
Abstract
More than a quarter of the world’s tropical forests are exploited for timber1. Logging impacts biodiversity in these ecosystems, primarily through the creation of forest roads that facilitate hunting for wildlife over extensive areas. Forest management certification schemes such as the Forest Stewardship Council (FSC) are expected to mitigate impacts on biodiversity, but so far very little is known about the effectiveness of FSC certification because of research design challenges, predominantly limited sample sizes2,3. Here we provide this evidence by using 1.3 million camera-trap photos of 55 mammal species in 14 logging concessions in western equatorial Africa. We observed higher mammal encounter rates in FSC-certified than in non-FSC logging concessions. The effect was most pronounced for species weighing more than 10 kg and for species of high conservation priority such as the critically endangered forest elephant and western lowland gorilla. Across the whole mammal community, non-FSC concessions contained proportionally more rodents and other small species than did FSC-certified concessions. The first priority for species protection should be to maintain unlogged forests with effective law enforcement, but for logged forests our findings provide convincing data that FSC-certified forest management is less damaging to the mammal community than is non-FSC forest management. This study provides strong evidence that FSC-certified forest management or equivalently stringent requirements and controlling mechanisms should become the norm for timber extraction to avoid half-empty forests dominated by rodents and other small species.
Similar content being viewed by others

Main
Commercial logging concessions cover more than one-quarter of the world’s remaining tropical forests1. Forest certification schemes aim to have more positive socio-economic and environmental outcomes compared to conventional logging schemes. For example, the Forest Stewardship Council (FSC) aims to reduce direct environmental impacts by various means that include maintaining high conservation value forests and applying reduced impact logging practices (Supplementary Tables 1 and 2). A major concern for biodiversity is that timber extraction—by the creation of roads—opens previously remote forests, enabling illegal and unsustainable hunting4,5,6,7. This indirect effect of logging is known to mainly influence medium- to large-sized forest mammals, which are particularly vulnerable to human pressure8. FSC certification may alleviate these pressures because, among other measures, companies reduce accessibility to concessions by closing off old logging roads, prohibit wild meat transport and hunting materials, provide access to alternative meat sources for workers and their families, and carry out surveillance by rangers. An FSC certificate is valid for 5 years and logging companies are audited for compliance through third-party annual surveillance assessments.
In African tropical forests, FSC certification has been shown to be associated with reduced deforestation9, improved working and living conditions of employees and benefit-sharing with neighbouring institutions10. Studies in Latin America suggest that mammal occupancy in FSC-certified sites is comparable to that of protected areas11,12. There is, however, little data on the status of faunal communities in FSC-certified versus non-FSC forests2,3. Most studies on the effectiveness of FSC certification for wildlife conservation have focused on one or a few sites or species at a time13,14,15,16. Although these studies reported a positive impact of FSC certification on wildlife compared to non-FSC concessions, their research designs did not account for explanatory variables such as concession location, land-use history or stochastic effects17,18. One study included several sites and species and found no effect of FSC certification19. However, that study investigated only bird species richness: bird dispersal distances are much higher than those of terrestrial mammals and may thus be a weak indicator of local management. In addition to simply comparing species diversity, it is important to compare population sizes between forest management types. Hunting does not necessarily completely extirpate wildlife species, especially when forests are connected, but rather results in population declines4.
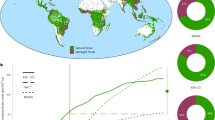
We used camera traps to assess whether FSC certification can mitigate the negative effects of timber extraction on wildlife by studying the encounter rate of a broad range of mammal species across several sites. We compared small- to large-sized mammal observations across seven paired FSC-certified and non-FSC concessions in Gabon and the Republic of Congo (Fig. 1). Gabon and the Republic of Congo lie in western equatorial Africa (WEA). We included all companies that were FSC-certified between 2018 and 2021 in this region, except for one which refused to allow access. WEA is particularly suitable for these analyses, as its forests are reasonably intact and logging concessions are embedded in a matrix of contiguous forest, which are therefore mostly devoid of influences other than the effects of logging and hunting20. Wild meat hunting is pervasive throughout WEA, whereby logging increases hunting pressure by increasing access (logging roads) and through the arrival of people working in the concessions in once-remote forests8. By ensuring spatial pairing of the FSC-certified and non-FSC concessions we minimized the influence of regional landscape heterogeneity. We calculated mammal encounter rates and grouped mammal species into five body mass classes. The relative encounter rate of these classes could be used as a proxy for hunting pressure, as larger-bodied species are targeted more by hunters8. In addition, larger-bodied species recover more slowly from hunting compared to smaller-bodied species, resulting in lower abundances of large versus small species under higher hunting pressure21,22. Finally, we explored how FSC-certified forest management affects mammal encounter rate by taxonomic group and by IUCN Red List categories. We hypothesized that FSC certification would effectively decrease hunting pressure and therefore predicted a higher encounter rate of larger-bodied species in FSC-certified compared with non-FSC logging concessions.

Between 28 to 36 cameras were deployed in each concession in systematic, 1 km spaced grids. Numbers and lines indicate the pairs of FSC-certified and non-FSC concessions.
