
Abstract
In all organisms, innate immune pathways sense infection and rapidly activate potent immune responses while avoiding inappropriate activation (autoimmunity). In humans, the innate immune receptor cyclic GMP–AMP synthase (cGAS) detects viral infection to produce the nucleotide second messenger cyclic GMP–AMP (cGAMP), which initiates stimulator of interferon genes (STING)-dependent antiviral signalling1. Bacteria encode evolutionary predecessors of cGAS called cGAS/DncV-like nucleotidyltransferases2 (CD-NTases), which detect bacteriophage infection and produce diverse nucleotide second messengers3. How bacterial CD-NTase activation is controlled remains unknown. Here we show that CD-NTase-associated protein 2 (Cap2) primes bacterial CD-NTases for activation through a ubiquitin transferase-like mechanism. A cryo-electron microscopy structure of the Cap2–CD-NTase complex reveals Cap2 as an all-in-one ubiquitin transferase-like protein, with distinct domains resembling eukaryotic E1 and E2 proteins. The structure captures a reactive-intermediate state with the CD-NTase C terminus positioned in the Cap2 E1 active site and conjugated to AMP. Cap2 conjugates the CD-NTase C terminus to a target molecule that primes the CD-NTase for increased cGAMP production. We further demonstrate that a specific endopeptidase, Cap3, balances Cap2 activity by cleaving CD-NTase–target conjugates. Our data demonstrate that bacteria control immune signalling using an ancient, minimized ubiquitin transferase-like system and provide insight into the evolution of the E1 and E2 machinery across domains of life.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
For the 2:2 E. cloacae Cap2–CD-NTase complex, coordinates are available at the RCSB Protein Data Bank (PDB) (http://www.rcsb.org) under accession code 7TO3, and electron micrscopy maps are available at the Electron Microscopy Data Bank (EMDB) (https://www.ebi.ac.uk/emdb/) under accession code EMD-26028. For the 2:1 E. cloacae Cap2–CD-NTase complex, coordinates are available under PDB accession code 7TQD and electron microscopy maps are available under EMDB accession code EMD-26066. For the E. cloacae Cap2 E1 domain–CD-NTase fusion in the apo state, coordinates and crystallographic structure factors are available under PDB accession code 7TSX, and diffraction data are available at the SBGrid Data Bank (http://data.sbgrid.org) under dataset number 878. For the E. cloacae Cap2 E1 domain–CD-NTase fusion in the AMP state, coordinates and crystallographic structure factors are available under PDB accession code 7TSQ, and diffraction data are available at the SBGrid Data Bank under dataset number 877.
Change history
13 March 2023
A Correction to this paper has been published: https://doi.org/10.1038/s41586-023-05931-x
References
Ni, G., Ma, Z. & Damania, B. cGAS and STING: at the intersection of DNA and RNA virus-sensing networks. PLoS Pathog. 14, e1007148 (2018).
Whiteley, A. T. et al. Bacterial cGAS-like enzymes synthesize diverse nucleotide signals. Nature 567, 194–199 (2019).
Cohen, D. et al. Cyclic GMP-AMP signalling protects bacteria against viral infection. Nature 574, 691–695 (2019).
Ye, Q. et al. HORMA domain proteins and a Trip13-like ATPase regulate bacterial cGAS-like enzymes to mediate bacteriophage immunity. Mol. Cell 77, 709–722.e7 (2020).
Morehouse, B. R. et al. STING cyclic dinucleotide sensing originated in bacteria. Nature 586, 429–433 (2020).
Burroughs, A. M., Zhang, D., Schäffer, D. E., Iyer, L. M. & Aravind, L. Comparative genomic analyses reveal a vast, novel network of nucleotide-centric systems in biological conflicts, immunity and signaling. Nucleic Acids Res. 43, 10633–10654 (2015).
Millman, A., Melamed, S., Amitai, G. & Sorek, R. Diversity and classification of cyclic-oligonucleotide-based anti-phage signalling systems. Nat. Microbiol. 5, 1608–1615 (2020).
Severin, G. B. et al. Direct activation of a phospholipase by cyclic GMP-AMP in El Tor Vibrio cholerae. Proc. Natl Acad. Sci. USA 115, E6048–E6055 (2018).
Lowey, B. et al. CBASS immunity uses CARF-related effectors to sense 3′–5′- and 2′–5′-linked cyclic oligonucleotide signals and protect bacteria from phage infection. Cell 182, 38–49.e17 (2020).
Lau, R. K. et al. Structure and mechanism of a cyclic trinucleotide-activated bacterial endonuclease mediating bacteriophage immunity. Mol. Cell 77, 723–733.e6 (2020).
Duncan-Lowey, B., McNamara-Bordewick, N. K., Tal, N., Sorek, R. & Kranzusch, P. J. Effector-mediated membrane disruption controls cell death in CBASS antiphage defense. Mol. Cell 81, 5039–5051.e5 (2021).
Davies, B. W., Bogard, R. W., Young, T. S. & Mekalanos, J. J. Coordinated regulation of accessory genetic elements produces cyclic di-nucleotides for V. cholerae virulence. Cell 149, 358–370 (2012).
Dziejman, M. et al. Comparative genomic analysis of Vibrio cholerae: genes that correlate with cholera endemic and pandemic disease. Proc. Natl Acad. Sci. USA 99, 1556–1561 (2002).
