
Abstract
Living amphibians (Lissamphibia) include frogs and salamanders (Batrachia) and the limbless worm-like caecilians (Gymnophiona). The estimated Palaeozoic era gymnophionan–batrachian molecular divergence1 suggests a major gap in the record of crown lissamphibians prior to their earliest fossil occurrences in the Triassic period2,3,4,5,6. Recent studies find a monophyletic Batrachia within dissorophoid temnospondyls7,8,9,10, but the absence of pre-Jurassic period caecilian fossils11,12 has made their relationships to batrachians and affinities to Palaeozoic tetrapods controversial1,8,13,14. Here we report the geologically oldest stem caecilian—a crown lissamphibian from the Late Triassic epoch of Arizona, USA—extending the caecilian record by around 35 million years. These fossils illuminate the tempo and mode of early caecilian morphological and functional evolution, demonstrating a delayed acquisition of musculoskeletal features associated with fossoriality in living caecilians, including the dual jaw closure mechanism15,16, reduced orbits17 and the tentacular organ18. The provenance of these fossils suggests a Pangaean equatorial origin for caecilians, implying that living caecilian biogeography reflects conserved aspects of caecilian function and physiology19, in combination with vicariance patterns driven by plate tectonics20. These fossils reveal a combination of features that is unique to caecilians alongside features that are shared with batrachian and dissorophoid temnospondyls, providing new and compelling evidence supporting a single origin of living amphibians within dissorophoid temnospondyls.
Similar content being viewed by others

Main
Of the nine tetrapod lineages surviving from the Triassic to the present day21, caecilians have the most depauperate fossil record, with only 11 total occurrences22; of these, only Rubricacaecilia monbaroni23 and Eocaecilia micropodia11,12 represent unambiguous stem caecilians. The estimated Permo–Carboniferous origin of caecilians leaves a gap exceeding 70 million years between putative Palaeozoic relatives and Eocaecilia1. The absence of a pre-Jurassic caecilian record provides little evidence informing the pattern of morphological transformations leading to the specialized caecilian body plan, the timing and pattern of caecilian origins and diversification, the functional and ecological origins of extant caecilians, and caecilian palaeobiogeography. Furthermore, this gap has resulted in longstanding disagreement regarding the relationships of living amphibian groups to each other and to other tetrapods with multiple mutually exclusive hypotheses proposed8,14. With the discovery of Gerobatrachus hottoni7, an early Permian dissorophoid bearing a combination of batrachian and amphibamiform features, the monophyly of Batrachia nested within amphibamiform dissorophoids reached near-consensus opinion8, demonstrating the crucial nature of new fossil evidence to questions of lissamphibian origins. Despite the improved understanding of batrachian origins, the origins of Lissamphibia remain contentious, now hinging on the relationships of caecilians to batrachians and Palaeozoic tetrapods1,8,13,14. Therefore, consensus on lissamphibian origins can be resolved only with the addition of new caecilian fossils filling the morphological gap between Eocaecilia and Palaeozoic tetrapods.
Here we approach such consensus by reporting the discovery of a new stem caecilian from a multitaxic microvertebrate and macrovertebrate bonebed in the Upper Triassic Chinle Formation of Petrified Forest National Park (PEFO), Arizona, USA (Extended Data Figs. 1 and 2). This material represents the most abundant caecilian-bearing fossil locality known, with at least 76 individuals consisting of isolated three-dimensional skeletal elements that we infer to belong to the same taxon, including elements from the upper and lower jaws, and postcrania (Supplementary Information, section 1).
Systematic palaeontology
Lissamphibia Haeckel, 1866
Gymnophionomorpha Marjanović and Laurin, 2008
Funcusvermis gilmorei gen. et sp. nov.
Etymology. Funcus, Latinized form of the English word funky (funk is an upbeat, rhythmic form of dance music); vermis, worm (Latin); in honour of the 1972 song Funky Worm from the album Pleasure by the Ohio Players. The species name honours N. Gilmore, collections manager at the Academy of Natural Sciences of Drexel University in Philadelphia, PA, USA.
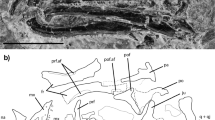
Holotype. PEFO 43891, right pseudodentary (Fig. 1 and Extended Data Figs. 3 and 4), accessioned at Petrified Forest National Park, Arizona, USA.

a–c, Composite reconstruction of craniomandibular elements in lateral (a), medial (b) and dorsal (c) views. d,e, Holotype right pseudodentary (PEFO 43891) in medial and ventral views. f, Paratype right pseudodentary (PEFO 46284) in medial view. g–i, Referred left maxillopalatine (PEFO 46481) in medial (g), ventral (h) and dorsal (i) views. j,k, Referred left pseudoangular (PEFO 46480) in medial and lateral views. l–o, Paratype right pseudodentary (PEFO 45800) in medial (l; expanded view in m) and dorsal (n; expanded view in o) views. abcnV, alveolar branch cranial nerve V; adtr, adsymphyseal tooth row; af, adductor fossa; att, attachment tissue; bp, basal pore; cnV, cranial nerve V insertions; cp, coronoid process; dpaf, dorsal pseudoangular facet; dpdf, dorsal pseudodentary facet; dtr, dentary tooth row; dz, dividing zone; ebcnV, external branch cranial nerve V; fr, facial ramus; hp, hamate process; imf, intramandibular foramen; jas, jaw articulation surface; lcm, lateral choanal margin; lecnV, lateral exit cranial nerve V; mtr, maxillary tooth row; om, orbital margin; pap, posterior pseudoangular process; pc, pulp cavity; pd, pedicel; pgp, preglenoid process; ptr, palatal tooth row; rtl, replacement tooth locus; sf, symphyseal foramen; sp, symphyseal prongs; vpaf, ventral pseudoangular facet; vpdf, ventral pseudodentary facet. Arrows indicate anterior direction.
Paratypes. PEFO 44432, PEFO 45800 and PEFO 46284 (all right pseudodentaries; Fig. 1 and Extended Data Figs. 3 and 4). Additional paratypes are listed in Supplementary Information, section 1.
Referred material. PEFO 46481, left maxillopalatine (Fig. 1 and Extended Data Fig. 4); PEFO 46480, left pseudoangular (Fig. 1 and Extended Data Fig. 3); PEFO 45810 (postatlantal vertebra), PEFO 43811 (right femur) (Extended Data Fig. 3). Additional referred specimens are listed in Supplementary Information, section 1.
Type locality and horizon. PFV 456, Thunderstorm Ridge, PEFO, Arizona, USA (Extended Data Fig. 2), within the upper Blue Mesa Member, Chinle Formation (Late Triassic: Norian); 223.036 ± 0.059 Ma (ref. 24 to 218.08 ± 0.037 Ma (ref. 25), or ~221 Ma (ref. 26); Adamanian estimated holochron27).
Diagnosis. A gymnophionomorph diagnosed by the following unique combination of features found in the holotype and paratype and referred specimens (asterisk denotes autapomorphies): symphyseal foramen* and notch subdividing the mandibular symphysis into medial and lateral processes*; at least 50 and at least 22 tooth pedicels in the dentary and adsymphyseal tooth rows, respectively. Further diagnosed by features found in referred specimens: co-ossified maxilla and palatine (compound maxillopalatine); palatal dentition of maxillopalatine terminated anteriorly by the lateral choanal margin*; maxillopalatine without osteological correlate of the tentacular organ*; absence of internal and retroarticular processes of the pseudoangular*; jaw articulation surface of pseudoangular formed by a subcircular flat pad; pseudoangular bearing a dorsally exposed adductor chamber occupying more than 30% of pseudoangular length*; three cranial nerve V insertions in pseudoangular*; femur present. Differential diagnosis in Supplementary Information, section 2.
