
Abstract
Among the caspases that cause regulated cell death, a unique function for caspase-7 has remained elusive. Caspase-3 performs apoptosis, whereas caspase-7 is typically considered an inefficient back-up. Caspase-1 activates gasdermin D pores to lyse the cell; however, caspase-1 also activates caspase-7 for unknown reasons1. Caspases can also trigger cell-type-specific death responses; for example, caspase-1 causes the extrusion of intestinal epithelial cell (IECs) in response to infection with Salmonella enterica subsp. enterica serovar Typhimurium (S. Typhimurium)2,3. Here we show in both organoids and mice that caspase-7-deficient IECs do not complete extrusion. Mechanistically, caspase-7 counteracts gasdermin D pores and preserves cell integrity by cleaving and activating acid sphingomyelinase (ASM), which thereby generates copious amounts of ceramide to enable enhanced membrane repair. This provides time to complete the process of IEC extrusion. In parallel, we also show that caspase-7 and ASM cleavage are required to clear Chromobacterium violaceum and Listeria monocytogenes after perforin-pore-mediated attack by natural killer cells or cytotoxic T lymphocytes, which normally causes apoptosis in infected hepatocytes. Therefore, caspase-7 is not a conventional executioner but instead is a death facilitator that delays pore-driven lysis so that more-specialized processes, such as extrusion or apoptosis, can be completed before cell death. Cells must put their affairs in order before they die.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
All relevant data are included in the Article or its Supplementary Information files. More details are available from the corresponding author upon request. Source data are provided with this paper.
References
Lamkanfi, M. et al. Targeted peptidecentric proteomics reveals caspase-7 as a substrate of the caspase-1 inflammasomes. Mol. Cell. Proteomics 7, 2350–2363 (2008).
Sellin, M. E. et al. Epithelium-intrinsic NAIP/NLRC4 inflammasome drives infected enterocyte expulsion to restrict Salmonella replication in the intestinal mucosa. Cell Host Microbe 16, 237–248 (2014).
Rauch, I. et al. NAIP-NLRC4 inflammasomes coordinate intestinal epithelial cell expulsion with eicosanoid and IL-18 release via activation of caspase-1 and -8. Immunity 46, 649–659 (2017).
Kuida, K. et al. Decreased apoptosis in the brain and premature lethality in CPP32-deficient mice. Nature 384, 368–372 (1996).
Lakhani, S. A. et al. Caspases 3 and 7: key mediators of mitochondrial events of apoptosis. Science 311, 847–851 (2006).
Wu, C. et al. BioGPS: an extensible and customizable portal for querying and organizing gene annotation resources. Genome Biol. 10, R130 (2009).
Reikvam, D. H. et al. Depletion of murine intestinal microbiota: effects on gut mucosa and epithelial gene expression. PLoS One 6, e17996 (2011).
Marchiando, A. M. et al. The epithelial barrier is maintained by in vivo tight junction expansion during pathologic intestinal epithelial shedding. Gastroenterology 140, 1208–1218 (2011).
Nozaki, K., Li, L. & Miao, E. A. Innate sensors trigger regulated cell death to combat intracellular infection. Annu. Rev. Immunol. 40, 469–498 (2022).
Andrews, N. W., Almeida, P. E. & Corrotte, M. Damage control: cellular mechanisms of plasma membrane repair. Trends Cell Biol. 24, 734–742 (2014).
Edelmann, B. et al. Caspase-8 and caspase-7 sequentially mediate proteolytic activation of acid sphingomyelinase in TNF-R1 receptosomes. EMBO J. 30, 379–394 (2011).
Ferlinz, K. et al. Functional characterization of the N-glycosylation sites of human acid sphingomyelinase by site-directed mutagenesis. Eur. J. Biochem. 243, 511–517 (1997).
Andrews, N. W., Corrotte, M. & Castro-Gomes, T. Above the fray: surface remodeling by secreted lysosomal enzymes leads to endocytosis-mediated plasma membrane repair. Semin. Cell Dev. Biol. 45, 10–17 (2015).
Llacuna, L., Marí, M., Garcia-Ruiz, C., Fernandez-Checa, J. C. & Morales, A. Critical role of acidic sphingomyelinase in murine hepatic ischemia-reperfusion injury. Hepatology 44, 561–572 (2006).
Müller, A. J. et al. The S. Typhimurium effector SopE induces caspase-1 activation in stromal cells to initiate gut inflammation. Cell Host Microbe 6, 125–136 (2009).
Quan, L. T. et al. Proteolytic activation of the cell death protease Yama/CPP32 by granzyme B. Proc. Natl Acad. Sci. USA 93, 1972–1976 (1996).
Darmon, A. J., Nicholson, D. W. & Bleackley, R. C. Activation of the apoptotic protease CPP32 by cytotoxic T-cell-derived granzyme B. Nature 377, 446–448 (1995).
Chinnaiyan, A. M. et al. Cytotoxic T-cell-derived granzyme B activates the apoptotic protease ICE-LAP3. Curr. Biol. 6, 897–899 (1996).
Gu, Y. et al. Processing and activation of CMH-1 by granzyme B. J. Biol. Chem. 271, 10816–10820 (1996).