We collected and catalogued nearly 1.3 million photos from 474 camera-trap locations for a total of 35,546 days, averaging 2,539 camera-trap days per concession (Extended Data Table 1). We detected a total of 55 mammal species (Extended Data Table 2). The mammal encounter rate estimated by our model (Fig. 2a) was 1.5 times higher in FSC-certified concessions compared to non-FSC concessions (Extended Data Table 3). We also found fewer signs of hunting (Fig. 2b) in FSC-certified than in non-FSC concessions. Estimated total faunal biomass derived from mammal encounter rates was 4.5 times higher in FSC-certified compared to non-FSC concessions (Extended Data Fig. 1). Larger species contributed more to the total biomass. We observed comparable species diversity in the two concession types, as only a few species, all with very low encounter rates, were lacking completely in one or other of the concession types (Extended Data Table 2).

a,b, Encounter rate of all observed mammals (P = 0.041) (a) and proportion of camera locations with hunting signs (P = 0.036) (b). Numbers represent paired FSC-certified (n = 7) and non-FSC (n = 7) concessions. The red line in a represents the linear mixed model predicted fixed effect (certification status) and grey lines represent random effects (concession pairs). Differences between hunting signs in b were analysed using a two-sided Wilcoxon signed-rank test. Data are represented as boxplots, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. *P < 0.05.
The differences between mammal encounter rates in FSC-certified and non-FSC concessions increased with body mass (Fig. 3 and Extended Data Tables 3 and 4). FSC-certified concessions had higher encounter rates of mammals above 10 kg than non-FSC concessions but there was no difference for mammals below 10 kg. Model estimates showed that mammals in body mass classes over 100, 30–100 and 10–30 kg, had encounter rates that were 2.7, 2.5 and 3.5 times higher, respectively, in FSC-certified concessions compared to non-FSC concessions. Mammal encounter rates of the IUCN Red List categories critically endangered, near threatened and least concern were 2.7, 2.3 and 1.4 times higher, respectively, in FSC-certified compared to non-FSC concessions (Fig. 4 and Extended Data Tables 3 and 4).

Numbers represent paired FSC-certified (n = 7) and non-FSC (n = 7) concessions, red lines represent linear mixed model predicted fixed effects (certification status) and grey lines represent random effects (concession pairs). Data are represented as boxplots, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. Pairwise comparisons were multivariate t adjusted. ***P < 0.001. Exact P values are summarized in Extended Data Table 3. Note that the scales of the y axes vary. Silhouettes of Gorilla gorilla, Syncerus caffer, Potamochoerus porcus, Cephalophus sp., Hyemoschus aquaticus, Philantomba monticola, Atherurus africanus and mice were created by T. Markus.

Numbers represent paired FSC-certified (n = 7) and non-FSC (n = 7) concessions, red lines represent linear mixed model predicted fixed effects (certification status) and grey lines represent random effects (concession pairs). Data are represented as boxplots, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. Pairwise comparisons were multivariate t adjusted. ***P < 0.001, *P < 0.05. Exact P values are summarized in Extended Data Table 3. Note that the scales of the y axes vary.
Mammal encounter rate in FSC-certified and non-FSC concessions varied between taxonomic groups (Fig. 5 and Extended Data Tables 3 and 4). In FSC-certified concessions, forest elephants were encountered 2.5 times, primates 1.8 times, even-toed ungulates 2 times and carnivores 1.5 times more compared to non-FSC concessions. The encounter rate of pangolins and rodents did not differ.

Numbers represent paired FSC-certified (n = 7) and non-FSC (n = 7) concessions, red lines represent linear mixed model predicted fixed effects (certification status) and grey lines represent random effects (concession pairs). Data are represented as boxplots, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. Pairwise comparisons were multivariate t adjusted. ***P < 0.001, **P < 0.01, *P < 0.05. Exact P values are summarized in Extended Data Table 3. Note that the scales of the y axes vary.
Discussion
The loss of large mammals
We conducted a large-scale quantitative study to assess the impact of FSC-certified forest management on mammal encounter rate across several logging concessions and for a broad range of mammals. Our data provide strong evidence that FSC-certified forest management results in higher overall mammal abundance, as approximated by encounter rate and faunal biomass relative to non-FSC forest management. This effect was most pronounced for species larger than 10 kg, which was consistent for all FSC–non-FSC concession pairs, probably because these medium to large species recover more slowly from population losses and may be targeted more often by hunters21,22. Not all large species with reduced encounter rates may be commonly targeted for hunting but they are often indiscriminately affected by snaring23. Non-FSC concessions contained proportionally more rodents and other small species than did FSC-certified concessions (Extended Data Table 2). The lack of hunting impacts on small mammal populations suggests some form of density compensation is in place: the hunting pressure on small mammal populations might be compensated by higher reproductive rates and/or a release from competition and predation in the non-FSC concessions24,25.
A particularly strong effect of FSC certification was found for the critically endangered forest elephant, which is in line with previous findings14. The distribution of this species is driven almost entirely by human activity: they avoid areas that are unsafe to them26,27. Their large home ranges can span several concessions28, thus they may actively seek to reside not only in protected areas but also in FSC-certified concessions where measures to prevent illegal hunting are in place. This suggests that FSC-certified concessions may provide an important refuge for wide-ranging elephants. By contrast, no difference was found in pangolin encounter rate (they are among the most trafficked mammals29) between the two types of logging regimes. Two out of the three pangolin species present in WEA are relatively small and generally have higher reproduction rates than mammals in larger size classes. Moreover, all three pangolin species had low encounter rates in our study (Extended Data Table 2), probably because two pangolin species are semi-arboreal and are therefore not effectively captured by ground-based camera traps, which reduces our ability to draw strong conclusions about these species and warrants further research. We did not observe a loss of species that were encountered frequently in either FSC-certified or non-FSC concessions, nor did we expect to. This is because human population density in WEA is relatively low and the forests are still highly connected20.