Phylogenetic relationships
We tested the relationships of Funcusvermis gilmorei in a modified dataset6 of 63 terminal taxa including stem tetrapods, stem and crown amniotes, and temnospondyl amphibians including stereospondyls and lissamphibians (Methods). Using both maximum parsimony and Bayesian inference optimality criteria (Methods), our phylogenetic analyses robustly support Funcusvermis as the earliest-diverging gymnophionomorph, sister taxon to the clade including Eocaecilia, Rubricacaecilia and Gymnophiona. All analyses unambiguously recovered a monophyletic Lissamphibia nested within amphibamiform dissorophoids, with Gerobatrachus and Doleserpeton annectens as successive outgroups to Lissamphibia (Fig. 3 and Extended Data Figs. 5–7). Our parsimony analysis recovered Lissamphibia consisting of a polytomous trichotomy of Gymnophionomorpha, Batrachia and Albanerpetontidae (Extended Data Fig. 5), whereas our Bayesian analysis recovered Lissamphibia consisting of a sister group relationship between Batrachia and a clade comprised of a sister group relationship between Gymnophionomorpha and Albanerpetontidae (Extended Data Fig. 7). The varying position of albanerpetontids in these and other recent analyses28 highlights the ghost lineage from 150 million years ago (Ma) preceding their earliest occurrences in the Middle Jurassic epoch29 as an outstanding gap obscuring conclusive resolution of relationships amongst major lissamphibian lineages. These results suggest that the caecilian-like anatomy in Chinlestegophis jenkinsi (a Late Triassic diminutive burrowing stereospondyl also found in the Chinle Formation14) is convergent with that of gymnophionomorphs such as Eocaecilia because of adaptations facilitating fossoriality (further discussed in Supplementary Information, section 3 and Extended Data Figs. 8–10).
Origins of the lissamphibian jaw apparatus
Funcusvermis indicates that many features of the lissamphibian mandibular ramus appeared initially in amphibamiform dissorophoids and were later lost or modified in batrachians, albanerpetontids and gymnophionomorphs (Fig. 3). All dentition in Funcusvermis (Fig. 1) is pedicellate—the oldest known example of this distinctive tooth form in crown Lissamphibia—reinforcing hypotheses that pedicellate teeth are derived in amphibamiform dissorophoids10, conserved in gymnophionomorphs and batrachians30, and lost in albanerpetontids29. The rod-like pseudodentary of Funcusvermis resembles that of Eocaecilia12 and the dentary of Doleserpeton9 in the presence of tightly packed homodont tooth pedicels in parallel labial (dentary) and lingual (adsymphyseal) rows. The symphyseal foramen, of similar form and position to those of albanerpetontids31, suggests that the Meckel’s cartilage never ossified at the mandibular symphysis, probably a conservation of the ancestral condition of temnospondyls32, and differing from the ossified condition of this element that forms a closed mandibular symphysis in batrachians and other gymnophionomorphs. As in Doleserpeton9, a vertical notch bisects the mandibular symphysis between the anterior termini of the dentary and adsymphyseal tooth rows forming medial and lateral processes in Funcusvermis (Fig. 1 and Extended Data Fig. 3); these are similar to the more pronounced symphyseal prongs of albanerpetontids (for example, in refs. 28,29,31), indicating that this feature may be ancestral to Lissamphibia and later lost in Batrachia and the common ancestor of Eocaecilia and Gymnophiona.
In Funcusvermis, the presence of 22 or more teeth in the adsymphyseal tooth row is similar to the more than 20 teeth reported in Eocaecilia12, suggesting a transformation of the condition exhibited in Doleserpeton (5–7 teeth9) through distal expansion via addition of new teeth. In living caecilians, the lingual tooth row forms embryonically on a distinct anlage that later ossifies to the medial surface of the dentary forming the medial part of the mandibular symphysis and the lingual tooth row of adult caecilians33. Recent identifications of a dorsally facing tooth-bearing adsymphyseal (equivalent to the parasymphyseal (plate)) medial to (and separate from) the dentary at the mandibular symphysis in early branching tetrapods34,35, juvenile temnospondyls36 and dissorophoid temnospondyls37 suggests that in taxa that appear to bear a lingual tooth row at the mandibular symphysis of the ‘dentary’ (for example, Doleserpeton, Funcusvermis and other gymnophionomorphs), the ‘dentary’ is actually composed of a tooth-bearing adsymphyseal (forming the lingual tooth row) co-ossified lingually to the dentary, and not a coronoid as previously thought14,33 (Fig. 3 and Extended Data Fig. 8; see Supplementary Information, section 2 for discussion of adsymphyseal homology).
The pseudoangular of Funcusvermis is highly similar to the postdentary morphology of dissorophoids exemplified by the amphibamid Doleserpeton9 (Fig. 3 and Extended Data Fig. 3); as in Doleserpeton, batrachians, and albanerpetontids, Funcusvermis lacks retroarticular and internal processes, suggesting their initial acquisition in the common ancestor of Eocaecilia and Gymnophiona. The absence of the retroarticular process and presence of a dorsally facing adductor fossa (insertion site of the m. adductor mandibulae complex (mAM)) occupying more than 30% of pseudoangular length (Supplementary Table 1) in the pseudoangular of Funcusvermis (Fig. 1) illuminate a major transformation from the ancestral lissamphibian condition to the unique musculoskeletal architecture of living gymnophionans. Jaw closure driven primarily by the mAM is ancestral for tetrapods, and is retained in batrachians, albanerpetontids and Funcusvermis, differing from the condition of all other gymnophionomorphs, which exhibit the distinctive caecilian dual jaw closure mechanism15 (DJCM). The DJCM is driven primarily by the hyobranchial muscle m. interhyoidus posterior (mIHP), and secondarily by the mAM; the mIHP inserts onto the ventral side of the retroarticular process and extends posteroventrally, acting as a first-order lever causing the anterior component of the lower jaw to pivot upwards with respect to the quadrate during jaw closure15. Acquisition of DJCM is hypothesized to be an adaptation for fossoriality: the mIHP contribution to bite force allows for reduction of the mAM and therefore compaction of the skull roof15,16, a suite of transformations shown to be acquired by the common ancestor of Eocaecilia and Gymnophiona. Although the skull roof of Funcusvermis is unknown, absence of the DJCM and our phylogenetic results suggest that it probably retained the plesiomorphic condition of cheek emargination (gymnokrotaphy, as in Gerobatrachus, batrachians, albanerpetontids and presumably the common ancestor of Lissamphibia) to accommodate the mAM, rather than a closed skull roof with large interpterygoid vacuities13 (stegokrotaphy, as in dissorophoids).
The obtuse angle of the orbital margin in the Funcusvermis maxillopalatine (Fig. 1) may suggest the presence of large orbits as in dissorophoids, batrachians and albanerpetontids (differing from the reduced orbits of other gymnophionomorphs); however, the incomplete orbital margin in the single maxillopalatine specimen (PEFO 46481) prohibits conclusive assessment of this feature. The orbital margin of Funcusvermis lacks a tentacular fossa or aperture (osteological correlates for the chemosensory tentacle organ18), suggesting its absence in early gymnophionomorphs and later derivation by the common ancestor of Eocaecilia and Gymnophiona12. The presence of a co-ossified maxilla and palatine (maxillopalatine) in Funcusvermis is shared with gymnophionans and differs from that of amphibamiforms, albanerpetontids and batrachians, evidence of maxillopalatine consolidation early in gymnophionomorph evolution; however, these bones are possibly separate in Eocaecilia12 and Rubricacaecilia23. Ventrally, the maxillopalatine of Funcusvermis bears parallel maxillary and palatal rows of tightly packed pedicellate teeth of similar size to those in the pseudodentary, seemingly intermediate between the condition of these dentitions in Doleserpeton and Eocaecilia, sharing an anterior truncation of the palatal tooth row by the internal nares with the former, and mesiodistal distal extension (through addition of new teeth) of the palatal row with the latter. A comprehensive comparative description of the Funcusvermis skull and postcranial elements is included in Supplementary Information, section 2.