Schulman, B. A. & Harper, J. W. Ubiquitin-like protein activation by E1 enzymes: the apex for downstream signalling pathways. Nat. Rev. Mol. Cell Biol. 10, 319–331 (2009).
Cappadocia, L. & Lima, C. D. Ubiquitin-like protein conjugation: structures, chemistry, and mechanism. Chem. Rev. 118, 889–918 (2018).
Lake, M. W., Wuebbens, M. M., Rajagopalan, K. V. & Schindelin, H. Mechanism of ubiquitin activation revealed by the structure of a bacterial MoeB–MoaD complex. Nature 414, 325–329 (2001).
Xu, X., Wang, T., Niu, Y., Liang, K. & Yang, Y. The ubiquitin-like modification by ThiS and ThiF in Escherichia coli. Int. J. Biol. Macromol. 141, 351–357 (2019).
Burroughs, A. M., Iyer, L. M. & Aravind, L. The natural history of ubiquitin and ubiquitin-related domains. Front. Biosci. Landmark Ed. 17, 1433–1460 (2012).
Nandi, D., Tahiliani, P., Kumar, A. & Chandu, D. The ubiquitin-proteasome system. J. Biosci. 31, 137–155 (2006).
Burroughs, A. M., Iyer, L. M. & Aravind, L. Natural history of the E1-like superfamily: Implication for adenylation, sulfur transfer, and ubiquitin conjugation. Proteins Struct. Funct. Bioinformatics 75, 895–910 (2009).
Lehmann, C., Begley, T. P. & Ealick, S. E. Structure of the Escherichia coli ThiS–ThiF complex, a key component of the sulfur transfer system in thiamin biosynthesis. Biochemistry 45, 11–19 (2006).
Kaiser, S. E. et al. Noncanonical E2 recruitment by the autophagy E1 revealed by Atg7–Atg3 and Atg7–Atg10 structures. Nat. Struct. Mol. Biol. 19, 1242–1249 (2012).
Yamaguchi, M. et al. Noncanonical recognition and UBL loading of distinct E2s by autophagy-essential Atg7. Nat. Struct. Mol. Biol. 19, 1250–1256 (2012).
Taherbhoy, A. M. et al. Atg8 transfer from Atg7 to Atg3: a distinctive E1–E2 architecture and mechanism in the autophagy pathway. Mol. Cell 44, 451–461 (2011).
Noda, N. N. et al. Structural basis of Atg8 activation by a homodimeric E1, Atg7. Mol. Cell 44, 462–475 (2011).
Hong, S. B. et al. Insights into noncanonical E1 enzyme activation from the structure of autophagic E1 Atg7 with Atg8. Nat. Struct. Mol. Biol. 18, 1323–1330 (2011).
Schäfer, A., Kuhn, M. & Schindelin, H. Structure of the ubiquitin-activating enzyme loaded with two ubiquitin molecules. Acta Crystallogr. D Biol. Crystallogr. 70, 1311–1320 (2014).
Olsen, S. K., Capili, A. D., Lu, X., Tan, D. S. & Lima, C. D. Active site remodelling accompanies thioester bond formation in the SUMO E1. Nature 463, 906–912 (2010).
Kranzusch, P. J. et al. Structure-guided reprogramming of human cGAS dinucleotide linkage specificity. Cell 158, 1011–1021 (2014).
Govande, A. A., Duncan-Lowey, B., Eaglesham, J. B., Whiteley, A. T. & Kranzusch, P. J. Molecular basis of CD-NTase nucleotide selection in CBASS anti-phage defense. Cell Rep. 35, 109206 (2021).
Chen, Z., Jenson, J. & Li, T. Ubiquitin-like conjugation by bacterial cgas enhances anti-phage defense. Research Square https://doi.org/10.21203/rs.3.rs-1578554/v1 (2022).
Burroughs, A. M. & Aravind, L. Identification of uncharacterized components of prokaryotic immune systems and their diverse eukaryotic reformulations. J. Bacteriol. 202, e00365–20 (2020).
Gao, L. et al. Diverse enzymatic activities mediate antiviral immunity in prokaryotes. Science 369, 1077–1084 (2020).
Doron, S. et al. Systematic discovery of antiphage defense systems in the microbial pangenome. Science 359, eaar4120 (2018).
Millman, A. et al. An expanded arsenal of immune systems that protect bacteria from phages. Cell Host Microbe 30, 1556–1569 (2022).
Rousset, F. et al. Phages and their satellites encode hotspots of antiviral systems. Cell Host Microbe 30, 740–753 (2022).
Vassallo, C.N., Doering, C.R., Littlehale, M.L. et al. A functional selection reveals previously undetected anti-phage defence systems in the E. coli pangenome. Nat. Microbiol. 7, 1568–1579 (2022).
Tal, N. et al. Cyclic CMP and cyclic UMP mediate bacterial immunity against phages. Cell 184, 5728–5739.e16 (2021).
Iyer, L. M., Burroughs, A. M. & Aravind, L. The prokaryotic antecedents of the ubiquitin-signaling system and the early evolution of ubiquitin-like beta-grasp domains. Genome Biol. 7, R60 (2006).