Conservation of large mammals through FSC certification brings wider benefits to forests, as these mammals play a pivotal role in ecological processes, including seed dispersal, seed predation, browsing, trampling, plant competition, nutrient cycling and predator–prey interactions30. It has also been suggested that forest carbon storage is higher when large mammal assemblages are more intact because the ecological processes they are part of (such as seed dispersal) often benefit large, high wood density trees31,32,33 and the benefits of their conservation may far outweigh the cost34. Futhermore, by reducing the amount of wild meat available for human consumption, FSC-certified concessions or similar stringent schemes may also reduce the chance of zoonotic disease transmission35.
Methodological considerations
The FSC takes a comprehensive and all-encompassing perspective when it comes to managing and promoting sustainable forest management practices. This approach recognizes that forests are complex ecosystems with intricate interconnections between their various components, including flora, fauna, soil, water and climate. In logged tropical forests, controlling hunting is probably the most important factor for the reduction of environmental impacts7. We found more hunting signs in non-FSC concessions, which supports the interpretation that FSC effectively reduces hunting pressure, although counting hunting signs is likely to be a relatively weak measure of the quantification of hunting pressure36. Hunting has long been known to be the most important driver of forest fauna decline in central African logged forests6,37 and the same phenomenon has been shown in Asia7. Of course, other factors such as retaining high conservation value areas and reduced impact logging practices are likely to contribute to the observed effects as well38. Our data do not allow for causal inference of the association of any of the specific measures implemented by FSC companies with the observed effects, as that would require setting up more detailed measure-based experiments.
For the sections of the concessions that we sampled, we ensured comparability between paired concessions. We maximized the similarity in geographic covariates that may drive variation in mammal abundance—elevation and distances to roads, rivers, human settlements and protected areas—between each pair of FSC-certified and non-FSC concessions (Extended Data Fig. 2 and Extended Data Table 5). Although we believe that these covariates are important drivers of mammal abundance39, including these covariates did not greatly improve the models, which underscores that camera grid locations were sufficiently similar in terms of these confounding influences. Precise logging intensity and logging history data per camera were not available for most concessions because the planning schemes of companies and actual exploitation of cutting blocks often did not match. Slight differences in logging history are not expected to have a large effect on the data because mammals are mobile and can return quickly to areas that have been exploited40. Fourteen logging concessions may be a large sample size for tropical ecology studies17 but a low sample size from a statistical perspective. Nonetheless, despite the small number of replicates, we found clear and consistent differences in encounter rate between FSC-certified and non-FSC forests.
We used encounter rate, defined as the number of observations divided by the number of camera-trap days. Encounter rates may be affected by unaccounted influences on detection probabilities41, which may complicate comparisons between species or between sites. We compare individual species across management types, which renders differences in detection across species less relevant. For camera-trap sites, however, variation in visibility or other factors may affect the number of detections, even though mammal population sizes are similar. However, we found no differences in any relevant site covariates between treatments at the camera-trap level. Visibility at ground level, slope, the presence of fruiting trees and small water courses around camera-trap locations did not differ between FSC-certified and non-FSC concessions (Extended Data Fig. 3 and Extended Data Table 5). We also compared the presence and type of trails or paths around camera-trap locations, which did not differ significantly except for the number of elephant paths, which was higher in FSC-certified concessions (Extended Data Fig. 4 and Extended Data Table 5). As camera traps were installed randomly at the predetermined GPS locations on the nearest tree with 4 m visibility, finding a higher frequency of elephant paths in FSC-certified concessions was, in itself, an indication of higher elephant abundance in FSC-certified concessions. Potential seasonal influences are accounted for by the paired design. It is, however, important to note that encounter rates are a mixed measure of abundance and activity and we cannot disentangle whether changes in encounter rate are the result of changes in abundance, activity—movement per day—or both. Species’ home ranges and movement patterns can change in response to disturbance, which can affect encounter rates. It is, however, unlikely that changes in activity solely make up the observed differences in encounter rates, given the consistency of the data in the three heaviest body mass classes. We also estimate relative biomass using encounter rates, which is a useful proxy to assess differences between forest management types but cannot be interpreted as true biomass (Extended Data Fig. 1).
Conservation implications
Of central African tropical forest, 21% is designated for protection but only 15% of the species’ ranges for central chimpanzees and the western lowland gorilla lie in protected areas42,43. More than half of these species’ ranges and a large part of the ranges of other mammals, such as forest elephants, lie in logging concessions26. Protected areas are essential for conservation but sometimes lack the resources for effective control of illegal hunting44,45. Logging companies often do have the means to protect forests and have an economic incentive to do so. We did not compare mammal encounter rates in protected areas with the same metric in logging concessions ourselves. However, our observed encounter rates for large mammals, which are the first species to disappear as a result of hunting and poaching, in FSC-certified concessions were comparable to published data from recently monitored protected areas in the same region46,47,48. The ratio of large versus small forest antelopes in the FSC-certified concessions is furthermore comparable to such ratios in a protected area in the region with almost no hunting, whereas those in non-FSC concessions are far lower49. Although the first priority for species protection should be to maintain unlogged forests where there is effective law enforcement, our results challenge the notion that, at least for large-bodied mammals in WEA, logging is always disastrous for wildlife50,51. We show that, if selectively logged forests are properly managed, they can provide an important contribution to biodiversity: our results confirm that FSC-certified forests support far more larger and threatened species than do non-FSC forests. The results of this study are likely to be applicable to other logged tropical forests where hunting, through increased accessibility, poses a risk to forest mammals. This is because wildlife protection measures and law enforcement are applied across all FSC-certified forests, as part of the FSC principles, criteria and indicators for which FSC-certified companies are audited for compliance (Supplementary Tables 1 and 2). We infer this with caution as timber extraction volumes, concession size and shape, presence of public roads, population density and other characteristics may differ between concessions and thereby affect the impacts of FSC-certified forest management52.