Evolution of caecilian fossoriality
Given our phylogenetic results, the ecological habits of Funcusvermis may be transitional between terrestrial amphibamid dissorophoids and fossorial gymnophionans. The compound bones in the compact skull of fossorial gymnophionans are thought to withstand the forces associated with head-first burrowing17, and at least some are present in Funcusvermis (for example, maxillopalatine). Small pits covering the lateral surfaces of the pseudodentary and maxillopalatine in Funcusvermis are also found in Eocaecilia12, Rubricacaecilia23 and gymnophionans38. External structure and internal microanatomy of these pits revealed by osteohistological sectioning of a Funcusvermis pseudodentary (PEFO 44432) show a marked resemblance to those of studied living caecilians (Extended Data Fig. 4), in which these pits act as anchor sites for collagen networks forming a tight skin-to-bone attachment and house glands that produce a lubricating mucus secretion, functions thought to aid in subterranean burrowing39. The dorsally flattened neural arch of the Funcusvermis postatlantal pleurocentrum (PEFO 45810; Extended Data Fig. 3) resembles those of Rubricacaecilia23, suggesting the acquisition of a tubular trunk, a feature crucial for underground locomotion in living caecilians40. These morphologies in Funcusvermis illustrate acquisition (by at least the Late Triassic) of some features that now facilitate fossoriality in living caecilians, later followed by acquisition of the DJCM and tentacular organ in Eocaecilia, and finally loss of the appendicular skeleton in gymnophionans.
Biogeography of early caecilians
The spatiotemporal occurrence of Funcusvermis empirically establishes lissamphibian geographic origins on the Pangaean supercontinent before its fragmentation20, and the similar palaeogeography of Eocaecilia12 to Funcusvermis suggests the non-gymnophionan gymnophionomorph origin may lie in the early Mesozoic era of equatorial central Pangaea. The occurrence of Rubricacaecilia in the Early Cretaceous epoch of equatorial Gondwana may further support this hypothesis, suggesting non-gymnophionan gymnophionomorph distribution across both Laurasian and Gondwanan components of Pangaea in the early Mesozoic prior to its breakup23. The equatorial provenance of Funcusvermis adds to an exclusively equatorial pattern of gymnophionomorph distribution: all fossil occurrences fall between a minimum of approximately 16° N and 27° S (Fig. 2 and Supplementary Table 2), and living caecilians are restricted to equatorial latitudes19 between 27° N and 34° S. The tropical distribution of extant gymnophionans is notably disjunct from non-gymnophionan gymnophionomorph fossil occurrences in present-day western North America and Morocco (Fig. 2). Drift of the North American and African plates during the Mesozoic41 may explain the extirpation of gymnophionomorphs from these areas later in the Phanerozoic as these previously humid palaeotropical regions moved north into the arid subtropics. Concurrently, the northern drift of Gondwana into the palaeotropics may have expanded suitable terrestrial habitats, consistent with molecular evidence of an early Mesozoic Gondwanan origin of gymnophionans20.

a, Biogeographic history of Gymnophionomorpha and Triassic batrachians; yellow indicates living caecilian distribution. b, Time-calibrated topology of lissamphibian relationships showing major divergences (topology derived from refs. 6,23,38). Estimated molecular divergence dates for major divergences are shown as blue circles (Gymnophionomopha–Batrachia divergence without Gerobatrachus calibration; Supplementary Table 4), pink circles (Gymnophionomopha–Batrachia divergence with Gerobatrachus calibration; Supplementary Table 5), yellow circles (Salientia–Caudata divergence; Supplementary Table 6) and green circles (Rhinatrematidae–Stegokrotaphia divergence; Supplementary Table 7); coloured vertical bars show the average for each set of divergence estimates. Numbered white and orange circles correspond to occurrences in Supplementary Tables 2 and 3, respectively. Crosses indicate extinct taxa.

Squares denote important apomorphies (including non-mandibular features); apomorphies are optimized computationally unless followed by an asterisk, which denotes an apomorphy suggested by our results but lacking sufficient sampling to optimize computationally. Topology is derived from parsimony results (Extended Data Fig. 5); Yaksha peretti, Salamandra and Rana approximate conditions are found in taxa sampled in the analysis. Illustrations represent right mandibles in medial (bottom) and dorsal (top) views for Doleserpeton annectens9, Eocaecilia micropodia12 (Illustration adapted from ref. 12, with the permission of Museum of Comparative Zoology, Harvard University), Epicrionops petersi12, Funcusvermis gilmorei, Rana, Salamandra and Y. peretti28, excepting Greererpeton burkemorani35 (dorsal only) and Cacops aspidephorus37 (medial only). All scale bars are 2 mm except for G. burkemorani (2 cm) and C. aspidephorus (2 cm). Brackets on the branches indicate stem groups, whereas circles indicate node groups. Crosses indicate extinct taxa.
The earliest batrachians hail from the Triassic of southern2, equatorial4,5 and northern3,6 Pangaea (Supplementary Table 3), indicating extensive latitudinal dispersal by at least the Middle Triassic epoch; this pattern is further reflected in the subsequent batrachian fossil record and their extant distribution. Unlike in extant batrachians, evaporative water loss is found to be a critical physiological constraint in living caecilians, limiting their distribution to humid environments near the equator19. The contrasting spatiotemporal histories of batrachians and gymnophionomorphs suggest a divergence of physiological constraints linked to humidity prior to the Triassic; conserved physiological traits in these groups may explain subsequent patterns of dispersal reflected in present-day lissamphibian biogeography.
Timing of lissamphibian origins
Prior to the results of this study, the chronology of lissamphibian origins remained unresolved owing to the reliance of molecular clock estimates on different node minima derived from competing phylogenetic hypotheses that include extinct taxa1. Funcusvermis lends novel and strong support for a monophyletic origin of living amphibians within dissorophoid temnospondyls30,42 (the ‘classic’ temnospondyl hypothesis), and thus the molecular clock estimates of caecilian–batrachian divergence using the temnospondyl hypothesis. Additionally, the recovery of Gerobatrachus as the sister taxon to Lissamphibia in our analysis suggests that taxon may not be a stem batrachian7,8 and should be used with caution as a minimum age calibration for Lissamphibia. Molecular clock estimates using the temnospondyl hypothesis topology unconstrained by Gerobatrachus as the minimum age calibration of Lissamphibia may result in the most accurate estimates of the caecilian–batrachian divergence, and studies following these criteria show divergence time estimates ranging from the Late Devonian (367.0 Ma) to Middle Pennsylvanian (314.8 Ma) epochs, with a mean in the Middle Mississippian (333.5 Ma) and a median in the Late Mississippian (325.6 Ma) epoch (Fig. 2b and Supplementary Table 4). Our results refocus the timeframe of lissamphibian origins to the Mississippian subperiod, older than previous estimates of a Pennsylvanian-Permian divergence based on calibrations using Gerobatrachus or Amphibamus grandiceps1 and those considering Gymnophionomorpha as the sister group to the stereospondyl Chinlestegophis1,14 (Fig. 2 and Supplementary Table 5).
Methods
New phylogenetic definition
Gymnophionomorpha Marjanović and Laurin 2008
Remarks. Gymnophionomorpha is defined here as the total group consisting of Caecilia tentaculata and all taxa that share a more recent common ancestor with it than with Salamandra salamandra Linnaeus, 1758, Rana temporaria Linnaeus, 1758, and Albanerpeton inexpectatum, Estes and Hoffstetter 1976. This newly proposed stem-based definition of Gymnophionomorpha is modified after that originally proposed43.
Assignment of elements
Although all specimens assigned to Funcusvermis were found as isolated, dissociated elements, their assignment to a single gymnophionomorph taxon is supported by: (1) specimens bearing a suite of features present exclusively in gymnophionomorphs to the exclusion of all other tetrapods (Supplementary Information, section 2); (2) skeletal elements represented by multiple specimens (77 pseudodentaries and 8 pseudoangulars) where all are identical in morphology, varying only in size (Supplementary Information, sections 1 and 2); (3) the pseudodentary and pseudoangular bear complementary facets where they would overlap when in articulation (Extended Data Fig. 3).