Orth, K., Chinnaiyan, A. M., Garg, M., Froelich, C. J. & Dixit, V. M. The CED-3/ICE-like protease Mch2 is activated during apoptosis and cleaves the death substrate lamin A. J. Biol. Chem. 271, 16443–16446 (1996).
Jorgensen, I., Rayamajhi, M. & Miao, E. A. Programmed cell death as a defence against infection. Nat. Rev. Immunol. 17, 151–164 (2017).
Batista, J. H. & da Silva Neto, J. F. Chromobacterium violaceum pathogenicity: updates and insights from genome sequencing of novel Chromobacterium species. Front. Microbiol. 8, 2213 (2017).
Maltez, V. I. et al. Inflammasomes coordinate pyroptosis and natural killer cell cytotoxicity to clear infection by a ubiquitous environmental bacterium. Immunity 43, 987–997 (2015).
Rongvaux, A. et al. Apoptotic caspases prevent the induction of type I interferons by mitochondrial DNA. Cell 159, 1563–1577 (2014).
McArthur, K. et al. BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science 359, eaao6047 (2018).
Sauer, J. D. et al. Listeria monocytogenes engineered to activate the Nlrc4 inflammasome are severely attenuated and are poor inducers of protective immunity. Proc. Natl Acad. Sci. USA 108, 12419–12424 (2011).
Warren, S. E. et al. Generation of a Listeria vaccine strain by enhanced caspase-1 activation. Eur. J. Immunol. 41, 1934–1940 (2011).
Clark, S. E., Schmidt, R. L., McDermott, D. S. & Lenz, L. L. A Batf3/Nlrp3/IL-18 axis promotes natural killer cell IL-10 production during Listeria monocytogenes infection. Cell Rep. 23, 2582–2594 (2018).
Kägi, D., Ledermann, B., Bürki, K., Hengartner, H. & Zinkernagel, R. M. CD8+ T cell-mediated protection against an intracellular bacterium by perforin-dependent cytotoxicity. Eur. J. Immunol. 24, 3068–3072 (1994).
Harty, J. T., Lenz, L. L. & Bevan, M. J. Primary and secondary immune responses to Listeria monocytogenes. Curr. Opin. Immunol. 8, 526–530 (1996).
Hsu, K. M., Pratt, J. R., Akers, W. J., Achilefu, S. I. & Yokoyama, W. M. Murine cytomegalovirus displays selective infection of cells within hours after systemic administration. J. Gen. Virol. 90, 33–43 (2009).
Kägi, D. et al. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature 369, 31–37 (1994).
Rühl, S. et al. ESCRT-dependent membrane repair negatively regulates pyroptosis downstream of GSDMD activation. Science 362, 956–960 (2018).
Gong, Y. N. et al. ESCRT-III acts downstream of MLKL to regulate necroptotic cell death and its consequences. Cell 169, 286–300 (2017).
Boucher, D. et al. Caspase-1 self-cleavage is an intrinsic mechanism to terminate inflammasome activity. J. Exp. Med. 215, 827–840 (2018).
Jost, P. J. et al. XIAP discriminates between type I and type II FAS-induced apoptosis. Nature 460, 1035–1039 (2009).
Halle, S. et al. In vivo killing capacity of cytotoxic T cells is limited and involves dynamic interactions and T cell cooperativity. Immunity 44, 233–245 (2016).
Keefe, D. et al. Perforin triggers a plasma membrane-repair response that facilitates CTL induction of apoptosis. Immunity 23, 249–262 (2005).
Thiery, J. et al. Perforin activates clathrin- and dynamin-dependent endocytosis, which is required for plasma membrane repair and delivery of granzyme B for granzyme-mediated apoptosis. Blood 115, 1582–1593 (2010).
Jorgensen, I., Zhang, Y., Krantz, B. A. & Miao, E. A. Pyroptosis triggers pore-induced intracellular traps (PITs) that capture bacteria and lead to their clearance by efferocytosis. J. Exp. Med. 213, 2113–2128 (2016).
Mariathasan, S. et al. Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 430, 213–218 (2004).
Kuida, K. et al. Altered cytokine export and apoptosis in mice deficient in interleukin-1 beta converting enzyme. Science 267, 2000–2003 (1995).
Zheng, T. S., Hunot, S., Kuida, K. & Flavell, R. A. Caspase knockouts: matters of life and death. Cell Death Differ. 6, 1043–1053 (1999).
Miao, E. A. et al. Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat. Immunol. 11, 1136–1142 (2010).
Wheat, R. L., Clark, P. Y. & Brown, M. G. Quantitative measurement of infectious murine cytomegalovirus genomes in real-time PCR. J. Virol. Methods 112, 107–113 (2003).
Barthel, M. et al. Pretreatment of mice with streptomycin provides a Salmonella enterica serovar Typhimurium colitis model that allows analysis of both pathogen and host. Infect. Immun. 71, 2839–2858 (2003).
Butler, S. L. et al. The antigen for Hep Par 1 antibody is the urea cycle enzyme carbamoyl phosphate synthetase 1. Lab. Invest. 88, 78–88 (2008).