Most terrestrial protected areas are isolated53 and increasing human modification and fragmentation of landscapes is limiting the ability of mammals to move54. Governments in forest-rich countries may enhance the effectiveness of conservation policies by requiring FSC certification in strategic locations, such as buffer zones around protected areas to reduce the edge-to-area ratio of the conservation landscape55. Non-FSC companies may also contribute to conservation, as they vary along a gradient of environmental and social responsibility56. This was, however, not the focus of our study. Concessions in our study region are large, often larger than 2,000 km2, and together with protected areas they can substantially contribute to mammal conservation. Well-managed logging concessions can contribute to Sustainable Development Goal (SDG) 12 (sustainable consumption and production) and SDG 15 (life on land) by performing a strategic function in preserving habitats and landscape connectivity while allowing for responsible economic activity57.
Our findings indicate that the requirements of FSC certification lead to effective mitigation of direct and indirect influences of logging on tropical forest mammals. The control of widespread and unsustainable hunting and poaching which is facilitated by the increased access to forests engendered by timber extraction is probably a key determinant of this impact. However, not all hunting is illegal and FSC certification protects customary rights to hunt non-protected species for subsistence. Sustainability of this practice is controlled by—among other requirements—controlling firearm permits, spatially assigning hunting zones and monitoring wildlife offtake. We believe that a strict set of requirements, control of compliance and regular enforcement, all integrally connected and ensured in the FSC system, are crucial for successful environmental protection through forest certification.
The need to upscale certification
We present a clear, evidence-based message about the positive impact of FSC certification. We show that medium- to large-sized mammals—which play vital functions in forests—are more abundant in FSC-certified concessions than in non-FSC concessions. This study calls for action, reinforcing previous studies that called for more forest certification and land-use planning that takes conservation into account14,26,43,58. To protect large mammals, we urge that FSC certification or similar stringent schemes become the norm, as conventional logging is likely to result in half-empty forests dominated by rodents and other small species. To increase logging companies’ interest in FSC certification, it is essential that sufficient demand is created for FSC-certified products by institutional and individual buyers. The information put forward by this study can play an important role in FSC’s global strategy to leverage sustainable finance to reduce biodiversity loss, whereby certificate holders can be rewarded for the biodiversity benefits that they incur59. Rendering FSC-certified forests eligible for payments by biodiversity schemes, especially driven by government regulation60, can contribute to fair valuation of standing forests. To ensure environmentally and socially responsible forest management practices10, we strongly support the application of regulatory frameworks which stimulate and require the selling and buying of timber certified by FSC or similar stringent schemes.
Methods
Data collection
We set up arrays of camera traps from 2018 to 2021 in 14 logging concessions owned by 11 different companies (5 FSC and 6 non-FSC) in Gabon and the Republic of Congo (Fig. 1). Seven FSC-certified concessions were each paired to the closest non-FSC concession that was similar in terms of terrain and forest type20. All concessions are situated in a matrix of connected forests. In each pair of concessions, camera traps (Bushnell Trophy Cam HD for pairs 1–6 and Browning 2018 Spec Ops Advantage for pair 7) were deployed simultaneously to account for seasonal differences, for 2–3 months. There was one exception where Covid restrictions obliged the cameras to remain in place for longer (Extended Data Table 1). Camera-trap grid locations in each pair of concessions were chosen on the basis of similarity between potential drivers of mammal abundance, including distance to settlements, roads, rivers, protected areas, elevation (Extended Data Fig. 2 and Extended Data Table 5) and time since logging (2–10 years before our study), although some camera grids overlapped older logging blocks. Camera traps were set out in systematic, 1 km spaced grids with a random start point. On reaching the predetermined GPS locations, the first potential installation location was used where cameras had at least 4 m of visibility. This ensured that each grid was representative of environmental heterogeneity: that is, not specifically targeting or ignoring trails or other landscape elements that could influence detection61. The 1 km intercamera distance exceeds most species’ home range sizes to avoid spatial autocorrelation. Species were not expected to migrate within the sampling duration of the study. Between 28 and 36 cameras were deployed in each concession, totalling 474 camera traps, distributed over 474 km2 (Extended Data Table 1). Cameras were installed at a height of 30 cm to enable observations of mammals of all sizes. Cameras were programmed to take bursts of three photos to maximize the chance of detection and to take a photo every 12 h for correct calculation of active days in the event of a defect before the end of the deployment period. For each camera, we recorded whether there was an elephant path, skidder trail, small wildlife trail or an absence of a trail or path, in the field of view of each camera (Extended Data Fig. 4 and Extended Data Table 5). We also visually estimated forest visibility (0–10 m, 11–20 m, greater than 20 m), slope (0–5°, 5–20°, greater than 20°), presence of fruiting trees within 30 m and presence of small water courses within 50 m (Extended Data Fig. 3 and Extended Data Table 1). When approaching each predefined camera point, we counted cartridges, snares and hunting camps from 500 m before the camera up to its location. Various field teams were employed in different concessions and hence there may be some influence of interobserver bias of hunting observations between sites.