Geological framework
The blue-coloured strata of the upper Blue Mesa Member of the Chinle Formation were deposited in a northwest-flowing fluviolacustrine system on the western margin of central Pangaea at a palaeolatitude of 5° to 15° N in a humid monsoonal climate26. Detrital zircon U-Pb radiometric ages provide robust geochronologic constraints on the Chinle Formation, bracketing deposition of the upper Blue Mesa Member24,25 to ~223–218 Ma (Extended Data Fig. 2). The gymnophionomorph fossils described herein were collected from the Thunderstorm Ridge locality (PFV 456) near the Puerco River in PEFO, Arizona, USA (Extended Data Fig. 2). The fossiliferous unit is a 15-cm-thick, poorly sorted siltstone horizon, bearing a dense concentration of carbonate nodules, angular intraformational clasts, micro- and macrovertebrate bones and coprolites. PFV 456 has yielded a diverse assemblage of vertebrates including chondrichthyans, actinopterygians, dipnoans, coelacanths, metoposaurids, salentians5, drepanosauromorphs44, lepidosauromorphs, archosauromorphs45, pseudosuchian archosaurs46, dinosauromorphs47 and cynodonts48. The lack of abrasion and polishing and the exceptional three-dimensional preservation of extremely delicate microvertebrate bones indicates initial deposition in a low-energy setting, followed by brief reworking and redeposition in a channel avulsion event that incorporated angular intraformational clasts and carbonate nodules into the fossiliferous layer. This sedimentological evidence in combination with the presence of abundant spinicaudatan exoskeletons, unionid bivalve steinkerns and obligate-aquatic, amphibious and fully terrestrial vertebrates indicates initial deposition in a marginal lacustrine palaeoenvironment occupied by a diverse vertebrate community.
Collection and preparation methods
The hypodigm and all referred specimens were collected by screenwashing fossiliferous matrix from PFV 456 (9 out of 11 fossil gymnophionomorph occurrences were recovered using screenwashing; Supplementary Table 2). Blocks of matrix weighing approximately 1.8–3.2 kg were individually disaggregated in water and subsequently washed through a series of wire mesh screens with a minimum screen opening of 0.5 mm (no. 35 mesh). Dividing the fossiliferous concentrate from each block into smaller fractions in this way accelerated the process of picking. The resulting concentrate fractions were picked using a dissecting microscope resulting in the identification and separation of all Funcusvermis specimens. Importantly, through processing individual blocks of matrix, Funcusvermis elements that fragmented into multiple parts during the screenwashing process could be re-associated after microscopic sorting. Elements found as multiple broken pieces were subsequently reassembled by adhering matching fractured surfaces using cyanoacrylate, typically a low viscosity PaleoBOND or Loctite brand. To facilitate rapidly and precisely adhering these miniscule fragments together we created a mechanism that combines aspects of a jeweler’s block ball vice, and a hobbyist tool, sometimes called a third hand or helping hand. It combines a socket made of wood or closed cell polyethylene foam and a hemispherical wooden ball to create a pivot that can turn or tilt in all directions. This is topped with a small rectangle of wood with a small concave arch cut into it to provide a workspace. Insect pins are slid through channels in the wood filled with soft microcrystalline wax, which allows the pins freedom of movement, but the resistance needed to precisely position the fossil fragments. The fragments are temporarily adhered to the pin tips with more microcrystalline wax. Adhesive was applied to the joint between fragments as a microdroplet suspended on a single filament such as a cotton fibre and drawn into the joint via capillary action leaving a minimum of excess residue. Reassembly took place under a variety of Leica and Wild binocular microscopes, primarily MZ6, MZ12 and M8 models, varying in power from a maximum of ×40–×80 magnification.
To reveal the details of the pseudodentary dentition of Funcusvermis, matrix covering the dentition and other anatomy of PEFO 45800 was prepared through the following process. Melted cyclododecane (CDD) was poured into a shallow ceramic watch glass and allowed to harden. A small trench the size of the specimen was excavated, and the specimen was placed in the trench in the desired orientation. A Ukrainian kistky (a wax pen), was used to melt the CDD around the specimen and allowed it to adhere to and support the specimen. Then matrix was removed using a 1/32 inch (0.79375 mm) carbide-needle in a pin vice primarily under high magnification under a Leica MZ12 and MZ6 microscope. The point of the needle was ground to a superfine conical point at about 10°–15° parallel to the shaft and flattened briefly along one side to provide an edge to remove adhesives. Some of the softer clay particles were removed with a porcupine quill. When needed, the specimen was consolidated with a very dilute solution of polyvinyl butyral (Butvar B-76) in acetone; the solution was mixed by eye, applying a bit to another vertebrate bone fragment and looking for sheen upon drying. Any excess Butvar film was removed by abrasion with the porcupine quill. The specimen was rotated in the CDD by trenching around the specimen until it was loose, shifting it, and then remelting the resulting CDD powder with the kistky. After all matrix was removed, the specimen was trenched out a final time and set aside in the fume hood to allow the CDD to sublimate.
Digital photography methods
Photographs of PEFO 45800 in Fig. 1 were acquired using a Leica MZ67 stereomicroscope and a Sony NEX-5T digital camera. Image stacking was conducted in Adobe Photoshop CC (https://www.adobe.com/products/photoshop.html).
Micro-computed tomographic scan methods
PEFO 44432, PEFO 45800, PEFO 45910, PEFO 46284, PEFO 46480 and PEFO 46481 were CT scanned with a Skyscan 1172 Microfocus X-radiographic Scanner at the Virginia Tech Institute for Critical Technology and Applied Science (ICTAS). PEFO 43891 was scanned with a Nikon XTH 225 ST High-Resolution X-ray Computed Tomography Scanner in the Shared Materials Instrumentation Facility at Duke University. Micro-computed scan parameters (resolution, source voltage, source current and scanning equipment type) for each scanned specimen included in Supplementary Table 8. Surface volume files (3D meshes) of specimens figured in Fig. 1 and Extended Data Figs. 3 and 4 are available for download under project 000382289 at Morphosource.org (https://www.morphosource.org/projects/000382289?locale=en).
3D segmentation methods
Scan datasets were processed using Dragonfly 2020.2 (http://www.theobjects.com/dragonfly) to produce 3D virtual reconstructions. PEFO 43891, PEFO 46284, and PEFO 46481 were segmented in Dragonfly 2020.2 to digitally remove matrix covering parts of the specimens.
Images of 3D surface meshes were produced using Meshlab 2021.07 (https://www.meshlab.net/).
Digital reconstruction methods
A composite reconstruction of a partial skull of Funcusvermis (Fig. 1) was produced using Meshmixer 3.5 (https://meshmixer.com). Digital 3D surface meshes representing the anterior (PEFO 43891; light pink in Fig. 1a–c) and posterior (PEFO 46284; dark pink in Fig. 1a–c) portions of a pseudodentary were scaled to the same dorsoventral height, and both specimens were overlapped to form a composite reconstruction of a complete pseudodentary. The pseudoangular (PEFO 46480) and maxillopalatine (PEFO 46481) were scaled to match the size of the reconstructed pseudodentary, and anatomically positioned relative to the pseudodentary to approximate their position in an articulated three-dimensional skull. A surface volume file (3D mesh) of the composite skull reconstruction is available for download under project 000382289 on Morphosource.org (https://www.morphosource.org/projects/000382289?locale=en).
Osteohistology methods
PEFO 44432 (right pseudodentary) was embedded in clear epoxy (Castolite AP), cut into 1 mm sections, and then ground to a ~100 µm thickness in the Virginia Tech Fossil Preparation Lab. Images of the histologically sectioned pseudodentary slide used in Extended Data Fig. 4 were acquired using a Sony NEX-5T digital camera mounted on a Nikon OPTIPHOT-POL Polarizing microscope. Fracturing of the specimen occurred during osteohistological preparation, causing fracture planes apparent in histological imaging (Extended Data Fig. 4).