Sato, T. et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459, 262–265 (2009).
Miyoshi, H. & Stappenbeck, T. S. In vitro expansion and genetic modification of gastrointestinal stem cells in spheroid culture. Nat. Protoc. 8, 2471–2482 (2013).
von Moltke, J. et al. Rapid induction of inflammatory lipid mediators by the inflammasome in vivo. Nature 490, 107–111 (2012).
Gregory, S. H., Jiang, X. & Wing, E. J. Lymphokine-activated killer cells lyse Listeria-infected hepatocytes and produce elevated quantities of interferon-γ. J. Infect. Dis. 174, 1073–1079 (1996).
Badovinac, V. P. & Harty, J. T. Adaptive immunity and enhanced CD8+ T cell response to Listeria monocytogenes in the absence of perforin and IFN-γ. J. Immunol. 164, 6444–6452 (2000).
Ahmed, R., Salmi, A., Butler, L. D., Chiller, J. M. & Oldstone, M. B. Selection of genetic variants of lymphocytic choriomeningitis virus in spleens of persistently infected mice. Role in suppression of cytotoxic T lymphocyte response and viral persistence. J. Exp. Med. 160, 521–540 (1984).
Cooper, H. S., Murthy, S. N., Shah, R. S. & Sedergran, D. J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Invest. 69, 238–249 (1993).
Acknowledgements
We thank R. Flavell, V. Dixit, R. Vance and M. Deshmukh for sharing mice directly or through The Jackson Laboratory. This work was supported by the following NIH grants: AI097518, AI133236, AI139304, AI119073, AI136920 (E.A.M.) AI097518-02S1 (V.I.M.), R56AI110682 and R01AI074862 (J.K.W.), R01AI050072 (M.G.B.) and P30CA016086 (UNC Flow Core). We also thank T. Abele, F. Lakhani, M. Artunduaga, M. DePrizio, S. Pereles, A. Vaidyanathan, R. Eplett, M. Mann, S. Redecke and L. Scarpelli for mouse colony upkeep.
Author information
Authors and Affiliations
Contributions
K.N. led the IEC project, V.I.M. led the NK cell and CTL project, and K.N. led the ASM aspects of both projects. E.A.M. supervised. K.N. and E.A.M. discovered the IEC extrusion defect and the ASM mechanism for the IEC and the NK cell and CTL models, and thereby unified the projects. V.I.M. and E.A.M. discovered the NK cell and CTL phenotypes. M.R. and E.A.M. initiated the project. In the IEC project, K.N. performed most of the experiments, with M.R. and A.L.T. performing some experiments. In the NK cell and CTL project, V.I.M. performed most of the experiments; K.N. performed ASM experiments; and J.E.M., C.A.L., C.K.H., L.L. and W.T.N. performed some experiments. H.N.L. and B.D.M. managed the mouse colony. N.J.M., M.G.B., J.K.W. and E.A.M. supervised the work. K.N., V.I.M. and E.A.M. wrote the paper. K.N. and E.A.M. oversaw the completion of the paper.
Corresponding author
Ethics declarations
Competing interests
C.A.L. is employed by AbbVie. This article is composed of the authors' work and ideas and does not reflect the ideas of AbbVie.
Peer review
Peer review information
Nature thanks Jonathan Kagan and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
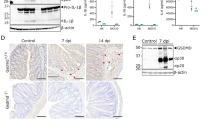
Extended Data Fig. 1 Caspase-7 is required for efficient IEC extrusion during S. Typhimurium infection.
a-b, Caspase-7 and caspase-3 mRNA levels in various tissues from n = 2 samples were examined using BioGPS (biogps.org)6. c, Caspase-7, caspase-3, caspase-1 and caspase-11 expression levels in isolated IEC from n = 6 samples were determined by from published transcriptome data7. d-e, Representative image (d) of cleaved caspase-7 staining of caecum from WT mice 6 h post-infection (hpi) with 106 S. Typhimurium and its quantification with uninfected control (uninfected mice (n = 6) and infected mice (n = 6)) (e). f, Cleaved caspase-7 staining of caecum from WT mice 12 hpi with 106 GFP-S. Typhimurium. g-h, Representative image (g) of cleaved caspase-3 staining of caecum from WT mice 6 hpi with 106 S. Typhimurium and its quantification with uninfected control (uninfected mice (n = 4) and infected mice (n = 6)) (h). I, Quantitation in as in (d) for littermate-controlled Casp3+/– (n = 8) and Casp3–/– mice (n = 6). j, EpCAM staining of caecum from Casp7–/–mice 24 hpi with 5x106 S. Typhimurium (→, extrusion site with 18 clustered cells was one of the largest observed, related to Fig. 1a,b). k-l, quantitation of EpCAM+ cells per extruding site in caecum from littermate Casp7+/– and Casp7–/– mice 6 hpi (k) or 15 hpi (l) with 5x106 S. Typhimurium; from the same experiment as the 24 hpi time point in Fig. 1b. Data are representative of 2 experiments (e, h, i), 3 experiments (d, f, g) or 1 experiment (a–c, k, l). Scale bar = 50 μm. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test). Data are shown as median ± SEM. Exact p values in Source Data EDF1
Extended Data Fig. 2 Caspase-7 facilitates IEC extrusion after FlaTox treatment.