Aravind, L., Iyer, L. M. & Burroughs, A. M. Discovering biological conflict systems through genome analysis: evolutionary principles and biochemical novelty. Annu. Rev. Biomed. Data Sci. 5, 367–391 (2022).
Oudshoorn, D., Versteeg, G. A. & Kikkert, M. Regulation of the innate immune system by ubiquitin and ubiquitin-like modifiers. Cytokine Growth Factor Rev. 23, 273–282 (2012).
Zinngrebe, J., Montinaro, A., Peltzer, N. & Walczak, H. Ubiquitin in the immune system. EMBO Rep. 15, 28–45 (2014).
Hu, H. & Sun, S.-C. Ubiquitin signaling in immune responses. Cell Res. 26, 457–483 (2016).
Qiu, J. et al. Ubiquitination independent of E1 and E2 enzymes by bacterial effectors. Nature 533, 120–124 (2016).
Grau-Bové, X., Sebé-Pedrós, A. & Ruiz-Trillo, I. The eukaryotic ancestor had a complex ubiquitin signaling system of archaeal origin. Mol. Biol. Evol. 32, 726–739 (2015).
Hennell James, R. et al. Functional reconstruction of a eukaryotic-like E1/E2/(RING) E3 ubiquitylation cascade from an uncultured archaeon. Nat. Commun. 8, 1120 (2017).
Kibby, E. M. et al. Bacterial NLR-related proteins protect against phage. Preprint at bioRxiv https://doi.org/10.1101/2022.07.19.500537 (2022).
Guzman, L. M., Belin, D., Carson, M. J. & Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 177, 4121–4130 (1995).
de Boer, H. A., Comstock, L. J. & Vasser, M. The tac promoter: a functional hybrid derived from the trp and lac promoters. Proc. Natl Acad. Sci. USA 80, 21–25 (1983).
Kropinski, A. M., Mazzocco, A., Waddell, T. E., Lingohr, E. & Johnson, R. P. Enumeration of bacteriophages by double agar overlay plaque assay. Methods Mol. Biol. 501, 69–76 (2009).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797 (2004).
Raran-Kurussi, S., Cherry, S., Zhang, D. & Waugh, D. S. Removal of affinity tags with TEV protease. Methods Mol. Biol. 1586, 221–230 (2017).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Tan, Y. Z. et al. Addressing preferred specimen orientation in single-particle cryo-EM through tilting. Nat. Methods 14, 793–796 (2017).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021).
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D 74, 531–544 (2018).
Williams, C. J. et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 27, 293–315 (2018).
Barad, B. A. et al. EMRinger: side chain-directed model and map validation for 3D cryo-electron microscopy. Nat. Methods 12, 943–946 (2015).
Kabsch, W. XDS. Acta Crystallogr. D 66, 125–132 (2010).
Evans, P. R. & Murshudov, G. N. How good are my data and what is the resolution? Acta Crystallogr. D 69, 1204–1214 (2013).
Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D 62, 72–82 (2006).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Afonine, P. V. et al. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D 68, 352–367 (2012).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Zhou, W., Ryan, J. J. & Zhou, H. Global analyses of sumoylated proteins in Saccharomyces cerevisiae. Induction of protein sumoylation by cellular stresses. J. Biol. Chem. 279, 32262–32268 (2004).
Ma, K., Vitek, O. & Nesvizhskii, A. I. A statistical model-building perspective to identification of MS/MS spectra with PeptideProphet. BMC Bioinformatics 13, S1 (2012).
Mirdita, M. et al. ColabFold: making protein folding accessible to all. Nat. Methods 19, 679–682 (2022).
Evans, R. et al. Protein complex prediction with AlphaFold-Multimer. Preprint at bioRxiv https://doi.org/10.1101/2021.10.04.463034 (2021).
Hong, S. B., Kim, B.-W., Kim, J. H. & Song, H. K. Structure of the autophagic E2 enzyme Atg10. Acta Crystallogr. D 68, 1409–1417 (2012).
Yamada, R. et al. Cell-autonomous involvement of Mab21l1 is essential for lens placode development. Development 130, 1759–1770 (2003).
Juang, Y.-C. et al. OTUB1 co-opts Lys48-linked ubiquitin recognition to suppress E2 enzyme function. Mol. Cell 45, 384–397 (2012).
Shrestha, R. K. et al. Insights into the mechanism of deubiquitination by JAMM deubiquitinases from cocrystal structures of the enzyme with the substrate and product. Biochemistry 53, 3199–3217 (2014).