Phylogenetic methods
See ‘Code availability’ to access and download phylogenetic matrix and analysis scripts.
Taxon sampling
Recent analyses recovered gymnophionomorphs at variable positions within Tetrapoda dependent on character and taxon sampling, including: (1) as ‘microsaur’ ‘lepospondyls’49 (note that taxa formerly included in ‘Lepospondyli’ are now understood as polyphyletic50); (2) as stereospondyl temnospondyls forming the sister group to C. jenkinsi14; (3) as ‘microsaurian’ or aïstopod ‘lepospondyls’51; and [4] as amphibamiform dissorophoid temnospondyls forming the sister group to batrachians6,13. The matrix of Schoch et al. (2020), recently used to hypothesize the phylogenetic position of the stem salamander Triassurus sixtelae and the origin of lissamphibians, was selected to test the phylogenetic relationships of F. gilmorei given its comprehensive sampling of taxa proposed to be sister groups to Gymnophionomorpha including stem and crown amniotes, stereospondyl and dissorophoid temnospondyl amphibians, batrachians, gymnophionomorphs and albanerpetontids. F. gilmorei was coded into the modified Schoch et al. (2020) matrix, for a total of 63 sampled terminal taxa. See Supplementary Information, section 4 for discussion of taxon sampling.
Character sampling and scoring
Modifications to the Schoch et al. (2020) matrix are detailed in Supplementary Information, section 4 and include addition of new characters, modification of preexisting characters, exclusion of preexisting characters, and recodings of preexisting character states. Funcusvermis was coded for 29 characters in total based on currently known skeletal material (Supplementary Table 9). The final matrix includes 355 morphological characters (Full character list in Supplementary Information, section 8; see ‘Code availability’ to access and download phylogenetic matrix and analysis scripts).
Maximum parsimony and Bayesian analysis
All characters were equally weighted and unordered in both analyses following previous versions6,14. The character–taxon matrix was first analysed in the phylogenetic analysis software package TNT 1.5 (ref. 52) using New Technology Search options with the following parameters: ratchet (1,000 iterations), sectoral search (1,000 rounds), tree fusing (100 rounds), and random additional sequence (1,000 replicates). A total of 71 most parsimonious trees of 1,468 steps each were recovered (consistency index = 0.287; retention index = 0.675). A strict consensus tree calculated from the most parsimonious trees is presented in Extended Data Fig. 5. Bootstrap support values were obtained using TNT 1.5, and a strict consensus topology of trees produced via 1,000 bootstrap replicates resampled with replacement is presented in Extended Data Fig. 6. A Bayesian inference analysis of the character–taxon matrix was conducted in the phylogenetic software package MrBayes v.3.2.6 (ref. 53) with the Mkv54 model and gamma rate variation and the following parameters: four runs (six Markov chain Monte Carlo chains each), sampled every 1,000 generations, for 10 million generations with a relative burn-in of 0.25. Convergence of independent runs was assessed using Tracer v.1.76.1 (http://beast.bio.ed.ac.uk/Tracer). A consensus cladogram with mapped posterior probability values is presented in Extended Data Fig. 7.
Nomenclatural acts
The Life Science Identifiers (LSID) for the new genus and species are registered with Zoobank (http://zoobank.org) under the identifiers urn:lsid:zoobank.org:pub:A2A6C7AD-2077-413B-9004-2E841270A289.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The holotype, paratypes and referred specimens of F. gilmorei are catalogued and available for study to qualified researchers at PEFO. Computed tomographic scan data, including surface volume files (3D meshes) and raw CT data of Funcusvermis specimens mentioned in the main text and extended data figures (including the holotype, paratypes and referred specimens), as well as a surface volume file of the composite skull reconstruction of Funcusvermis are available for download under project 000382289 on Morphosource.org (https://www.morphosource.org/projects/000382289?locale=en).
Code availability
Code for TNT and MrBayes scripts used in the phylogenetic analyses conducted herein are available in Supplementary Information, section 8; the matrix is available for download under project 4166 on Morphobank.org (http://morphobank.org/permalink/?P4166).
References
Pardo, J. D., Lennie, K. & Anderson, J. S. Can we reliably calibrate deep nodes in the tetrapod tree? Case studies in deep tetrapod divergences. Front. Genet. 11, 1159 (2020).
Rage, J.-C. & Roček, Z. Redescription of Triadobatrachus massinoti (Piveteau, 1936) an anuran amphibian from the early Triassic. Palaeontographica A 206, 1–16 (1989).
Evans, S. E. & Borsuk-Białynicka, M. A stem-group frog from the Early Triassic of Poland. Acta Palaeontol. Pol. 43, 573–580 (1998).
Heckert, A. B., Mitchell, J. S., Schneider, V. P. & Olsen, P. E. Diverse new microvertebrate assemblage from the Upper Triassic Cumnock Formation, Sanford Subbasin, North Carolina, USA. J. Paleontol. 86, 368–390 (2012).
Stocker, M. R. et al. The earliest equatorial record of frogs from the Late Triassic of Arizona. Biol. Lett. 15, 20180922 (2019).
Schoch, R. R., Werneburg, R. & Voigt, S. A Triassic stem-salamander from Kyrgyzstan and the origin of salamanders. Proc. Natl Acad. Sci. USA 117, 11584–11588 (2020).
Anderson, J. S., Reisz, R. R., Scott, D., Fröbisch, N. B. & Sumida, S. S. A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders. Nature 453, 515–518 (2008).
Anderson, J. S. Focal review: the origin(s) of modern amphibians. Evol. Biol. 35, 231–247 (2008).
Sigurdsen, T. & Bolt, J. R. The Lower Permian amphibamid Doleserpeton (Temnospondyli: Dissorophoidea), the interrelationships of amphibamids, and the origin of modern amphibians. J. Vertebr. Paleontol. 30, 1360–1377 (2010).
Schoch, R. R. The putative lissamphibian stem-group: phylogeny and evolution of the dissorophoid temnospondyls. J. Paleontol. 93, 137–156 (2019).
Jenkins, P. A. & Walsh, D. M. An Early Jurassic caecilian with limbs. Nature 365, 246–250 (1993).
Jenkins, F. A., Walsh, D. M. & Carroll, R. L. Anatomy of Eocaecilia micropodia, a limbed caecilian of the Early Jurassic. Bull. Mus. Comp. Zool. 158, 285–365 (2007).
Maddin, H. C., Jenkins, F. A. Jr & Anderson, J. S. The braincase of Eocaecilia micropodia (Lissamphibia, Gymnophiona) and the origin of caecilians. PLoS ONE 7, e50743 (2012).
Pardo, J. D., Small, B. J. & Huttenlocker, A. K. Stem caecilian from the Triassic of Colorado sheds light on the origins of Lissamphibia. Proc. Natl Acad. Sci. USA 114, E5389–E5395 (2017).
Nussbaum, R. A. The evolution of a unique dual jaw‐closing mechanism in caecilians: (Amphibia: Gymnophiona) and its bearing on caecilian ancestry. J. Zool. 199, 545–554 (1983).
Kleinteich, T., Haas, A. & Summers, A. P. Caecilian jaw-closing mechanics: integrating two muscle systems. J. R. Soc. Interface 5, 1491–1504 (2008).
Sherratt, E., Gower, D. J., Klingenberg, C. P. & Wilkinson, M. Evolution of cranial shape in caecilians (Amphibia: Gymnophiona). Evol. Biol. 41, 528–545 (2014).
Schmidt, A. & Wake, M. H. Olfactory and vomeronasal systems of caecilians (Amphibia: Gymnophiona). J. Morphol. 205, 255–268 (1990).
Pincheira‐Donoso, D., Meiri, S., Jara, M., Olalla‐Tárraga, M. Á. & Hodgson, D. J. Global patterns of body size evolution are driven by precipitation in legless amphibians. Ecography 42, 1682–1690 (2019).
San Mauro, D., Vences, M., Alcobendas, M., Zardoya, R. & Meyer, A. Initial diversification of living amphibians predated the breakup of Pangaea. Am. Nat. 165, 590–599 (2005).
