fig. 6 β-catenin overexpression, APC/CSNK1A1 knockdown, and changes in WNT signaling activation in colorectal cancer cell lines.
a, Quantitative RT-qPCR of indicated mRNAs in cells after transduction of non-targeting shRNA (NT1) and anti-APC shRNAs (APC-shRNA#1 and #3). The Y-axis represents the expression level of each indicated gene relative to NT1 control. Bars represent means and the error bars represent standard deviations in each group across n = 3 biologically independent samples. b, Quantitative RT-qPCR of indicated mRNAs in cells after transduction of non-targeting shRNA (NT1) and anti-CSNK1A1 shRNA in HT29 (left) and RKO (right) cells. The Y-axis represents the expression level of each indicated gene relative to NT1 control. Bars represent means and the error bars represent standard deviations in each group across n = 3 biologically independent samples.
Extended Data Fig. 7 RNA-seq results in HT29 cell line following WNT hyperactivation.
a, b, Volcano plots showing the top significantly up- and down-regulated genes detected by RNAseq comparing (a) APC-shRNA-transduced and NT-shRNA-transduced and (b) GFP-transduced to β-catenin-transduced HT29 cells. The Y-axis represents the statistical significance (-log10_pvalue) of each gene, and the X-axis represents the log2 (fold change) for each gene. Selective genes related to the WNT or cell cycle pathways are labeled. Colors of dots are annotated as: gene with statistically significant positive log2 fold change (red), negative fold change (grey), or no significant change (black). Dotted line represents the statistical significance cutoff of p = 0.01. c, Gene-Set Enrichment Analysis (GSEA) showing selective significantly enriched and depleted pathways related to cell cycle and WNT signaling in GFP compared to β-catenin transduced HT29 cells. The X-axis represents the Normalized Enrichment Score of each gene module. The adjusted p value for each gene module is indicated.
Extended Data Fig. 8 Apoptosis and senescence profiling of colorectal cancer cell lines after WNT hyperactivation.
a, b, FACS-based detection of Annexin V-FITC stained RKO and HT29 cells indicating apoptotic cells, (a) 5 days after β-catenin or β-catenin-D164AΔC (non-TCF-binding negative control) overexpression, and (b) 4 days after APC shRNA or non-targeting shRNA infection (including 2 days of doxycycline induction). The percentage of Annexin V-FITC positive cells relative to total analyzed cells in each replicate is indicated. c, d, FACS-based detection of β-Gal-stained RKO and HT29 cells indicating senescent cells, (c) 5 days after β-catenin or β-catenin-D163AΔC (non-TCF-binding negative control) overexpression, or (d) 2 days after APC shRNA or non-targeting shRNA induction (including 2 days of doxycycline induction). Empty circles indicate no addition of β-Gal staining substrate, and solid circle indicate addition of β-Gal staining substrate. e-f, FACS gating strategies for (e) Annexin V-FITC staining in a-b, and (f) β-Gal staining in c-d.
Extended Data Fig. 9 Transduction of patient-derived colorectal cancer organoid via lentiviral infection using GFP encoding viruses.
a, APC mutation status of 3 patient-derived organoids. b, representative pictures of 3 patient-derived organoids after transduction of a lentivirus encoding GFP. Experiment was performed once because of the assay development nature.
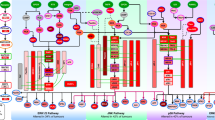
Extended Data Fig. 10 Schematic summary of cancer context-specific pathway activation lethality highlighted in this study.
In cancer models with specific driver mutations that activate certain oncogenic pathways (MAPK, PI3K, WNT), the elevations of background signaling make them susceptible to loss of viability mediated by further activations of the initially altered signaling pathway.
Supplementary information
Supplementary Tables 1 and 2.
Mutual exclusivity analysis of genome mutations in 69,393 patients with cancer using the SELECT algorithm; logFC scores of the ORF overexpression screen (normalized to GFP).
Source data
Source Data Extended Data Fig. 5
Unprocessed western blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chang, L., Jung, N.Y., Atari, A. et al. Systematic profiling of conditional pathway activation identifies context-dependent synthetic lethalities. Nat Genet 55, 1709–1720 (2023). https://doi.org/10.1038/s41588-023-01515-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-023-01515-7
This article is cited by
-
Hyperactivation of oncogenic driver pathways as a precision therapeutic strategy
Nature Genetics (2023)