Acknowledgements
The authors thank R. Parker and A. Desai for critical reading of the manuscript; M. Herzik for advice on cryo-EM data collection and processing; C. Ebmeirer and the Mass Spectrometry Facility at the University of Colorado Boulder (RRID:SCR_018992) for assistance with sample preparation, experimental details and data analysis (NIH S10-OD025267) which was funded by the Core Voucher from the CU Boulder Financial Futures Program (A.T.W.); the CU Boulder Department of Biochemistry Shared Instruments Pool core facility (RRID:SCR_018986) and its staff; J. Kralj at CU Boulder for use of his laboratory’s plate reader; and members of the Corbett and Whiteley laboratories for advice and helpful discussion. The authors acknowledge the facilities of the cryo-EM facility at UC San Diego, and technical assistance from R. Ashley in cryo-EM sample preparation and data collection. This work was funded by NIH R35 GM144121 (K.D.C.), the NIH Office of the Director DP2 AT012346 and a Mallinckrodt Foundation Grant (A.T.W.) and NIH R01 GM116897 and S10 OD023498 (H.Z.). H.E.L. is supported as a fellow of the Jane Coffin Childs Memorial Fund for Medical Research and R.K.L. is supported by NIH F31GM137600. X-ray diffraction data were collected at the Northeastern Collaborative Access Team beamlines, which are funded by the National Institute of General Medical Sciences from the National Institutes of Health (P30 GM124165). The Eiger 16M detector on 24-ID-E is funded by a NIH-ORIP HEI grant (S10OD021527). This research used resources of the Advanced Photon Source; a US Department of Energy (DOE) Office of Science User Facility operated for the DOE Office of Science by Argonne National Laboratory under contract no. DE-AC02-06CH11357.
Author information
Authors and Affiliations
Contributions
Experiments were designed and conceived by H.E.L., Q.Y., A.T.W. and K.D.C. Phage assays were performed by H.E.L. and A.E.S. The in vivo bacterial assays, cGAMP generation assays, and bioinformatic analysis of E1, E2 and JAB domains were performed by H.E.L. Biochemical experiments were performed by Q.Y. Mass spectrometry experiments were performed by Q.Y., Y.Q., H.Z. and H.E.L. Structural experiments and analysis were performed by Q.Y., Y.G. and K.D.C. Protein purification was performed by Q.Y. and R.K.L. Bioinformatic analysis of CD-NTase and Cap2 proteins was performed by K.D.C. and H.E.L. Figures were prepared by H.E.L., A.T.W. and K.D.C. The manuscript was written by H.E.L., A.T.W. and K.D.C.. All authors contributed to editing the manuscript and support the conclusions.
Corresponding authors
Ethics declarations
Competing interests
The University of Colorado Boulder and the University of California San Diego have patents pending for Cap2 and Cap3 technologies on which H.E.L., Q.Y., Y.G., K.D.C. and A.T.W are listed as inventors.
Peer review
Peer review information
Nature thanks Ivan Dikic, Karl-Peter Hopfner and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Phage protection assays, inputs for IPs, and CD-NTase antibody verification along with CryoEM information.
(a) Image of double agar overlay phage infection assay used to measure efficiency of plating for a lysate of phage T2. E. coli MG1655 expressing the indicated vectors is shown. Zones of clearance (plaques) represent successful phage infection and replication. Apparent plaque forming units (PFU) per mL is calculated for the lysate infecting each bacterial genotype. Fold protection is the PFU per mL of empty vector divided by Vc CBASS, ~104 in this assay. (b) Efficiency of plating of the indicated phage when infecting E. coli expressing CBASS with the indicated genotype. Data plotted as in Fig. 1b. C.D. CD-NTase: DID131AIA.; C.D. capV: C62A. (c) Efficiency of plating of the indicated phage when infecting E. coli expressing V. cholerae CBASS with the indicated genotypes. Data plotted as in Fig. 1b. (d) Western blot analysis of cell lysates (inputs) and αVSV-G immunoprecipitation of E. coli expressing CBASS with the indicated genotypes. These samples correspond to the mass spectrometry in Fig. 1c. αRNAP western blot serves as a loading control for bacterial cells. (-): CBASS operon, CD-NTase without VSV-G; (+): CBASS operon, CD-NTase with N-terminal VSV-G. (e) Whole cell western blot analysis of E. coli expressing either an empty vector (EV) or CBASS (wild-type). αCD-NTase Western blot used a custom CD-NTase antibody; arrow indicates monomeric CD-NTase at the expected molecular weight. αRNAP western blot serves as a loading control for bacterial cells. (f) Efficiency of plating of the indicated phage when infecting E. coli expressing CBASS with the indicated genotypes. Data plotted as in Fig. 1b. (g) Whole cell western blot analysis of E. coli expressing the indicated genotypes of CBASS. Data are the input for the immunoprecipitation presented in Fig. 1d. (h) Whole cell western blot analysis of E. coli expressing the indicated genotypes of CBASS. Data are the input for the immunoprecipitation presented in Fig. 1e. For (f),(g) and (h) ±ϕ indicates phage T2 at an MOI of 2. (i) Operon structure of CBASS from E. cloacae. See Supplementary Table 5 for relevant accession numbers. (j) Size exclusion chromatography elution profile (Superdex 200 Increase 10/300 GL) and SDS-PAGE analysis of E. cloacae Cap2–CD-NTase. The fraction used for cryoEM analysis is shaded in gray. C.D. Cap2: C109A/C548A. (k) Representative electron micrograph of E. cloacae Cap2–CD-NTase. (l) Fourier Shell Correlation (FSC) curve for the final refinement of the 2:2 Cap2–CD-NTase complex. (m) 3D FSC analysis55 for the 2:2 Cap2–CD-NTase complex. (n) Fourier Shell Correlation (FSC) curve for the final refinement of the 2:1 Cap2–CD-NTase complex. (o) 3D FSC analysis for the 2:1 Cap2–CD-NTase complex. (p) Local resolution of the final refined map for the 2:2 Cap2–CD-NTase complex, colored from blue (≤1.8 Å) to magenta (≥4.00 Å). (q) Local resolution of the final refined map for the 2:1 Cap2–CD-NTase complex colored from blue (≤1.8 Å) to magenta (≥4.00 Å). Outline indicates the areas of missing density compared to the 2:2 Cap2–CD-NTase complex.