a, Representative images of WT and Casp7–/– organoids after FlaTox treatment in 12 pooled experiments. b, Percentage of ruptured WT and Casp7–/– organoids after PBS treatment in pooled live imaging experiments. c-d, Percentage of ruptured WT and Casp3–/– organoids after FlaTox (c) or PBS (d) treatment in pooled experiments. e, Representative images of indicated organoids 30 min after FlaTox treatment, stained with phalloidin and for cleaved caspase-7 (Related to Fig. 1d). f, Immunoblot of IEC organoids treated with FlaTox probed for the indicated caspases. Data are representative of 3 experiments (e, f) or pooled from 12 (a), or 3 (b–d) experiments. Scale bar = 20 μm. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided unpaired t-test). Data are shown as mean ± SEM. Exact p values available in Source Data EDF2
Extended Data Fig. 3 Caspase-7 prolongs membrane integrity against gasdermin D pores.
a, Gasdermin D (GSDMD) cleavage in organoids treated with FlaTox. b, Representative images in live-cell imaging showing PI intensity of WT and Casp7–/– organoids treated with FlaTox. c-d, Quantitation of PI intensity in live-cell imaging of WT and Casp7–/– organoids treated with PBS (c) or TNF + CHX or PBS control (d). e-f, Representative images (e) and quantitation (f) in live-cell imaging of calcein intensity of WT and Casp7–/– organoids treated with FlaTox. g-h, Quantitation in live-cell imaging of PI intensity of WT and Casp3–/– organoids treated with FlaTox (g) or PBS (h). Data are representative of 3 experiments. Scale bar = 50 μm. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-way ANOVA with Sidak’s post-hoc test). Data shown as mean ± SEM. Exact p values in Source Data EDF3
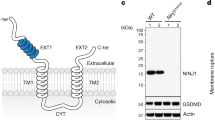
Extended Data Fig. 4 ASM cleavage by caspase-7 requires gasdermin D.
a, Validation of ASM antibody performed by immunoblot of Crispr/CAS9 targeted HeLa cells. b–d, ASM cleavage of organoids that were removed of dead cells and stimulated 20 min with FlaTox. (b) organoid cells were split into 3 tubes, one for T = 0 without stimulation (left blots, shorter 30 s exposure and longer 2 min exposure) and two for 20 min PBS or FlaTox treatment (right blot, shorter 30 s exposure). (c) ASM cleavage from WT, Casp3–/–, or Nlrc4–/– organoids. (d) WT organoid lysates treated with FlaTox in (b) was mock or treated with PNGase F to remove glycosylation; a band shift excludes the possibility that the 57-kDa band represents a deglycosylation event of pro-ASM. Data are representative of 3 experiments.
Extended Data Fig. 5 Caspase-7 activates ASM to generate ceramide.
a, Established mechanism whereby sphingomyelin is cleaved by ASM to generate ceramide, a lipid that naturally invaginates membranes to drive membrane repair via endocytosis (adapted from (Andrews et al, 2014)10). b-c, Ceramide specificity of anti-ceramide antibodies was validated by treating HeLa cells with exogenous C-16 ceramide for 2.5 h followed by staining with the anti-ceramide antibodies 15B4 (b) or MAB_0014 (c). d, Ceramide staining of WT organoids at 20 min post PBS or FlaTox. e, Ceramide staining of organoids at 20 min post FlaTox, with inset expanded images of the boxed areas. f, Ceramide and cleaved caspase-7 staining of indicated organoids at 20 min post FlaTox. g-i, Live imaging of indicated organoids after FlaTox treatment, quantitated for rupture percentage (g), calcein intensity (h), or extrusion starting time (i). j-l, Live imaging of indicated Casp7–/– + dimethylformamide (DMF) or vehicle or Casp7–/– + ceramide organoids after FlaTox treatment with quantitation of PI intensity (j), extrusion starting time (k), or rupture percentage (l). m, Cleaved caspase-7 staining of caecal tissues from IMP-treated WT mice 24 hpi with 5x106 S. Typhimurium, related to Fig. 2d (> indicates cleaved caspase-7+ cells that appear stuck in the monolayer and lack normal extrusion morphology). n, Cleaved caspase-7 staining of WT and WT+IMP organoids at 30 min post FlaTox. Data are representative of 2 experiments (b, c) or 3 experiments (d–f, j, m-n), or are pooled from 3 experiments (h, l). For (g), live Imaging of WT (n = 6 datasets), WT+IMP (n = 5 datasets), and Casp7–/– (n = 3 datasets) organoids pooled from 7 experiments were analysed. For (i), WT (n = 28) and Casp3–/– (n = 27) organoids pooled from 3 experiments were analysed. For (k), Casp7–/– + DMF (n = 21) and Casp7–/– + ceramide (n = 23) organoids pooled from 3 experiments were analysed. Scale bar = 20 μm (b–d, f, n), 50 μm (e, m). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided unpaired t-test in (g, i, k, l), two-way ANOVA with Tukey’s post-hoc test in (h) or with Sidak’s post-hoc test in (j)). Data are shown as mean ± SEM. Exact p values in Source Data EDF5
Extended Data Fig. 6 Ceramide production by caspase-7 requires gasdermin D pores.
a–c, Ceramide and EpCAM staining of caecum from WT (a), Gsdmd–/– (b), or WT+IMP (c) mice 24 hpi with 5x106 S. Typhimurium (→, extruding or extruded cells). Scale bar = 50 μm. Data are representative of 3 experiments.
