
Abstract
Comprehensive assessments of species’ extinction risks have documented the extinction crisis1 and underpinned strategies for reducing those risks2. Global assessments reveal that, among tetrapods, 40.7% of amphibians, 25.4% of mammals and 13.6% of birds are threatened with extinction3. Because global assessments have been lacking, reptiles have been omitted from conservation-prioritization analyses that encompass other tetrapods4,5,6,7. Reptiles are unusually diverse in arid regions, suggesting that they may have different conservation needs6. Here we provide a comprehensive extinction-risk assessment of reptiles and show that at least 1,829 out of 10,196 species (21.1%) are threatened—confirming a previous extrapolation8 and representing 15.6 billion years of phylogenetic diversity. Reptiles are threatened by the same major factors that threaten other tetrapods—agriculture, logging, urban development and invasive species—although the threat posed by climate change remains uncertain. Reptiles inhabiting forests, where these threats are strongest, are more threatened than those in arid habitats, contrary to our prediction. Birds, mammals and amphibians are unexpectedly good surrogates for the conservation of reptiles, although threatened reptiles with the smallest ranges tend to be isolated from other threatened tetrapods. Although some reptiles—including most species of crocodiles and turtles—require urgent, targeted action to prevent extinctions, efforts to protect other tetrapods, such as habitat preservation and control of trade and invasive species, will probably also benefit many reptiles.
Similar content being viewed by others
Main
Although comprehensive extinction-risk assessments have been available for birds, mammals and amphibians for well over a decade3, reptiles have, until now, not been comprehensively assessed. Therefore, conservation science and practice has typically relied on the International Union for Conservation of Nature (IUCN) Red List categories and distributions of the other three tetrapod classes to inform policy and guide priorities for investments2, despite differing expectations as to how effective common strategies will be across classes9,10. With a high diversity in arid regions and some islands and archipelagos (for example, Antilles, New Caledonia and New Zealand) compared with other tetrapods, reptiles were thought to require different conservation strategies and geographical priorities6. In the absence of Red List assessments, researchers have resorted to indirect measures of extinction risk such as range size and human pressure6,11. Here we examine the results of a comprehensive Red List assessment of reptiles and outline their implications for the conservation needs of reptiles.
Comprising the turtles (Testudines), crocodiles (Crocodylia), squamates (Squamata: lizards, snakes and amphisbaenians) and tuatara (Rhynchocephalia), reptiles are a paraphyletic class representing diverse body forms, habitat affinities and functional roles in their respective ecosystems12. The largely terrestrial squamates are by far the most speciose group (9,820 species in this assessment), whereas the primarily aquatic turtles and crocodiles are often larger bodied but include only 351 and 24 species, respectively. Rhynchocephalians diverged from the snake and lizard lineage in the Triassic period and include one extant species13. Given this diversity of reptiles, threats to their persistence are likely to be equally varied, and so these need to be specified to guide effective conservation action.
Extinction risk and threats
We assessed reptiles globally using the IUCN Red List criteria with input from 961 scientists (Supplementary Note 1) achieved primarily through 48 workshops (Supplementary Table 1). Across all 10,196 species assessed, 21.1% are threatened with extinction (categorized as vulnerable, endangered or critically endangered; Supplementary Table 2). As a group, a greater number of reptile species are threatened than birds or mammals, but fewer than amphibians. Proportionately more mammals and amphibians are threatened than reptiles (Fig. 1a). The reptile threat prevalence falls within a previous estimate of 15–36% threatened (best estimate 19%) from a random sample of 1,500 reptile species8. To our knowledge, this study represents the first global test of a sampled Red List extrapolation. The proportion of turtles and crocodiles that are threatened (57.9% and 50.0%, respectively) is much higher than those of squamates (19.6%) and tuatara (0%), and comparable to the most-threatened tetrapod groups, salamanders (57.0%) and monotremes (60.0%) (Fig. 1b). Within squamates, iguanid (73.8%) and xenosaurid (60.0%) lizards and uropeltid (61.1%) and tropidophiid (60.0%) snakes are highly threatened. Since 1500, 31 reptile species (0.3%) have been driven extinct, including 24 squamates and 7 turtles, with 2 squamate species from Christmas Island categorized as extinct in the wild (persisting only as captive populations); 40 critically endangered species are ‘possibly extinct’ (that is, species that are likely to be extinct, but that have a small chance that they may be extant; Extended Data Fig. 1). Additional species probably became extinct before being documented by science14.