Extended Data Fig. 2 The CD-NTase rigidifies Cap2 through a bipartite interaction and crystal structures of an E. cloacae Cap2 E1-CD-NTase fusion.
(a) Closeup view of the interaction between Cap2 (yellow/blue) and CD-NTase (orange), with key residues shown as sticks and labeled. (b) Coomassie stained SDS-PAGE of proteins that purified by Ni2+-affinity chromatography from E. coli co-expressing E. cloacae 6×His-tagged cd-ntase (His6-CD-NTase) and catalytically inactivated cap2 (C548A/C109A; C.D.) with the indicated genotype. (c) CryoEM density for 2:1 Cap2–CD-NTase complex, with domains labeled and colored as in Fig. 2b. Outline indicates the areas of missing density compared to the 2:2 Cap2–CD-NTase complex, including one protomer of CD-NTase and the E2 and linker domains for the (a) protomer of Cap2. (d) Two views of the 2:1 Cap2–CD-NTase complex, with domains labeled and colored as in panel (c). Outlines indicate the areas of missing density compared to the 2:2 Cap2–CD-NTase complex. (e) Design of a fusion between the C-terminal E1 domain of E. cloacae Cap2 (residues 363-600) and the C-terminus of CD-NTase (residues 370-381), with a three-residue GSG linker. (f) 2.1 Å resolution crystal structure of the E. cloacae Cap2–CD-NTase fusion crystallized in the presence of ATP, with two Cap2 E1 domains colored yellow and gray, and the two CD-NTase C-termini colored orange. See also Extended Data Table 2. (g) Closeup of the Cap2 adenylation active site, showing the CD-NTase–AMP conjugate and active site residues. Residues 533–546 are disordered and represented by a dotted line. Bound Mg2+ ion is shown in black. (h) View as in (g), with 2FO-FC electron density contoured at 1.0 σ around the CD-NTase–AMP conjugate and active site residues. (i) View as in (g), with FO-FC omit map density contoured at 1.5 σ around the CD-NTase–AMP conjugate. (j) Closeup of the Cap2 adenylation active site in a 1.8 Å-resolution structure of the Cap2-CD-NTase fusion crystallized in the absence of added nucleotide (apo state). (k) View as in (j), with 2Fo-Fc electron density contoured at 1.0 σ around the CD-NTase C-terminus.
Extended Data Fig. 3 Cap2 is related to autophagy E1 and E2 proteins.
(a) Protein alignment of Cap2 from E. cloacae, Cap2 from V. cholerae, ATG10 from S. cerevisiae (4EBR72), and ATG7 from S. cerevisiae (3T7H24). Domains are indicated above the alignment with colors corresponding to Fig. 2. The secondary structure of Cap2 from E. cloacae is indicated in purple with alpha helices depicted as cylinders and beta sheets as arrows. The catalytic cysteines found in the E2 and E1 domains are highlighted in red. See Supplementary Table 5 for relevant accession numbers. (b) Domain schematic of E. cloacae Cap2 and S. cerevisiae ATG7, with approximate root-mean-squared distance (Cα r.m.s.d.) values for the linker/NTD and E1 domains noted. (c) Structures of E. cloacae Cap2 (left), compared to S. cerevisiae ATG7 (right; PDB ID 4GSK22), with one protomer colored as in panel (a) and the dimer mate colored gray. For each protein, the E1 active-site cysteine residue (C548 for Cap2, C507 for ATG7) is shown as a sphere and labeled. (d) Structures of the E. cloacae Cap2 linker domain (left), compared to the S. cerevisiae ATG7 NTD (right; PDB ID 4GSK22). ATG7 features a second subdomain (residues 147-268, shown in white) inserted into the loop separating β-strands 6 and 7 (labeled) where Cap2 has a partially disordered loop (residues 319–356). (e) Structure of the Cap2 E2 domain (active-site C109 shown as a sphere), compared to Kluyveromyces marxianus ATG10 (PDB ID 3VX723), S. cerevisiae ATG3 (PDB ID 2DYT73), and Homo sapiens UBE2D2 (PDB ID 4DDG74). Structural features not shared are shown in white. The active-site cysteine of each protein is shown as a sphere.
Extended Data Fig. 4 Analysis of Cap2 mutants and epitope-tagged CD-NTase and evidence that Cap2 conjugates the CD- NTase C-terminus to a target.