Extended Data Fig. 7 ASM cleavage is requires for IEC extrusion.
a, Strategy to generate ASM D249A mutant mice by CRISPR–Cas9. Target sequence for guide RNA in exon 2 is shown in red. Repair oligo DNAs (200 nt) containing indicated mutation are also used for electroporation with Cas9. Successful mutation was confirmed by Sanger sequencing. b, Quantification of EpCAM+ cells per extruding site in non-infected WT, Smpd1DA/DA, and Casp7–/– caeca (n = 4 in each group). c–f, Live imaging of indicated IEC organoids, which were quantified for rupture percentage after PBS (c) or FlaTox (d) treatment, extrusion starting time after FlaTox treatment (e), and PI Intensity after TNF+CHX or PBS treatment (f). Data are pooled from 2 (b) or 3 (c-d) experiments or are representative of 3 experiments (f). For (e), WT (n = 27) and Smpd1DA/DA (n = 41) organoids pooled from 3 experiments were analysed. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided unpaired t-test in (d, e), two-way ANOVA with Sidak’s post-hoc test in (f)). Data are shown as mean ± SEM. Exact p values in Source Data EDF7
Extended Data Fig. 8 Caspase-7 ameliorates tissue pathology during S. Typhimurium infection or DSS colitis.
a–c, The indicated mice were infected with 106 S. Typhimurium for 2 days, then caeca were collected and stained with H&E. Shown are representative images (a) (Scale bar = 100 μm), pathological score (b) of WT (n = 11), Casp7–/– (n = 9), and Casp1–/–Casp11–/– (n = 7) mice, and serum IFNγ (c) of WT, Casp7–/–, and Casp1–/–Casp11–/– mice (n = 6). d, EpCAM staining of caeca from littermate Casp7+/– and Casp7–/– mice 2 dpi with 5x106 S. Typhimurium. Dotted lines indicate the space between the extruding IECs and epithelial layer. Scale bar = 50 μm. e–g, CFU of WT (n = 31), Casp7–/– (n = 27), and Casp1–/–Casp11–/– (n = 18) mice 4 dpi with 106 S. Typhimurium in spleen (e), MLN (f), and gentamicin-treated caecum (g). h–j, CFU of WT (n = 8) and Smpd1DA/DA (n = 6) mice 4 dpi with 106 S. Typhimurium in (h) spleen, (i) MLN, and (j) gentamicin-treated caecum. k-l, Casp7+/– and Casp7–/– mice treated with the indicated time course of DSS. Mouse weight (k) and pathology score (l) at day 5. Data are representative of 2 experiments with (a, b, d, k, l), or are pooled from 2 experiments (c), 3 experiments (h–j), or 5 experiments (e-g). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test in (b), two-sided unpaired t-test in (c, h–j), one-way ANOVA with Dunnett’s post-hoc test in (e–g), two-way ANOVA with Sidak’s post-hoc test in (k)). Data are shown as mean ± SEM. Exact p values in Source Data EDF8
Extended Data Fig. 9 Caspase-7 and perforin phenocopy defence against C. violaceum infection.
a-b, Mice were infected IP with 104 C. violaceum and bacterial burdens in the liver were determined at 3 dpi for separately bred mice (a) or littermate-controlled mice (b). c, Ly5A+ splenocytes were collected from wild-type mice and expanded ex vivo in IL-2 for the NK adoptive transfer experiments in Fig. 4d,e. Shown is the percentage of transferred cells that were NK cells. d, NK cell adoptive transfer schematic for experiments in Fig. 4d,e. Data are pooled from 2 experiments in (a), representative of two experiments in (b), and representative of 3 experiments in (c). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (One-way ANOVA (a) or two-sided Mann–Whitney U-test (b)). Bars indicate mean with standard deviation (c). Exact p and n values in Source Data EDF9
Extended Data Fig. 10 IL-18 primes NK cell cytotoxicity to activate caspase-7 in hepatocytes.
a-b, e–g, Casp1-11DKO mice were infected with 104 C. violaceum, treated with PBS or IL-18 (IP 0.2 μg recombinant mouse IL-18 (MBL) at day 0 and 1), and collected at 2 dpi. Livers were stained for the indicated markers via immunofluorescence. Single-channel zoom images are from inset box. Merged inset is shown in main Fig. 4f. Scale bars, 50 μm. c, Quantification of cleaved caspase-7 signal within lesions, with each dot as the average per mouse. d, Percent of cleaved caspase-7-positive cells that co-stain with CPS1. g–j, Casp7–/– or WT mice were left uninfected or infected with 104 C. violaceum as indicated, and collected at 2 dpi. Representative images of livers stained for nuclei (DAPI; blue), cleaved caspase-7 (g-h, j, green), cleaved caspase-3 (i, red), TUNEL (j, red). k-l, WT mice were infected with 104 C. violaceum and collected at 3 dpi, then stained for cleaved caspase-7 (k, white), cleaved PARP (l, white). Scale bars, 50 μm. Data are pooled (c-d) or representative (a-b, e–l) of 2 experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test). Bars indicate median values. Exact p and n values in Source Data EDF10
Extended Data Fig. 11 Caspase-7 enhances ceramide production through ASM after NK–CTL attack.