Padian, K. & Sues, H.-D. in Great Transformations in Vertebrate Evolution (eds Dial, K. P., Shubin, N. & Brainerd, E. L.) 351–374 (Univ. Chicago Press, 2021).
Santos, R. O., Laurin, M. & Zaher, H. A review of the fossil record of caecilians (Lissamphibia: Gymnophionomorpha) with comments on its use to calibrate molecular timetrees. Biol. J. Linn. Soc. 131, 737–755 (2020).
Evans, S. E. & Sigogneau‐Russell, D. A stem‐group caecilian (Lissamphibia: Gymnophiona) from the Lower Cretaceous of North Africa. Palaeontology 44, 259–273 (2001).
Ramezani, J. et al. High-precision U-Pb zircon geochronology of the Late Triassic Chinle Formation, Petrified Forest National Park (Arizona, USA): temporal constraints on the early evolution of dinosaurs. GSA Bull. 123, 2142–2159 (2011).
Rasmussen, C. et al. U-Pb zircon geochronology and depositional age models for the Upper Triassic Chinle Formation (Petrified Forest National Park, Arizona, USA): implications for Late Triassic paleoecological and paleoenvironmental change. GSA Bull. 133, 539–558 (2021).
Nordt, L., Atchley, S. & Dworkin, S. Collapse of the Late Triassic megamonsoon in western equatorial Pangea, present-day American Southwest. GSA Bull. 127, 1798–1815 (2015).
Martz, J. W. & Parker, W. G. in Terrestrial Depositional Systems (eds Zeigler, K. E. & Parker, W. G.) 39–125 (Elsevier, 2017).
Daza, J. D. et al. Enigmatic amphibians in mid-Cretaceous amber were chameleon-like ballistic feeders. Science 370, 687–691 (2020).
Gardner, J. D. Monophyly and affinities of albanerpetontid amphibians (Temnospondyli; Lissamphibia). Zool. J. Linn. Soc. 131, 309–352 (2001).
Bolt, J. R. Lissamphibian origins: possible protolissamphibian from the Lower Permian of Oklahoma. Science 166, 888–891 (1969).
Gardner, J. D. & Averianov, A. O. Albanerpetontid amphibians from the Upper Cretaceous of Middle Asia. Acta Palaeontol. Pol. 43, 453–476 (1998).
Carroll, R. L. The Palaeozoic ancestry of salamanders, frogs and caecilians. Zool. J. Linn. Soc. 150, 1–140 (2007).
Müller, H., Oommen, O. V. & Bartsch, P. Skeletal development of the direct-developing caecilian Gegeneophis ramaswamii (Amphibia: Gymnophiona: Caeciliidae). Zoomorphology 124, 171–188 (2005).
Ahlberg, P. E. & Clack, J. A. Lower jaws, lower tetrapods—a review based on the Devonian genus Acanthostega. Earth Environ. Sci. Trans. R. Soc. Edinb. 89, 11–46 (1998).
Bolt, J. R. & Lombard, R. E. The mandible of the primitive tetrapod Greererpeton, and the early evolution of the tetrapod lower jaw. J. Paleontol. 75, 1016–1042 (2001).
Shishkin, M. A. & Sulej, T. The Early Triassic temnospondyls of the Czatkowice 1 tetrapod assemblage. Acta Palaeontol. Pol. 65, 31–77 (2009).
Anderson, J. S., Scott, D. & Reisz, R. R. The anatomy of the dermatocranium and mandible of Cacops aspidephorus Williston, 1910 (Temnospondyli: Dissorophidae), from the Lower Permian of Texas. J. Vertebr. Paleontol. 40, e1776720 (2020).
Wilkinson, M., San Mauro, D., Sherratt, E. & Gower, D. J. A nine-family classification of caecilians (Amphibia: Gymnophiona). Zootaxa 2874, 41–64 (2011).
Jared, C. et al. Skin gland concentrations adapted to different evolutionary pressures in the head and posterior regions of the caecilian Siphonops annulatus. Sci. Rep. 8, 3576 (2018).
O’Reilly, J. C., Ritter, D. A. & Carrier, D. R. Hydrostatic locomotion in a limbless tetrapod. Nature 386, 269–272 (1997).
Muttoni, G. & Kent, D. V. Jurassic monster polar shift confirmed by sequential paleopoles from Adria, promontory of Africa. J. Geophys. Res. 124, 3288–3306 (2019).
Parsons, T. S. & Williams, E. E. The relationships of the modern Amphibia: a re-examination. Q. Rev. Biol. 38, 26–53 (1963).
Marjanović, D. & Laurin, M. A reevaluation of the evidence supporting an unorthodox hypothesis on the origin of extant amphibians. Contrib. Zool. 77, 149–199 (2008).
Jenkins, X. A. et al. Using manual ungual morphology to predict substrate use in the Drepanosauromorpha and the description of a new species. J. Vertebr. Paleontol. 40, e1810058 (2020).
Kligman, B. T., Marsh, A. D., Nesbitt, S. J., Parker, W. G. & Stocker, M. R. New trilophosaurid species demonstrates a decline in allokotosaur diversity across the Adamanian–Revueltian boundary in the Late Triassic of western North America. Palaeodiversity 13, 25–37 (2020).
Marsh, A. D., Smith, M. E., Parker, W. G., Irmis, R. B. & Kligman, B. T. Skeletal anatomy of Acaenasuchus geoffreyi Long and Murry, 1995 (Archosauria: Pseudosuchia) and its implications for the origin of the aetosaurian carapace. J. Vertebr. Paleontol. 40, e1794885 (2020).
Marsh, A. D. & Parker, W. G. New dinosauromorph specimens from Petrified Forest National Park and a global biostratigraphic review of Triassic dinosauromorph body fossils. PaleoBios https://doi.org/10.5070/P9371050859 (2020).
Kligman, B. T., Marsh, A. D., Sues, H.-D. & Sidor, C. A. A new non-mammalian eucynodont from the Chinle Formation (Triassic: Norian), and implications for the early Mesozoic equatorial cynodont record. Biol. Lett. 16, 20200631 (2020).
Huttenlocker, A. K., Pardo, J. D., Small, B. J. & Anderson, J. S. Cranial morphology of recumbirostrans (Lepospondyli) from the Permian of Kansas and Nebraska, and early morphological evolution inferred by micro-computed tomography. J. Vertebr. Paleontol. 33, 540–552 (2013).
Pardo, J. D., Szostakiwskyj, M., Ahlberg, P. E. & Anderson, J. S. Hidden morphological diversity among early tetrapods. Nature 546, 642–645 (2017).
Marjanović, D. & Laurin, M. Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix. PeerJ 6, e5565 (2019).
Goloboff, P. A. & Catalano, S. A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32, 221–238 (2016).
Huelsenbeck, J. P. & Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17, 754–755 (2001).
Lewis, P. O. A likelihood approach to estimating phylogeny from discrete morphological character data. Syst. Biol. 50, 913–925 (2001).
Eltink, E., Schoch, R. R. & Langer, M. C. Interrelationships, palaeobiogeography and early evolution of Stereospondylomorpha (Tetrapoda: Temnospondyli). J. Iber. Geol. 45, 251–267 (2019).
Bystrow, A. Dvinosaurus als neotenische Form der Stegocephalen. Acta Zool. 19, 209–295 (1938).
Dutuit, J.-M. Introduction à l’étude paléontologique du Trias continental Marocain. Description des premiers stegocephales recueillis dans le couloir d’Argana (Atlas Occidental). Mémoires du Muséum National d’Histoire 36, 1–253 (1976).
Dias, E. V., Dias-da-Silva, S. & Schultz, C. L. A new short-snouted rhinesuchid from the Permian of southern Brazil. Revista Brasileira de Paleontologia 23, 98–122 (2020).
Damiani, R. J. & Kitching, J. W. A new brachyopid temnospondyl from the Cynognathus Assemblage Zone, Upper Beaufort Group, South Africa. J. Vertebr. Paleontol. 23, 67–78 (2003).