(a) Efficiency of plating of the indicated phage when infecting E. coli expressing CBASS with the indicated genotype. Data plotted as in Fig. 1b. (b) Efficiency of plating of the indicated phage when infecting E. coli expressing CBASS with the indicated genotype. Data plotted as in Fig. 1b. (c) Western blot analysis of cell lysates from E. coli expressing CBASS with the indicated genotypes demonstrating that the mutations do not affect expression levels. (d) Western blot analysis of cell lysates from E. coli expressing CBASS with the indicated genotypes demonstrating that the mutations do not affect protein expression levels. (e) SDS-PAGE analysis of E. cloacae Cap2 activity assay. The indicated genotypes of His6-Cap2 and the CD-NTase were expressed from a single plasmid and the formation of a CD-NTase–His6-Cap2 conjugate was used as an indicator of Cap2 activity. (-): no CD-NTase; (+): wild-type CD-NTase; (∆C): CD-NTase lacking its C-terminal 19 residues; C.D. Cap2: C548A/C109A. Blue asterisk indicates a putative intermediate with CD-NTase thioester-linked to the Cap2 E1 catalytic cysteine (C548). The formation of a CD-NTase–Cap2 conjugate in the absence of a functional E1 catalytic cysteine (C548A) indicates that in vitro, this residue is dispensable for catalysis and the nearby E2 catalytic cysteine (C109) can function in instead. (f) Cap2 E1 active site (yellow) in Cap2–CD-NTase cryoEM structure with the residues mutated in (a) indicated and the E1 active-site cysteine residue (C548 for Cap2) shown as a sphere and labeled. The CD-NTase C-terminus (orange) conjugated to AMP (black). (g) Left: SDS-PAGE analysis of Ni2+-purified E. cloacae His6-Cap2, expressed either alone or with full-length CD-NTase. Right: Protease treatment (TEV or Cap3) of the CD-NTase–His6-Cap2 conjugate. (h) Schematic of the inferred CD-NTase–His6-Cap2 conjugate formed upon coexpression of E. cloacae His6-Cap2 and CD-NTase, with cleavage sites for Cap3 and TEV protease indicated. (i) SDS-PAGE analysis with detection by coomassie (left) or αHA western blot (right) of αHA immunoprecipitated E. cloacae Cap2 coexpressed with HA-CD-NTase. C.D. Cap2: C548A/C109A. Red asterisk indicates band used for tryptic mass spectrometry analysis in (f-g). (j) Peptides detected in tryptic mass spectrometry of the marked band in (e), showing conjugation of CD-NTase to the N-terminus of a second HA-CD-NTase molecule. See Supplementary Table 7 for mass spectrometry data. (k) Collision-induced dissociation mass spectrum of the peptide indicated in (f), with b1 peak indicated (mass of 350.1533 is that of Met+(H+)+(Phe-Ala)). (l) SDS-PAGE analysis of Ni2+-purified E. cloacae His6-Cap2 with CD-NTase with the indicated genotype.
Extended Data Fig. 5 The C-terminus of the CD-NTase is conserved in type II CBASS systems and the quantification of CD-NTase-mediated second messenger generation.
(a) Sequence logos of C-terminal 10 residues of the CD-NTase in type I (2284 sequences), type II (1556 sequences), and type II (short) (593 sequences) CBASS systems7. Type II (short) CBASS systems encode an E2 ubiquitin transferase-like enzyme without a linked E1 domain, and do not encode a JAB isopeptidase. (b) Phylogenetic tree adapted from Whiteley et al.2, with sequence logos of the C-terminus for CD-NTase clades analyzed in Cap3 experiments (Fig. 4d, Extended Data Fig. 6d–g) shown. Saturated colors bordered with solid lines depict branches of the tree that contain type II systems, whereas the de-saturated colors bordered with dashed lines depict clades with non-type II systems. The CD-NTases used in this study are listed below each sequence logo. Blue circles with numbers represent CD-NTase numbers as reported previously2. (c) cGAMP generated by αVSV-G immunoprecipitation from E. coli expressing CBASS operons with the indicated genotypes. Western blots of input are in (e) and immunoprecipitation in (f). N = 3 technical replicates representative of three independent biological replicates. Data is presented as the mean values ± the SEM. ϕ (-): no infection; ϕ (+): phage T2 at an MOI of 2; CapV (+): wild-type; CapV (C.D.): S62A; CD-NTase (+): wild-type; CD-NTase (C.D.): DID131AIA; CD-NTase (V): N-terminal VSV-G epitope tagged CD-NTase; Cap3 (+): wild-type; Cap3 (∆): genetically deleted cap3; Cap2 (+): wild-type; Cap2 (F): C-terminal 3x-FLAG epitope tagged Cap2; Cap2 (E1): C522A; Cap2 (E2): C90A. (d) Data in (c) presented on a log10 scale. (e) Whole cell western blot analysis of E. coli expressing CBASS with the indicated genotype. Data corresponds to the input for the immunoprecipitation shown in (a) and (b). (f) Western blot analysis of αVSV-G immunoprecipitation of E. coli expressing CBASS with the indicated genotypes. These data correspond to the samples used to measure cGAMP synthesis in (c) and (d). (g) Quantification of the cGAMP produced by the V. cholerae CD-NTase with, or without, a C-terminal GFP fusion. (h) Quantification of the cAAG produced by the E. cloacae CD-NTase with, or without, a C-terminal GFP fusion. For (g) and (h) N = 3 independent biological replicates and the data is presented as the mean ± the SD.
Extended Data Fig. 6 Cap3 overexpression inhibits phage protection by cognate CBASS.