a–c, Mice were treated with IMP or PBS, followed by intraperitoneal infection with 104 C. violaceum and collected at 3 dpi. Representative images of infected livers from WT (a), Casp7–/– (b), or WT+IMP (c) mice stained for nuclei (DAPI; blue), cleaved caspase-7 (white), and ceramide (red). Scale bars, 50 μm. Data are representative of 3 experiments.
Extended Data Fig. 12 CTL transfer model during L. monocytogenes infection.
a, Percentage of bacterial burden localized to hepatocytes at 3 dpi. Equal weight liver sections were removed and the hepatocyte fraction was graphed as a percentage of the total burden by weight. 3 mice per genotype. b-c, Mice were infected IV with 5x103 L. monocytogenes and IP treated with combinations of isotype antibody or NK depleting antibody (100 μg anti-NK1.1 PK136), with IL-18 (0.2 μg recombinant mouse IL-18) or isotype control as indicated. Mouse numbers (b) n = 5 each group; (c) WT PBS n = 6, WT IL-18 n = 6, Casp7–/– n = 7 each. d, Timeline for adoptive transfers. Donor mice were PBS treated (naïve) or vaccinated with 1x106 ∆actA L. monocytogenes (immune) in Fig. 5. e-f, Liver and splenic burdens 8 days post primary infection with 5x103 L. monocytogenes. n = 6 mice each. g, Flow gating strategy for flow experiments depicted in (h-i). h-i, Donor mice were NK depleted (> 99% eliminated) and their CTLs enriched by negative selection (> 72% purity). j, Mice were adoptively transferred with the indicated numbers of purified CTLs (naïve were given 1x107 as the maximum transferred number), infected with 5x104 L. monocytogenes and bacterial burdens were analysed in the liver and spleen at 3 dpi. 2 naïve mice, 3 per immunized group. k, Bulk splenocyte transfer (5x107, as previously described in23) with NK depletion was comparable to that of isolated CTLs. Naïve n = 3, immunized n = 4 mice. l, Mice were infected with 5x104 of L. monocytogenes and adoptively transferred with immunized WT CTLs as in d. Liver bacterial burdens were determined at 3 dpi. Mouse numbers: Casp7+/– (n = 2F), Casp7–/– (n = 3M+2F), and Casp6–/– (n = 5F+3M). Note full clearance in Casp7+/– mice may be due to gender disparities or simply to low numbers leading to stochasticity and poor sampling. m-n, Donor mice were PBS treated (naïve) or vaccinated with 1x106 ∆actA L. monocytogenes (immune). (m) Adoptive transfer data from Fig. 5b,c are shown on the same graph, as all the transfers were done in the same 2 pooled experiments. Numbers of mice (m) WT mice with Casp7–/– CTLs n = 7 each, naïve Prf1–/– CTLs n = 8, immune Prf1–/– CTLs n = 7; naïve knockout recipients n = 7 each, immune recipients Prf1–/– n = 8, Casp7–/– n = 7; (n) naïve WT n = 8, immune WT n = 10, Ifng–/– n = 9 each. Data are representative of two experiments (a, e-f, h) or pooled from two experiments (b-c, j–n). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test (a–c, k, l), or (j, m, n) one–way ANOVA with Tukey’s multiple comparison test). Bars indicate mean with standard deviation (a). Box plots show median, 25th–75th percentile, whiskers are minimum and maximum. Exact p and n values in Source Data EDF12
Extended Data Fig. 13 CTL transfer induces cleavage of caspase-7 during L. monocytogenes infection.
a, Timeline for adoptive transfers with recipients infected IV with 5x104 L. monocytogenes in Extended Data Fig. 13. b-c, Flow cytometry of enriched hepatocytes for cleaved caspase-7 staining and gating scheme, with representative plot (b) and quantification of n = 6 mice per group (c). d, Representative images of livers stained for nuclei (DAPI; blue), cleaved caspase-7 (green), and CPS1 (red). Scale bars, 50 μm. e-f, Immunofluorescence control staining. Casp7–/– mice were left uninfected (e) or CTL transferred and infected with 5x104 L. monocytogenes (f), and collected at 3 dpi. Representative images of livers stained for nuclei (DAPI; blue) and cleaved caspase-7 (green). Scale bars, 50 μm. g, Quantification of cleaved caspase-7 signal within lesions (dot = average per mouse; n = 7 WT mice per group). h-i, Colocalization of cleaved caspase-7 signal and L. monocytogenes with representative image (h) and quantification (i) from n = 6 naïve and n = 5 immune WT mice. Scale bar, 50 μm. Graphed data are pooled from 2 experiments (c, g, i). Images representative of 2 experiments (d, h) or one experiment (e, f). * P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test). Bars indicate median values. Exact p and n values in Source Data EDF13
Extended Data Fig. 14 MCMV and LCMV are cleared independent of caspase-6 and caspase-7.