Photo processing and data analysis
Camera-trap efforts yielded 1,278,853 photos, including 645,165 photos with animals. All photos were annotated in the program Wild.ID v.1.0.1. We identified animals up to the species level if photo quality permitted and otherwise designated the species as ‘indet’62. As reliable species identification of small mammals is difficult, they were grouped into squirrels, rats and mice and shrews. Rare observations of humans, birds, bats, reptiles and domestic dogs were excluded from the analyses.
Observations of the same species that were at least 10 min apart were considered as separate detections. We assessed the sensitivity of this threshold by calculating the number of detections for intervals of 10, 30, 60 and 1,440 min, which all yielded proportionally similar numbers of observations across body mass classes (Supplementary Table 3). When several animals were observed, the number of individuals was determined by taking the highest number of individuals in a photo within the 10 min threshold. Sampling effort was defined as the number of camera days minus downtime due to malfunctioning cameras or obstruction of vision by vegetation.
Mammal behaviour may be different in hunted concessions, as mammals may be shyer of non-natural objects such as camera traps, which could in turn negatively affect their probability of detection. If this dynamic existed, shyness was assumed to fade over time with habituation to the materials, resulting in an increase of observations over time. We tested for an interaction between certification status and the number of observations over time using a linear model with a log-transformed number of observations for the first 68 days of all deployments, as that was the shortest concession deployment period, ensuring that all concessions were equally represented. We did not find that certification status was related to a trend in observations over time (Extended Data Fig. 5). We recognize, however, that other factors may have influenced detection probability, such as movement rates, which may be affected by hunting.
For each species for each concession, we calculated encounter rate, weighted by group size, as the number of observations divided by the sampling effort and we reported all findings using the metric ‘observations per camera-trap day’. Encounter rate was calculated for all species combined, per body mass class, per IUCN Red List category63 and per taxonomic group. Body mass of each species was determined by taking the mean across sexes62. Taxonomic groups Hyracoidea and Tubulidentata were excluded from the taxonomic analysis because of low sample sizes. Shrews were included as rodents in the taxonomy analysis even though they are formally not rodents because they are difficult to distinguish from mice. We consider this acceptable given that shrews are functionally very similar to rodents in the light of this study. To study the impact of certification on total estimated faunal biomass, the encounter rate of each species was multiplied by its average body mass divided by the sampling effort.
To assess whether encounter rates varied between FSC-certified and non-FSC concessions, we quantified the means of the paired concessions using linear mixed-effects models with concession pairs, concessions and cameras as random effects, whereby cameras were nested in concessions, in concession pairs, in a multilevel random effect structure. We allowed the means of concession pairs to vary between body mass class, IUCN Red List category and taxonomic group, if supported by model selection. We tested whether potential drivers of mammal abundance (Extended Data Figs. 2, 3 and 4) were important using a model-selection approach based on minimization of Bayesian information criterion values (Supplementary Table 4). We found that the inclusion of geographic covariates did not substantially improve the model for body mass classes, taxonomic groups and IUCN categories. Only for all mammals pooled together, the inclusion of elevation and distance to rivers resulted in slightly improved models but differences were negligible and did not support strong evidence for a significant influence of these covariates64. Quadratic geographic covariate terms and camera-trap site covariates did not result in better models. Pairwise comparisons were multivariate t adjusted. We used two-sided Wilcoxon signed-rank tests for all other analyses (Extended Data Table 5). Statistical analyses were performed in R v.4.2.2.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data that support the findings of this study are available in the Zenodo repository under https://doi.org/10.5281/zenodo.10061155 (ref. 65). Source data are provided with this paper.
Code availability
R code for statistical analyses and data tables are available in the Zenodo repository under https://doi.org/10.5281/zenodo.10061155 (ref. 65).
References
Blaser, J., Sarre, A., Poore, D. & Johnson, S. Status of Tropical Forest Management 2011 (ITTO, 2011).
Romero, C. et al. Evaluation of the impacts of Forest Stewardship Council (FSC) certification of natural forest management in the tropics: a rigorous approach to assessment of a complex conservation intervention. Int. For. Rev. 19, 36–49 (2018).
van der Ven, H. & Cashore, B. Forest certification: the challenge of measuring impacts. Curr. Opin. Environ. Sustain. 32, 104–111 (2018).
Benítez-López, A. et al. The impact of hunting on tropical mammal and bird populations. Science 356, 180–183 (2017).
Kleinschroth, F., Laporte, N., Laurance, W. F., Goetz, S. J. & Ghazoul, J. Road expansion and persistence in forests of the Congo Basin. Nat. Sustain. 2, 628–634 (2019).
Leisher, C. et al. Ranking the direct threats to biodiversity in sub-Saharan Africa. Biodivers. Conserv. 31, 1329–1343 (2022).
Tilker, A. et al. Habitat degradation and indiscriminate hunting differentially impact faunal communities in the Southeast Asian tropical biodiversity hotspot. Commun. Biol. 2, 396 (2019).
Abernethy, K. A., Coad, L., Taylor, G., Lee, M. E. & Maisels, F. Extent and ecological consequences of hunting in Central African rainforests in the twenty-first century. Phil. Trans. R. Soc. B 368, 20120303 (2013).