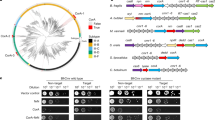
(a) Efficiency of plating of the indicated phage when infecting E. coli expressing CBASS with the indicated genotype. cd-ntase-VSV-G (-): wild-type CD-NTase; cd-ntase-VSV-G (+): C-terminal VSV-G epitope tagged CD-NTase; wild-type indicates otherwise a full CBASS operon; ∆cap3: CBASS operon with only cap3 deletion. Data plotted as in Fig. 1b. (b) Western blot analysis of cell lysates from E. coli expressing CBASS with the indicated genotypes, abbreviated as in (a). Cells were infected with phage T5 at an MOI of 2 for the indicated time prior to harvesting for analysis. (c) Efficiency of plating of the indicated phage when infecting E. coli expressing CBASS ∆cap3 in the absence or presence of overexpressed cap3 with the indicated genotype. Data plotted as in Fig. 4a. C.D. cap3: HTH101ATA. A Two-sided Student’s t-test was used to calculate significance; n.s., p > 0.05; *, p < 0.05; **, p < 0.001. See Extended Data Fig. 6h for protein alignment. (d) Efficiency of plating of the indicated phage when infecting E. coli expressing a full CBASS operon from V. cholerae in the absence or presence of overexpressed cap3 from another CBASS system, indicated on the x-axis. Data plotted as in Fig. 4a. (e) Efficiency of plating of the indicated phage when infecting E. coli expressing a full CBASS operon from E. cloacae in the absence or presence of overexpressed cap3 from another CBASS system, indicated on the x-axis. Data plotted as in Fig. 4a. (f) Efficiency of plating of the indicated phage when infecting E. coli expressing a full CBASS operon from C. freundii in the absence or presence of overexpressed cap3 from another CBASS system, indicated on the x-axis. Data plotted as in Fig. 4a. (g) Efficiency of plating of the indicated phage when infecting E. coli expressing a full CBASS operon from E. coli in the absence or presence of overexpressed cap3 from another CBASS system, indicated on the x-axis. Data plotted as in Fig. 4a. For (d-g) the red dashed boxes indicated the data utilized in Fig. 4d, See Supplementary Table 5 for relevant accession numbers. (h) Protein alignment of the JAMM/JAB protease Sst2 from S. pombe55 (Uniprot ID Q9P371; residues 235–435), Cap3 from E. cloacae, and Cap3 from V. cholerae. The active site glutamate, as well as two zinc-coordinating histidine residues, are noted. For experiments using Cap3 from E. cloacae, the first 16 annotated amino acids (green box) were removed as we found the translation start site is likely misannotated for this gene. See Supplementary Table 5 for relevant accession numbers.
Extended Data Fig. 7 Cap3 cleavage of a CD-NTase model substrate.
(a) Domain schematic and predicted structure/model of the V. cholerae Cap3-CD-NTase complex56 with the CD-NTase C-terminus and Zn2+ ion manually modeled from an overlay with a structure of S. pombe Sst2 bound to ubiquitin75 (PDB ID 4K1R). (b) Summary of tryptic digest mass spectrometry analysis of the V. cholerae Cap3-treated CD-NTase bands as in Fig. 4b. Pink arrow indicates the inferred Cap3 cleavage site; gray arrows indicate trypsin cleavage sites. See Supplementary Table 3 for data. (c) Coomassie stained SDS-PAGE of a V. cholerae model substrate (CD-NTase-GFP fusion protein) with the indicated mutations in the CD-NTase C-terminus, with and without incubation with V. cholerae Cap3. (d) Domain schematic and predicted structure/model of the E. cloacae Cap3-CD-NTase complex56 with the CD-NTase C-terminus and Zn2+ ion manually modeled from an overlay with a structure of S. pombe Sst2 bound to ubiquitin75 (PDB ID 4K1R). (e) Coomassie stained SDS-PAGE of an E. cloacae model substrate (CD-NTase-GFP fusion protein) with the indicated mutations in the CD-NTase C-terminus, with and without incubation with E. cloacae Cap3. (f) Coomassie stained SDS-PAGE of an E. cloacae model substrate (CD-NTase-GFP fusion protein) incubated with E. cloacae Cap3 with the indicated reaction condition/genotype. (g) Summary of tryptic digest mass spectrometry analysis of the E. cloacae Cap3-treated CD-NTase bands as in (f), showing the putative Cap3 cleavage site. Pink arrow indicates the inferred Cap3 cleavage site; gray arrows indicate trypsin cleavage sites. See Supplementary Table 3 for data.
Extended Data Fig. 8 CD-NTase immunoprecipitation reveals numerous potential protein targets and Cap2 homologs are found in other bacteria.