a-b, Mice were infected IP with 5x104 PFU of MCMV. Viral titre in the liver was determined at 4 dpi from (a) n = 6 WT, n = 9 Casp7–/–, n = 8 Prf1–/–; (b) n = 10 WT, n = 11 Casp6–/–, n = 11 Prf1–/–. c-d, Mice were infected IP with 2x105 PFU of LCMV. Viral titre in the liver was determined by plaque assay at 8 dpi from (c) n = 5 WT, n = 6 Casp7–/–, n = 6 Prf1–/–; (d) n = 8 WT, n = 9 Casp6–/–, n = 5 Prf1–/–. All data are pooled from 2 experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test). Bars indicate median values. Exact p and n values in Source Data EDF14
Extended Data Fig. 15 Granzyme B, active caspase-3 and active caspase-7 are not sufficient to kill bacteria in vitro.
a–d, NK co-culture killing assays (see “In vitro co-culture assays” in methods for detailed description) with C. violaceum-infected YAC-1 cells or L. monocytogenes-infected Hepa1-6 and YAC-1 cells as indicated. (a, c) Western blot analysis of cleaved caspase-3 (CC3) and/or cleaved caspase-7 (CC7). (b, d) Bacterial counts 5 h post co-culture. e-f, Hepa1-6 cell lysates were spiked with granzyme B at the indicated amounts for 1 h and 8x106 L. monocytogenes was added. Western blot analysis of CC3 and CC7 (e) and time course of bacterial numbers (f). g, Invasion assay with L. monocytogenes incubated in Hepa1-6 cell lysates for 16 h with granzyme B. All data are representative of 2 experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test). Bars indicate mean
Extended Data Fig. 16 Pyroptotic activation of caspase-7 is not required for clearance of C. violaceum.
a–c, Mice were infected IP with 1x104 C. violaceum and monitored for survival (a) or collected for bacterial counts at 2 dpi (b-c). d–h, Mice were infected with C. violaceum and spleen burdens determined 3 dpi; spleen burdens shown are from the same mice whose liver burdens are shown in Extended Data Fig. 9a, Fig. 4a,b, and Fig. 4d,e. i–k, Mice were infected IP with B. thailandensis at 2x107 (i) or 1x104 CFUs (j-k) and mice were monitored for survival (i) or bacterial burdens were enumerated 3 dpi (j-k). l-m, Mice were infected IP with 1x105 total S. typhimurium (5x104 of WT plus 5x104 of FliCON), with bacterial burdens determined 2 dpi and burdens graphed as competitive index (CI) of FliCON to WT bacteria (l) or graphed as total burdens (m). n, Liver bacterial burdens 3 dpi with 5x104 L. monocytogenes, with adoptive transfer schematic. Donor mice were PBS treated (naïve) or vaccinated with 1x106 ΔactA L. monocytogenes (immune). O, Presence or absence of Smpd1 (encoding ASM), Casp7, Prf1, and Gsdmd in the indicated taxonomic groups were determined by gene annotation and verified by reciprocal BLAST searches against the respective mouse gene product. Data are pooled from three experiments (a–c), pooled from two experiments (d–h, l–n) or representative of two experiments (i–k). Mouse numbers (a) n = 9 WT, n = 15 Gsdmd–/–; (b-c) n = 12 WT, n = 11 Casp1–/–Casp11–/–, n = 11 Gsdmd–/–; (i) n = 5 WT, n = 3 Casp1–/–Casp11–/–, n = 5 Casp7–/–; (j–k) n = 3 WT, n = 4 Casp7–/–, n = 5 Casp1–/–Casp11–/–; (l–m) n = 9 WT, n = 9 Casp7–/–, n = 6 Casp1–/–Casp11–/–; (n) n = 6 naïve Casp1–/–Casp11–/–, n = 7 immune Casp1–/–Casp11–/–, n = 7 each Casp7–/–. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (Two-sided Mann–Whitney U-test, or (a, i) log-rank Mantel–Cox test). Bars indicate median values. Box plots show median, 25th–75th percentile, whiskers are minimum and maximum. Exact p and n values in Source Data EDF16
Extended Data Fig. 17 Model for membrane repair driven by caspase-7.
a, IECs detect the activity of the Salmonella T3SS via NLRC4 and activate caspase-1 in response. Caspase-1 then activates both the gasdermin D pore as well as caspase-7. We propose a model in which caspase-7 diffuses through the gasdermin D pore, thereby entering the extracellular space. Simultaneously, the gasdermin D pore triggers lysosomal exocytosis, thus delivering ASM to the extracellular space. In this model, caspase-7 and ASM meet in the extracellular space. Note that additional studies will be needed to validate the location of the caspase-7 and ASM interaction. Caspase-7 cleaves ASM, increasing its enzymatic activity and generating more ceramide. This ceramide can then be used for continuous endocytic repair of gasdermin D pores to facilitate IEC extrusion. b, NK cells or CTLs attack by degranulating perforin and granzyme B. Perforin pores allow granzyme B to enter the target cell, where it activates caspase-3 (not shown) and caspase-7. Activated caspase-7 can exit the cell to encounter ASM, which generates ceramide that should allow endocytosis of perforin pores. We propose that this maintains cell integrity long enough for the cell to complete caspase-3-driven apoptosis.