a, Taxonomic patterns organized by class. The numbers above each column refer to the numbers and percentages of species threatened (that is, those categorized as critically endangered, endangered or vulnerable). b, Extinction risk by major taxonomic groups. Blue lines indicate the best estimate of the percentage of species threatened. CR, critically endangered; DD, data deficient; EN, endangered; EW, extinct in the wild; EX, extinct; LC, least concern; NT, near threatened; VU, vulnerable.
Limited information resulted in 1,507 species (14.8%) being categorized as data deficient, similar to the number in mammals (15.1%) and lower than for amphibians (20.4%), but much higher than for birds (0.5%). Taxonomic groups with fossorial or other secretive habits and/or restricted to poorly studied areas (such as blindsnakes (Gerrhopilidae and Typhlopidae)) had greater proportions of species categorized as data deficient (Supplementary Table 2). The greatest numbers of data-deficient species occur in Asia (585), South America (284) and Africa (271), with fewer data-deficient species in North and Middle America (163), Oceania (124), Australia (55), the Caribbean (34) and Europe (3). Uncertainty about the status of data-deficient species suggests that the proportion of reptiles threatened with extinction ranges from 18.0% (assuming no data-deficient species are threatened) to 32.8% (assuming all data-deficient species are threatened) with a best estimate of 21.1%.
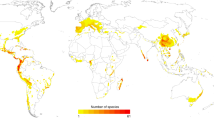
Concentrations of threatened reptiles are mostly in regions in which other tetrapods are also threatened (Extended Data Fig. 2). Threatened reptiles are concentrated in southeastern Asia, West Africa, northern Madagascar, the northern Andes and the Caribbean, but largely absent from Australian drylands; the Kalahari, Karoo and Sahara deserts; northern Eurasia; and the Rocky Mountains and northern North America (Fig. 2a). In remarkably few regions, however, are reptiles disproportionately threatened relative to other tetrapods (that is, have at least twice the number of species in a threatened category): parts of southern Asia and northeastern USA (Fig. 2b and Extended Data Table 1). Moreover, for most (87%) terrestrial regions in which tetrapods occur, no tetrapod class is disproportionately threatened compared with the other classes.

a, Distribution of reptile species that are threatened (critically endangered, endangered or vulnerable). b, Regions with disproportionate numbers of threatened species for each tetrapod class (areas for each class where the proportional threat in species diversity is at least twice the loss for the next-most threatened class). c, Loss of reptile phylogenetic diversity (PD) if all threatened species became extinct. d, Regions with disproportionate phylogenetic diversity loss for each tetrapod class (calculated as in b). Grey, areas with no threatened species (a, c) or regions in which no class is disproportionately threatened (b, d). Data are shown at a resolution of 50 km.
With deep phylogenetic lineages and high species diversity, reptiles stand to lose a large amount of phylogenetic diversity (a measure of difference within an evolutionary tree15) if the current extinction crisis continues apace. Assuming all threatened species (and only these species) become extinct, the combined loss of reptile phylogenetic diversity (calculated using existing phylogenetic trees16,17) will be approximately 15.6 billion years. Southeastern Asia, India, West Africa and the Caribbean8 (Fig. 2c) comprise the top 15% areas of phylogenetic diversity loss, with high concentrations of threatened and evolutionarily distinct species (Extended Data Fig. 3). Comparing the distributions of threatened phylogenetic diversity across all four tetrapod groups reveal relatively small geographical areas of disproportionate importance for any class (Fig. 2d and Extended Data Table 2).
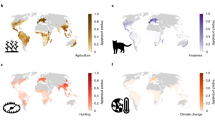
The anthropogenic factors increasing extinction risk in reptiles are mainly habitat destruction from agricultural expansion, urban development and logging (Fig. 3a). Other important threats are invasive species and hunting, which includes commercial harvest and trade (Fig. 3a). Among reptile groups, crocodiles and turtles are most frequently affected by hunting and less by agriculture, whereas squamates are most frequently threatened by agriculture (Fig. 3a). The major threats are broadly similar across tetrapods (Fig. 3b). For all tetrapod groups, agriculture threatens the most species, logging is the second or third most prevalent threat, and invasive species and disease are the fourth or fifth most prevalent threat. Threats causing habitat destruction (complete removal of habitat) affect proportionately more species than those causing habitat change (degradation of habitat). The largest differences in relative threat prevalence are for hunting, which threatens mammals much more than the other tetrapods, and urban development, which affects amphibians, reptiles and mammals more than birds.