Tritsch, I. et al. Do forest-management plans and FSC certification help avoid deforestation in the Congo Basin? Ecol. Econ. 175, 106660 (2020).
Cerutti, P. O. et al. Social Impacts of the Forest Stewardship Council Certification (Center for International Forestry Research, 2014).
Roopsind, A., Caughlin, T. T., Sambhu, H., Fragoso, J. M. V. & Putz, F. E. Logging and indigenous hunting impacts on persistence of large neotropical animals. Biotropica 49, 565–575 (2017).
Tobler, M. W. et al. Do responsibly managed logging concessions adequately protect jaguars and other large and medium-sized mammals? Two case studies from Guatemala and Peru. Biol. Conserv. 220, 245–253 (2018).
Bahaa-el-din, L. et al. Effects of human land-use on Africa’s only forest-dependent felid: the African golden cat Caracal aurata. Biol. Conserv. 199, 1–9 (2016).
Stokes, E. J. et al. Monitoring great ape and elephant abundance at large spatial scales: measuring effectiveness of a conservation landscape. PLoS ONE 5, e10294 (2010).
Polisar, J. et al. Using certified timber extraction to benefit jaguar and ecosystem conservation. Ambio 46, 588–603 (2017).
Sollmann, R. et al. Quantifying mammal biodiversity co-benefits in certified tropical forests. Divers. Distrib. 23, 317–328 (2017).
Ramage, B. S. et al. Pseudoreplication in tropical forests and the resulting effects on biodiversity conservation. Conserv. Biol. 27, 364–372 (2013).
Burivalova, Z., Hua, F., Koh, L. P., Garcia, C. & Putz, F. A critical comparison of conventional, certified and community management of tropical forests for timber in terms of environmental, economic and social variables. Conserv. Lett. 10, 4–14 (2017).
Campos-Cerqueira, M. et al. How does FSC forest certification affect the acoustically active fauna in Madre de Dios, Peru? Remote Sens. Ecol. Conserv. https://doi.org/10.1002/rse2.120 (2019).
Grantham, H. S. et al. Spatial priorities for conserving the most intact biodiverse forests within Central Africa. Environ. Res. Lett. 15, 0940b5 (2020).
Atwood, T. B. et al. Herbivores at the highest risk of extinction among mammals, birds and reptiles. Sci. Adv. 6, eabb8458 (2020).
Cardillo, M. et al. Multiple causes of high extinction risk in large mammal species. Science 309, 1239–1241 (2005).
Figel, J. J., Hambal, M., Krisna, I., Putra, R. & Yansyah, D. Malignant snare traps threaten an irreplaceable megafauna community. Trop. Conserv. Sci. 14, 1940082921989187 (2021).
Yasuoka, H. et al. Changes in the composition of hunting catches in southeastern Cameroon: a promising approach for collaborative wildlife management between ecologists and local hunters. Ecol. Soc. 20, 25 (2015).
Peres, C. A. & Dolman, P. M. Density compensation in neotropical primate communities: evidence from 56 hunted and nonhunted Amazonian forests of varying productivity. Oecologia 122, 175–189 (2000).
Maisels, F. et al. Devastating decline of forest elephants in Central Africa. PLoS ONE 8, e59469 (2013).
Wall, J. et al. Human footprint and protected areas shape elephant range across Africa. Curr. Biol. 31, 2437–2445 (2021).
Beirne, C. et al. African forest elephant movements depend on time scale and individual behavior. Sci. Rep. 11, 12634 (2021).
Challender, D. W. S., Heinrich, S., Shepherd, C. R. & Katsis, L. K. D. in Pangolins (eds Challender, D. W. S. et al.) 259–276 (Academic, 2020).
Rogers, H. S., Donoso, I., Traveset, A. & Fricke, E. C. Cascading impacts of seed disperser loss on plant communities and ecosystems. Annu. Rev. Ecol. Evol. Syst. 52, 641–666 (2021).
Bello, C. et al. Defaunation affects carbon storage in tropical forests. Sci. Adv. 1, e1501105 (2015).
Chanthorn, W. et al. Defaunation of large-bodied frugivores reduces carbon storage in a tropical forest of Southeast Asia. Sci. Rep. 9, 10015 (2019).
Berzaghi, F., Bretagnolle, F., Durand-Bessart, C. & Blake, S. Megaherbivores modify forest structure and increase carbon stocks through multiple pathways. Proc. Natl Acad. Sci. USA 120, e2201832120 (2023).
Berzaghi, F., Chami, R., Cosimano, T. & Fullenkamp, C. Financing conservation by valuing carbon services produced by wild animals. Proc. Natl Acad. Sci. USA 119, e2120426119 (2022).
Johnson, C. K. et al. Global shifts in mammalian population trends reveal key predictors of virus spillover risk. Proc. R. Soc. B 287, 20192736 (2020).
Ibbett, H. et al. Experimentally assessing the effect of search effort on snare detectability. Biol. Conserv. 247, 108581 (2020).
Wilkie, D. S., Sidle, J. G., Boundzanga, G. C., Auzel, P. & Blake, S. in The Cutting Edge (eds Fimbel, R. A. et al.) 375–400 (Columbia Univ. Press, 2001).
Bicknell, J. E., Struebig, M. J. & Davies, Z. G. Reconciling timber extraction with biodiversity conservation in tropical forests using reduced-impact logging. J. Appl. Ecol. 52, 379–388 (2015).