Supplementary information
Supplementary Figures
This file contains the uncropped western blots.
Video 1
WT IEC organoid treated with FlaTox, related to Extended Data Fig. 2a. Time-lapse imaging of single WT IEC organoid treated with FlaTox, still images are shown in Extended Data Fig. 2a. This is a representative video from 12 live imaging experiments; each experiment contains 10–20 videos. The pooled data is graphed in Figure 1c. Scale bar = 50 μm.
Video 2
Casp7–/– IEC organoid treated with FlaTox, related to Extended Data Fig. 2a. Time-lapse imaging of single Casp7–/– IEC organoid treated with FlaTox, still images are shown in Extended Data Fig. 2a. This is a representative video from 12 live imaging experiments; each experiment contains 10-20 videos. The pooled data is graphed in Figure 1c. Scale bar = 50 μm.
Video 3
PI uptake in a WT organoid treated with FlaTox, related to Figure 1e and Extended Data Fig.3b. Time-lapse imaging shows the PI uptake in a WT IEC organoid treated with FlaTox, still images are shown in Figure 1e and Extended Data Fig. 3b. The same video repeats twice, first showing DIC+PI, and the second showing only PI. This is a representative video from 30 videos of 3 live imaging experiments. The data is graphed in Figure 1f. Scale bar = 50 μm.
Video 4
PI uptake in a Casp7–/– organoid treated with FlaTox, related to Figure 1e and Extended Data Fig.3b. Time-lapse imaging shows the PI uptake in a Casp7–/– IEC organoid treated with FlaTox, still images are shown in Figure 1e and Extended Data Fig. 3b. The same video repeats twice, first showing DIC+PI, and the second showing only PI. This is a representative video from 26 videos of 3 live imaging experiments. The data is graphed in Figure 1f. Scale bar = 50 μm.
Video 5
Calcein egress from a WT organoid with FlaTox, related to Extended Data Fig. 3e. Time-lapse imaging shows calcein egress from a WT IEC organoid treated with FlaTox, still images are shown in Extended Data Fig. 3e. This is a representative video from 18 videos of 3 live imaging experiments. The data is graphed in Extended Data Fig. 3f. Scale bar = 50 μm.
Video 6
Calcein egress from a Casp7–/– organoid with FlaTox, related to Extended Data Fig. 3e. Time-lapse imaging shows calcein egress from a Casp7–/– IEC organoid treated with FlaTox, still images are shown in Extended Data Fig. 3e. This is a representative video from 17 videos of 3 live imaging experiments. The data is graphed in Extended Data Fig. 3f. Scale bar = 50 μm.
Video 7
PI uptake in a WT organoid treated with FlaTox at high frequency, related to Extended Data Fig. 3b. Time-lapse imaging shows the PI uptake in a WT IEC organoid treated with FlaTox at higher frequency, related to Extended Data Fig. 3b. DIC images and PI images were merged. This is a representative video from 10 videos of 2 live imaging experiments. Scale bar = 50 μm.
Video 8
PI uptake in a Casp7–/– organoid treated with FlaTox at high frequency, related to Extended Data Fig. 3b. Time-lapse imaging shows the PI uptake in a Casp7–/– IEC organoid treated with FlaTox at higher frequency, related to Extended Data Fig. 3b. DIC images and PI images were merged. This is a representative video from 12 videos of 2 live imaging experiments. Scale bar = 50 μm.
Video 9
PI uptake in a Smpd1DA/DA organoid treated with FlaTox at high frequency, related to Figure 3d. Time-lapse imaging shows the PI uptake in a Smpd1DA/DA IEC organoid treated with FlaTox at higher frequency, related to Figure 3d. DIC images and PI images were merged. This is a representative video from 10 videos of 2 live imaging experiments. Scale bar = 50 μm.
Source data
Rights and permissions
About this article

Cite this article
Nozaki, K., Maltez, V.I., Rayamajhi, M. et al. Caspase-7 activates ASM to repair gasdermin and perforin pores. Nature 606, 960–967 (2022). https://doi.org/10.1038/s41586-022-04825-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-04825-8
This article is cited by
-
DHX9 maintains epithelial homeostasis by restraining R-loop-mediated genomic instability in intestinal stem cells
Nature Communications (2024)
-
The gasdermin family: emerging therapeutic targets in diseases
Signal Transduction and Targeted Therapy (2024)
-
An innate granuloma eradicates an environmental pathogen using Gsdmd and Nos2
Nature Communications (2023)
-
Pore-forming proteins as drivers of membrane permeabilization in cell death pathways
Nature Reviews Molecular Cell Biology (2023)
-
Inflammasome activation occurs in CD4+ and CD8+ T cells during graft-versus-host disease
Cell Death & Disease (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