Schoch, R. R. & Witzmann, F. Cranial morphology of the plagiosaurid Gerrothorax pulcherrimus as an extreme example of evolutionary stasis. Lethaia 45, 371–385 (2012).
Schoch, R. R. Studies on braincases of early tetrapods: Structure, morphological diversity, and phylogeny-1 Trimerorhacis and other prmitive temnospondyls. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen 213, 233–259 (1999).
Ruta, M. & Bolt, J. R. The brachyopoid Hadrokkosaurus bradyi from the early Middle Triassic of Arizona, and a phylogenetic analysis of lower jaw characters in temnospondyl amphibians. Acta Palaeontol. Pol. 53, 579–592 (2008).
Bystrow, A. & Efremov, J. Benthosuchus sushkini Efr.—a labyrinthodont from the Eotriassic of Sharzhenga River. Trudy Paleontol. Inst. 10, 1–152 (1940).
Warren, A. Karoo tupilakosaurid: a relict from Gondwana. Earth Environ. Sci. Trans. R. Soc. Edinb. 89, 145–160 (1998).
Holmes, R. B., Carroll, R. L. & Reisz, R. R. The first articulated skeleton of Dendrerpeton acadianum (Temnospondyli, Dendrerpetontidae) from the Lower Pennsylvanian locality of Joggins, Nova Scotia, and a review of its relationships. J. Vertebr. Paleontol. 18, 64–79 (1998).
Steyer, J. S. The first articulated trematosaur ‘amphibian’ from the Lower Triassic of Madagascar: implications for the phylogeny of the group. Palaeontol. 45, 771–793 (2002).
Englehorn, J., Small, B. J. & Huttenlocker, A. A redescription of Acroplous vorax (Temnospondyli: Dvinosauria) based on new specimens from the Early Permian of Nebraska and Kansas, USA. J. Vertebr. Paleontol. 28, 291–305 (2008).
Warren, A. Laidleria uncovered: a redescription of Laidleria gracilis Kitching (1957), a temnospondyl from the Cynognathus Zone of South Africa. Zool. J. Linn. Soc. 122, 167–185 (1998).
Bolt, J. R. & Chatterjee, S. A new temnospondyl amphibian from the Late Triassic of Texas. J. Paleontol. 74, 670–683 (2000).
Milner, A. & Sequeira, S. The temnospondyl amphibians from the Viséan of east Kirkton, West Lothian, Scotland. Earth Environ. Sci. Trans. R. Soc. Edinb. 84, 331–361 (1993).
Schoch, R. R. & Milner, A. R. Encyclopedia of Paleoherpetology, Part 3A. Temnospondyli (Verlag Dr. Friedrich Pfeil, 2014).
Damiani, R., Schoch, R. R., Hellrung, H., Werneburg, R. & Gastou, S. The plagiosaurid temnospondyl Plagiosuchus pustuliferus (Amphibia: Temnospondyli) from the Middle Triassic of Germany: anatomy and functional morphology of the skull. Zool. J. Linn. Soc. 155, 348–373 (2009).
Chernin, S. A new brachyopid, Batrachosuchus concordi sp. nov. from the Upper Luangwa Valley, Zambia with a redescription of Batrachosuchus browni Broom, 1903. Palaeontol. Afr. 20, 87–109 (1977).
Sulej, T. Osteology, variability, and evolution of Metoposaurus, a temnospondyl from the Late Triassic of Poland. Acta Palaeontol. Pol. 64, 29–139 (2007).
Acknowledgements
We thank C. Beightol, W. Reyes, E. Patellos, X. Jenkins, E. Smith and members of the 2019 Virginia Tech Paleobiology Research Group field team for their fieldwork efforts. D. Wagner, D. Boudreau, P. Varela, L. McWhinney, N. Pezzoni, N. Green, C. Heltzel, C. Sheehy and Z. Lester conducted microscopic picking resulting in the collection of many specimens. C. Lash, D. Wagner, D. Boudreau and P. Varela provided logistical support in the PEFO laboratory and collections. R. Mueller and J. Socha facilitated μCT scanner access at Virginia Tech. J. Gladman facilitated μCT scanner access at the Shared Materials Instrumentation Facility at Duke University. J. Gillette and D. Gillette (MNA) provided access to Eocaecilia specimens. J. Martz illustrated the stratigraphy of PEFO in Extended Data Fig. 2. B. Creisler assisted in formulating the name of the new taxon. T. Young facilitated funding from the Petrified Forest Museum Association for the illustration of F. gilmorei by A. Atuchin. This study was supported by the David R. Wones Geological Scholarship (to B.T.K.), the USA National Park Service (PMIS 209814 to B.T.K., W.G.P. and A.D.M.), the Virginia Tech Department of Geosciences (to B.T.K., M.R.S. and S.J.N.), the National Science Foundation (DEB1655609 to M.R.S.; EAR480383 to S.J.N., M.R.S. and B.T.K.; and DPP1947094 to C. Sidor, supporting B.M.G.), the David B. Jones Foundation (to M.R.S. and S.J.N.) and the Petrified Forest Museum Association (to B.T.K.). We thank the Willi Henning Society for free access to TNT software. This is Petrified Forest National Park Contribution no. 87. Views expressed herein are those of the authors and do not represent the views of the United States Government. Finally, we thank James ‘Diamond’ Williams and the Ohio Players for their permission to name the genus after their song Funky Worm from the 1972 album Pleasure.
Author information
Authors and Affiliations
Contributions
B.T.K., A.D.M., W.G.P. and M.R.S. conceived the project. B.T.K., A.D.M., W.G.P., M.R.S. and S.J.N. conducted fieldwork collecting fossiliferous sediment. M.E.S., B.T.K. and A.D.M. conducted laboratory work identifying and physically preparing specimens. B.T.K. and M.R.S. sampled μCT data. B.T.K. conducted μCT digital processing and preparation. S.J.N. and B.T.K. conducted histological sampling. B.M.G. led revisions of the phylogenetic dataset with assistance from B.T.K. B.M.G. and B.T.K. coded the phylogenetic matrix. B.T.K. and B.M.G. conducted phylogenetic analyses. B.T.K., A.D.M. and B.M.G. prepared figures. B.T.K. wrote the manuscript, with contributions and editing from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Jason Anderson, Rainer Schoch and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Life reconstruction of Funcusvermis gilmorei (lower) and Acaenasuchus geoffreyi (upper) in a paleoenvironmental reconstruction of PFV 456.
Illustration by Andrey Atuchin.
Extended Data Fig. 3 Anatomy of Funcusvermis gilmorei based on reconstructed 3D surface meshes from segmented micro-computed tomographic scan data.
a–c, Referred left pseudoangular (PEFO 46480) in ventral, lateral, and posterior views. e–i, Holotype right pseudodentary (PEFO 43891) in medial, anteromedial, anterior, ventral, dorsal, and anterolateral views. j–l, Paratype right pseudodentary (PEFO 46284) in medial, dorsal, and lateral views. m–o, Paratype right pseudodentary (PEFO 45800) in dorsal, medial, and lateral views. p, Paratype right pseudodentary (PEFO 46284) and referred left pseudoangular (PEFO 46480) with yellow arrows showing matching facet surfaces and neurovascular canals. q–u, Referred postatlantal vertebra (PEFO 45810) in dorsal, ventral, right lateral, anterior, and posterior views. v–y, Referred right femur (PEFO 43811) in dorsal, ventrolateral, lateral, and proximal views. abcnV, alveolar branch cranial nerve V; adtr, adsymphyseal tooth row; aeecnV, anterior exit external branch of cranial nerve V; c, centrum; cnVII, cranial nerve VII insertion; dp, diapophysis; dpaf, dorsal pseudoangular facet; dpdf, dorsal pseudodentary facet; dtr, dentary tooth row; ebcnV, external branch cranial nerve V; fh, femoral head; lasdg, labial subdental groove; ld, lateral depression; lecnV, lateral exit cranial nerve V; lisdg, lingual subdental groove; mf, medial fossa; mkv; midventral keel; ms, mandibular symphysis; mt, medial trochanter; na, neural arch; nc, neural canal; p, pit; paaspd, pseudoangular attachment surface of the pseudodentary; pap, posterior pseudoangular process; pdaspa, pseudodentary attachment surface of the pseudoangular; peecnV, posterior exit external branch cranial nerve V; poz, postzygapophysis; prez, prezygapophysis; ptadtr, posterior terminus adsymphyseal tooth row; ptdtr, posterior terminus dentary tooth row; sf, symphyseal foramen; sp, symphyseal prongs; vpaf, ventral pseudoangular facet; vpdf, ventral pseudodentary facet. Arrows point in anterior direction. Scale bars equal 1 mm.