Laméris, D. W., Tagg, N., Kuenbou, J. K., Sterck, E. H. M. & Willie, J. Drivers affecting mammal community structure and functional diversity under varied conservation efforts in a tropical rainforest in Cameroon. Anim. Conserv. 23, 182–191 (2020).
Morgan, D. et al. African apes coexisting with logging: comparing chimpanzee (Pan troglodytes troglodytes) and gorilla (Gorilla gorilla gorilla) resource needs and responses to forestry activities. Biol. Conserv. 218, 277–286 (2018).
Sollmann, R., Mohamed, A., Samejima, H. & Wilting, A. Risky business or simple solution—relative abundance indices from camera-trapping. Biol. Conserv. 159, 405–412 (2013).
Doumenge, C., Palla, F., Madzous, I. & Ludovic, G. (eds) State of Protected Areas in Central Africa: 2020 (OFAC-COMIFAC, Yaounde, Cameroon & IUCN, 2021).
Strindberg, S. et al. Guns, germs and trees determine density and distribution of gorillas and chimpanzees in Western Equatorial Africa. Sci. Adv. 4, eaar2964 (2018).
Laurance, W. F. et al. Averting biodiversity collapse in tropical forest protected areas. Nature 489, 290–293 (2012).
Poulsen, J. R. et al. Poaching empties critical Central African wilderness of forest elephants. Curr. Biol. 27, R134–R135 (2017).
Poulain, F. et al. A camera trap survey in the community zone of Lobéké National Park (Cameroon) reveals a nearly intact mammalian community. Afr. J. Ecol. 61, 523–529 (2023).
Hedwig, D. et al. A camera trap assessment of the forest mammal community within the transitional savannah–forest mosaic of the Batéké Plateau National Park, Gabon. Afr. J. Ecol. 56, 777–790 (2018).
Bruce, T. et al. Using camera trap data to characterise terrestrial larger‐bodied mammal communities in different management sectors of the Dja Faunal Reserve, Cameroon. Afr. J. Ecol. 56, 759–776 (2018).
Breuer, T., Breuer‐Ndoundou Hockemba, M., Opepa, C. K., Yoga, S. & Mavinga, F. B. High abundance and large proportion of medium and large duikers in an intact and unhunted afrotropical protected area: insights into monitoring methods. Afr. J. Ecol. 59, 399–411 (2021).
Potapov, P. et al. The last frontiers of wilderness: tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 3, e1600821 (2017).
Gibson, L. et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478, 378–381 (2011).
Burivalova, Z., Şekercioğlu, Ç. H. & Koh, L. P. Thresholds of logging intensity to maintain tropical forest biodiversity. Curr. Biol. 24, 1893–1898 (2014).
Ward, M. et al. Just ten percent of the global terrestrial protected area network is structurally connected via intact land. Nat. Commun. 11, 4563 (2020).
Brennan, A. et al. Functional connectivity of the world’s protected areas. Science 376, 1101–1104 (2022).
Clark, C. J., Poulsen, J. R., Malonga, R. & Elkan, P. W. Logging concessions can extend the conservation estate for Central African tropical forests. Conserv. Biol. 23, 1281–1293 (2009).
Rayden, T. & Essono, R. E. Evaluation of the Management of Wildlife in the Forestry Concessions Around the National Parks of Lopé, Waka and Ivindo (WWF, 2010).
Edwards, D. P., Tobias, J. A., Sheil, D., Meijaard, E. & Laurance, W. F. Maintaining ecosystem function and services in logged tropical forests. Trends Ecol. Evol. 29, 511–520 (2014).
Nasi, R., Billand, A. & van Vliet, N. Managing for timber and biodiversity in the Congo Basin. For. Ecol. Manag. 268, 103–111 (2012).
FSC Global Strategy 2021–2026: Demonstrating the Value and Benefits of Forest Stewardship (FSC, 2020).
Salzman, J., Bennett, G., Carroll, N., Goldstein, A. & Jenkins, M. The global status and trends of payments for ecosystem services. Nat. Sustain. 1, 136–144 (2018).
Zwerts, J. A. et al. Methods for wildlife monitoring in tropical forests: comparing human observations, camera traps and passive acoustic sensors. Conserv. Sci. Pract. 3, e568 (2021).
Kingdon, J. The Kingdon Field Guide to African Mammals (Bloomsbury, 2015).
The IUCN Red List of Threatened Species Version 2022-1 (IUCN, accessed 10 August 2022).
Anderson, D. R. Model Based Inference in the Life Sciences: A Primer on Evidence Vol. 31 (Springer, 2008).
Zwerts, J. A. FSC-certified forest management benefits large mammals compared to non-FSC. Zenodo https://doi.org/10.5281/zenodo.10061155 (2023).
Acknowledgements
We thank the logging companies for access to their concessions, 263 people from WEA for fieldwork assistance and 23 students and assistants for data processing. We also thank Y. Hautier for his insights concerning the statistical analyses. The work was carried out with permission from the Gabonese Centre National de la Recherche Scientifique et Technologique (CENAREST) under research permit no. AV AR0046/18 and the Congolese Institut National de Recherche Forestière under research permit nos. 219 and 126 issued by the National Forest Research Institute (IRF) of Congo with the help of the Wildlife Conservation Society (WCS) Congo, under no. 219MRSIT/IRF/DG/DS on 17 July 2019 and the extension under no. 126MRSIT/IRF/DG/DS on 4 August 2020. J.A.Z. received support for this work from the Dutch Research Council NWO through the graduate programme Nature Conservation, Management and Restoration (grant no. 022.006.011), Programme de Promotion de l’Exploitation Certifiée des Forêts (PPECF) de la COMIFAC (à travers la KfW) under grant no. C146, WWF Netherlands, WWF Germany and the Prince Bernhard Chair for International Nature Conservation of Utrecht University.