(a) Western blot analysis of cell lysates generated from E. coli expressing CBASS ∆cap3 with the additional indicated genotypes. (b) Western blot analysis of αVSV-G immunoprecipitations generated from E. coli expressing CBASS ∆cap3 with the additional indicated genotypes. These samples were used in the mass spectrometry analysis displayed in (c-f). (c) Mass spectrometry of immunoprecipitated VSV-G-CD-NTase as shown in (b). Data are label free quantitation (LFQ) score and fold enrichment comparing immunoprecipitations from bacteria expressing CBASS ∆cap3 to a strain expressing CBASS ∆cap3 cap2C522A(E1). Both strains encode an N-terminally VSV-G tagged CD-NTase. Cap2 and CD-NTase are represented as colored circles corresponding to Fig. 1a and are labeled. Proteins which we determined were significantly enriched (LFQ > 108 and a fold enrichment >4) are colored in pink. Circles above the dotted line are proteins with peptides identified only in the sample listed on the x-axis. See Supplementary Table 8 for data. (d) Characterization of the predicted functions of the proteins that were significantly enriched in (c). (e) Mass spectrometry of immunoprecipitated VSV-G-CD-NTase as in (c) comparing immunoprecipitations from bacteria expressing CBASS ∆cap3 where CD-NTase has an N-terminal VSV-G tag to a strain expressing CBASS lacking a VSV-G tag (negative control). (f) Mass spectrometry of immunoprecipitated VSV-G-CD-NTase as in (c) comparing immunoprecipitations from bacteria expressing CBASS ∆cap3 cap2C522A(E1) where CD-NTase has an N-terminal VSV-G tag to a strain expressing CBASS lacking a VSV-G tag (negative control). (g) Top: Structural comparison between E. cloacae Cap2 and a predicted structure/model of Azohydromonas australica Pap2. Cα r.m.s.d. values are reported for superposition of individual domains: E2 domain (52 Cα atoms overlaid), linker (90 Cα atoms), and E1 (133 Cα atoms). Predicted catalytic cysteine residues are noted for each protein. Bottom: Structural prediction of a Pap3 (pink) and the C-terminus of PycC (yellow) from A. australica. Predicted active site residues of Pap3 are shown as sticks with a zinc ion (gray) modeled from a structure of S. pombe Sst2 bound to ubiquitin (PDB ID 4K1R). (h) Structural comparison between E. cloacae Cap2 and a predicted structure/model of the Cap2-like protein from the Xanthamonas arboricola MBL-group operon. Cα r.m.s.d. values are reported for superposition of individual domains: E2 domain (94 Cα atoms overlaid), linker (50 Cα atoms), and E1 (154 Cα atoms). Predicted catalytic cysteine residues are noted for each protein. (i) Predicted structure/model of a complex between X. arboricola JAB domain (pink) and the C-terminus of MBL (yellow). Predicted active site residues of the JAB domain are shown in sticks, with a zinc ion (gray) modeled from a structure of S. pombe Sst2 bound to ubiquitin75 (PDB ID 4K1R). The conserved glycine residue of MBL (white) is positioned for cleavage. (j) Sequence logo for the C-terminal 9 residues of 268 MBL encoded within MBL-group operons. See also Supplementary Table 9. (k) Operon structure of previously described and proposed phage defence systems that contain E1, E2 and JAB domain containing proteins along with operons of unknown function that contain these domains. Operons are grouped by conserved protein domains. The E1-superfamily these groups is also indicated in paratheses20,39. Genes are colored by domain type; E1 and E2 domains, blue; JAB domains, purple; all other domains, grey. Metallo-β-lactamase (MBL); metal binding domain (CEHH); tandem β-grasp fold domain containing protein (multi-ub); single β-grasp fold domain containing protein (ub); Domains of unknown function (DUF); genes with no discernable domains (?). See also Supplementary Table 9.
Supplementary information
Supplementary Discussion
A detailed discussion of the evolutionary relationship between Cap2, and proteins involved in autophagy.
Supplementary Figure 1
Uncut gels. Unedited images of immunoblots and coomassie analysis corresponding to the indicated figures.
Supplementary Figure 2
Cryo-EM workflow. Workflow for structure determination in cryoSPARC. Templates used for particle picking were chosen from initial 2D classes from blob-picked particles in a 200-image subset.
Supplementary Table 1
Peptides identified in CD-NTase immunoprecipitation
Supplementary Table 2
Structural features of bacterial CD-NTase proteins
Supplementary Table 3
Trypsin Mass Spectrometry of Cap3-treated CD-NTase–GFP
Supplementary Table 4
E. coli strains and plasmids used in this study
Supplementary Table 5
Protein sequences
Supplementary Table 6
Phages used in this study
Supplementary Table 7
Trypsin Mass Spectrometry of Cap2-CD-NTase
Supplementary Table 8
Peptides identified in CD-NTase immunoprecipitation in ∆cap3 background
Supplementary Table 9
Bioinformatic analysis of other E1, E2 and JAB containing operons
Supplementary Table 10
CD-NTase phylogenetic information
Supplementary Data 1
Raw data for phage protection assays including statistics
Supplementary Data 2
Raw data for cGAMP ELISA assays
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ledvina, H.E., Ye, Q., Gu, Y. et al. An E1–E2 fusion protein primes antiviral immune signalling in bacteria. Nature 616, 319–325 (2023). https://doi.org/10.1038/s41586-022-05647-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05647-4
This article is cited by
-
Reversible conjugation of a CBASS nucleotide cyclase regulates bacterial immune response to phage infection
Nature Microbiology (2024)
-
Phage defence system CBASS is regulated by a prokaryotic E2 enzyme that imitates the ubiquitin pathway
Nature Microbiology (2024)
-
Conservation and similarity of bacterial and eukaryotic innate immunity
Nature Reviews Microbiology (2024)
-
The Vibrio cholerae CBASS phage defence system modulates resistance and killing by antifolate antibiotics
Nature Microbiology (2024)
-
Unexpected bonds: ubiquitin-like conjugation of cGAS/CD-NTases supports their enzymatic activity and antiphage defense
Signal Transduction and Targeted Therapy (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