Extended Data Fig. 4 Cross sectional views of Funcusvermis gilmorei dentigerous elements from micro-computed tomographic scan data and osteohistological sectioning.
a–d, Coronal cross sections of referred left maxillopalatine (PEFO 46481). e–k, Coronal cross sections of holotype right pseudodentary (PEFO 43891). l–n, Coronal cross sections of paratype right pseudodentary (PEFO 46284). o–p, Microanatomy from osteohistological section of paratype right pseudodentary (PEFO 44432) in coronal view. r, Referred left maxillopalatine (PEFO 46481) in lateral view. Paratype right pseudodentary (PEFO 44432) in lateral (s) and medial (t) views. abcnV, alveolar branch cranial nerve V; dpdf, dorsal pseudodentary facet; ebcnV, external branch cranial nerve V; frac, fractures; fr, facial ramus; ig, interdental groove; laam, labial alveolar margin; lapw, labial pedicel wall; lasdg, labial subdental groove; liam, lingal alveolar margin; lipw, lingual pedicel wall; lisdg, lingual subdental groove; oc, osteocyte lacunae; om, orbital margin; p, pit; pc, pulp cavity; peecnV, posterior exit external branch cranial nerve V; sf, symphyseal foramen; vpdf, ventral pseudodentary facet. Red lines indicate locations of cross sections. Yellow outlines denote margins of canals connected to external pits. Scale bars equal 1 mm unless otherwise noted.
Extended Data Fig. 5 Phylogenetic tree from parsimony analysis.
Strict consensus tree of 71 most parsimonious trees (1,468 steps each) from TNT parsimony analysis (Methods). CI = 0.287, RI = 0.675.
Extended Data Fig. 6 Phylogenetic tree from parsimony analysis showing bootstrap support.
Strict consensus topology of trees produced via 1,000 bootstrap replicates resampled with replacement (Methods). Node values indicate bootstrap support.
Extended Data Fig. 7 Topology from Bayesian inference phylogenetic analysis.
Majority-rule consensus tree from Bayesian inference analysis (Methods). Node labels show posterior probability values.
Extended Data Fig. 8 Phylogenetic distribution and comparative morphology of a single row of coronoid teeth in Temnospondyli.
a, Phylogenetic topology (derived from refs. 14,55) with presence of a semi-continuous row of teeth across all three coronoids (See Supplementary Information 3 for further discussion of this feature). b–k, Comparative silhouettes of lower jaws in lingual (b–g), and dorsal (h–m), views. b, The dvinosaur Dvinosaurus primus56. c, The trematosaur Almasaurus habbazi57. d, The rhinesuchid Rastosuchus hammeri58. e, The brachyopid Vanastega plurimidens59. f, The stereospondyl Chinlestegophis jenkinsi14. g, The plagiosaurid Gerrothorax pulcherrimus60 (Illustration adapted from ref. 60 under CC BY 3.0 license). h, The capitosaur Mastodonsaurus giganteus61. i, The brachyopid Hadrokkosaurus bradyi62. j, The trematosaur Benthosuchus sushkini63. k, Chinlestegophis jenkinsi14. Arrows indicate anterior direction.
Extended Data Fig. 9 Phylogenetic distribution and comparative morphology of a lateral exposure of the palatine (LEP) and the loss of the lacrimal in Temnospondyli.
a, Phylogenetic topology (derived from refs. 14,55) with variable presence of the lacrimal and the LEP (See Supplementary Information 3 for further discussion of this feature). Comparative silhouettes of skulls in dorsal b–h and lateral i–k views; the prefrontal is coloured yellow here as it is the main element to positionally compensate for an absent lacrimal. b, The dvinosaur Thabanchuia oomie64 (Illustration reproduced with permission from Cambridge University Press from ref. 64). c, The amphibamid Doleserpeton annectens9 (Illustration adapted from ref. 9 under copyright © Society of Vertebrate Paleontology, www.vertpaleo.org, reprinted by permission of Informa UK Limited, trading as Taylor & Francis Group, www.tandfonline.com on behalf of Society of Vertebrate Paleontology, www.vertpaleo.org. d, The ‘dendrerpetid’ Dendrerpeton helogenes65, which represents the plesiomorphic condition (Illustration adapted from ref. 65. under copyright © Society of Vertebrate Paleontology, www.vertpaleo.org, reprinted by permission of Informa UK Limited, trading as Taylor & Francis Group, www.tandfonline.com on behalf of Society of Vertebrate Paleontology, www.vertpaleo.org). e, The trematosaur Wantzosaurus elongatus66. f, The dvinosaur Acroplous vorax67 (Illustration adapted from ref. 67 under copyright © Society of Vertebrate Paleontology, www.vertpaleo.org, reprinted by permission of Informa UK Limited, trading as Taylor & Francis Group, www.tandfonline.com on behalf of Society of Vertebrate Paleontology, www.vertpaleo.org). g, the rhytidosteid Laidleria gracilis68 (Illustration adapted from ref. 68. by permission of the Zoological Journal of the Linnean Society). h, Chinlestegophis jenkinsi14. i, Acroplous vorax67 in lateral view. j, Rileymillerus cosgriffi69 in lateral view. k, Chinlestegophis jenkinsi14 in lateral view. The prefrontal is also coloured yellow here as it is the main element to positionally compensate for an absent lacrimal. Silhouettes are not to scale.
Extended Data Fig. 10 Phylogenetic distribution and comparative morphology of a broad cultriform process and a posteriorly situated occiput in Temnospondyli.
a, Phylogenetic topology (derived from refs. 14,55) with presence of a broad cultriform process and a posteriorly situated occiput (See Supplementary Information 3 for further discussion of this feature). b–j, Comparative silhouettes of skulls in lateral i–k, and ventral b–h, views. b, The dvinosaur Dvinosaurus primus56. c, The ‘dendrerpetid’ Balanerepeton woodi70,71 (Illustration adapted from ref. 70. with permission from Cambridge University Press), which represents the plesiomorphic condition. d, The plagiosaurid Plagiosuchus pustuliferus72 (Illustration adapted from ref. 72. by permission of the Zoological Journal of the Linnean Society). e, The brachyopid Batrachosuchus browni73. f, The metoposaurid Metoposaurus krasiejowensis74. g, Chinlestegophis jenkinsi14. h, Batrachosuchus browni73. i, The dvinosaur Thabanchuia oomie64 (Illustration reproduced with permission from Cambridge University Press from ref. 64). j, Chinlestegophis jenkinsi14. Silhouettes are not to scale.
Supplementary information
Supplementary Information
This supplementary information file contains the following sections: 1. Complete list of material assigned to F. gilmorei. 2. Expanded description of F. gilmorei. 3. Comparisons to stereospondyl temnospondyls and C. jenkinsi. 4. Revisions and additions to phylogenetic dataset. 5. Supplementary Tables 1–9. 6. Institutional abbreviations. 7. Supplementary references. 8. Character list and phylogenetic datasets.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kligman, B.T., Gee, B.M., Marsh, A.D. et al. Triassic stem caecilian supports dissorophoid origin of living amphibians. Nature 614, 102–107 (2023). https://doi.org/10.1038/s41586-022-05646-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05646-5
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