Author information
Authors and Affiliations
Contributions
J.A.Z., M.v.K., J.v.d.W. and G.B.T. conceptualized this article. J.A.Z., E.A.M.G. and H.F.D.Z. were responsible for data curation. J.A.Z. and E.A.M.G. conducted the formal analysis. J.A.Z., M.v.K., E.A.M.G., E.H.M.S. and P.A.V. developed the methodology. J.A.Z. and H.F.D.Z. undertook investigations. J.A.Z. and E.A.M.G. created the visualizations. P.A.V., J.A.Z., M.v.K. and J.v.d.W. acquired funding. J.A.Z. and G.B.T. were responsible for project administration. J.A.Z., M.v.K., G.B.T., E.H.M.S., P.A.V. and F.M. supervised the work. J.A.Z. wrote the original draft manuscript. J.A.Z., M.v.K., J.v.d.W., F.M., G.B.T., P.A.V. and E.H.M.S. reviewed and edited the final article.
Corresponding author
Ethics declarations
Competing interests
J.A.Z. is an unpaid individual member of the FSC Environmental chamber, sub-chamber North. G.B.T. is an unpaid individual member of the FSC Environmental chamber, sub-chamber South and, since 2018, also a member of an advisory committee to the Board of Directors of FSC, the Policy and Standard Committee. J.v.d.W. and H.F.D.Z. have unpaid institutional membership of FSC through WWF International. The remaining authors declare no competing interests.
Peer review
Peer review information
Nature thanks Julia Fa, Roland Kays and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Estimated faunal biomass derived from mammal encounter rates.
(a) Estimated faunal biomass was higher (p = 0.016) in FSC-certified (n = 7) than in non-FSC concessions (n = 7). Numbers represent paired FSC-certified and non-FSC concessions linked by grey lines. Data is represented as a boxplot, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. Two-sided Wilcoxon signed-rank, *: p < 0.05. Panels (b–e) represent the contributions of different body mass classes to the estimated faunal biomass derived from mammal encounter rates in FSC-certified (n = 7) and non-FSC concessions (n = 7). (b) in kg / camera-trap day; (c) as a proportion of total faunal biomass; (d) in kg /day for species up to 100 kg; (e) as a proportion of the total faunal biomass for species up to 100 kg. FSC-certified concessions had higher overall biomass whereby mammals weighing more than 10 kg made up a larger proportion of the total biomass than in non-FSC concessions.
Extended Data Fig. 2 Geographic covariates.
(a) Distance to roads, (b) rivers, (c) human settlements, (d) and protected areas, as well as (e) elevation, did not differ significantly between camera locations in FSC-certified (n = 7) and non-FSC concessions (n = 7). Numbers represent paired FSC-certified and non-FSC concessions linked by grey lines. Data are represented as boxplots, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. Two-sided Wilcoxon signed-rank, ns: p > 0.05. Exact p-values are summarized in Extended Data Table 4.
Extended Data Fig. 3 Camera trap site covariates.
(a) The presence of fruiting trees within 30 m, (b) visibility, (c) the presence of small water courses within 50 m distance and (d) slope, expressed in proportions, did not differ significantly between camera locations in FSC-certified (n = 7) and non-FSC concessions (n = 7). Numbers represent paired FSC-certified and non-FSC concessions linked by grey lines. Data are represented as boxplots, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. Two-sided Wilcoxon signed-rank, ns: p > 0.05. Exact p-values are summarized in Extended Data Table 4.
Extended Data Fig. 4 The presence of trails or paths in the field of view of randomly placed cameras.
Each camera trap installation location was characterized as either an elephant path, skidder trail, small wildlife trail or as an absence of a trail or path. Only elephant paths, expressed in proportions, were encountered more often in FSC-certified concessions (n = 7) than in non-FSC concessions (n = 7), whereas the presence or absence of the other three types of installation locations was equivalent between the two forest management types. Camera trap sites were selected as the closest location from the predetermined GPS locations with both a suitable tree and a minimum of four metres visibility. Following this method, randomly encountering more elephant paths is in itself an indication of higher elephant abundances in FSC-certified concessions. Numbers represent paired FSC-certified and non-FSC concessions linked by grey lines. Data are represented as boxplots, where central lines represent medians and lower and upper lines correspond to the first and third quartiles, whiskers reflect 1.5 times the interquartile range. Two-sided Wilcoxon signed-rank, *: p <= 0.05, ns: p > 0.05. Exact p-values are summarized in Extended Data Table 4.
Extended Data Fig. 5 Observations over time.
This analysis explored whether variation in hunting induced mammal shyness for non-natural objects influenced detection differentially in FSC-certified (n = 7) and non-FSC concessions (n = 7). We did not find support for an effect of certification status on the number of observations over time. Linear model: p = 0.892.
Supplementary information
Supplementary Information
Supplementary Tables 1, 2 and 4.
Supplementary Table 3
Observation numbers calculated with different time thresholds between camera trap observations of the same species.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zwerts, J.A., Sterck, E.H.M., Verweij, P.A. et al. FSC-certified forest management benefits large mammals compared to non-FSC. Nature 628, 563–568 (2024). https://doi.org/10.1038/s41586-024-07257-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-024-07257-8
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