a, Crocodiles, lizards (including amphisbaenians), snakes and turtles. b, All tetrapods. Only threats to species categorized as critically endangered, endangered or vulnerable were included. Some species are subject to more than one threat (mean = 2.4; s.d. = 1.3 threats per species).
Climate change is a looming threat to reptiles, for example, by reducing thermally viable windows for foraging18, skewing offspring sex ratios in species that have temperature-dependent sex determination19 and contracting ranges20. Given the Red List three-generation horizon for assessments, the lack of long-term studies limits the documentation of climate change as a near-future threat to reptiles21, in contrast to, for example, birds (Fig. 3b). Disease is documented as a threat for only 11 species of reptiles (<1% of extant, non-data-deficient species), although pathogens such as Ophidiomyces ophiodiicola (which causes snake fungal disease22) pose a potential threat and are little studied outside North America. Intentional use of reptiles (local consumption and trade) is an important threat to reptiles23, and was found to threaten 329 species (3.2%), especially turtles (30.8% of all turtle species).
More than half of all reptile species occur in forested habitats (Fig. 4c). Although some reptiles, particularly lizards, are speciose in arid or seasonally dry habitats such as deserts, grasslands, shrublands and savannahs6,24, these species are less threatened than those occupying forest habitats (13.7% of species restricted to arid habitats versus 26.6% of species restricted to forests; Fisher’s exact test, P = 0.00001; Fig. 4b). The top threats to reptiles—agriculture, urban development and logging—are also the top threats to species inhabiting forested habitats, affecting 65.9%, 34.8% and 27.9% of forest-dwelling threatened reptiles, respectively, helping to explain the higher extinction risk of forest species. Agriculture and logging are significantly more likely to threaten forest-dwelling than non-forest dwelling reptiles (Fisher’s exact test, P = 0.00001), whereas urbanization threatens forest-dwelling no differently than non-forest dwelling reptiles (Fisher’s exact test, P = 0.25). Turtles and crocodiles are much more frequently associated with wetlands than other reptiles (Fig. 4a).

a, Habitats used by crocodiles, lizards (includes amphisbaenians), snakes and turtles. b, Percentage of reptiles using each habitat that are threatened. c, Habitats used by tetrapods. d, Percentage of threatened tetrapod species using each habitat. See Supplementary Table 3 for additional, rarely used habitats not shown here. Artificial habitats are not shown.
Like reptiles, more than twice as many bird and mammal species occur in forests compared with any other habitat type (Fig. 4c). Forests are also the most common habitat for amphibians, although wetlands are important for many species, especially for breeding (Fig. 4c). Also similar to reptiles, the proportions of forest-inhabiting bird, mammal and amphibian species that are threatened are higher than for species that do not inhabit forests (16.7% versus 13.0%, 27.5% versus 20%, and 42.4% versus 34.4%, respectively; Fisher’s exact tests, P = 0.00001). Threat levels for each tetrapod class in arid habitats tend to be lower (less than 23% of species occurring in such regions). Across tetrapods, forests support high diversity and are also subject to widespread threats.
Surrogacy of other tetrapods for reptiles
With numerous threatened tetrapod species (227 birds, 194 mammals, 607 amphibians and 474 reptiles) ranging completely outside formally protected areas, assessing surrogacy is important to gauge the magnitude of efforts needed to conserve these species. We addressed surrogacy using a complementarity representation approach for threatened species, which better addresses the extent to which areas selected for surrogates capture target features than, for example, spatial congruence25. When combined, threatened birds, mammals and amphibians—the tetrapod groups for which comprehensive Red List data were previously available—are good surrogates for the conservation of threatened reptile diversity when prioritizing the richness of rarity-weighted threatened species at both 50-km and 100-km resolution (median species accumulation indices are 0.66 and 0.76, respectively; Extended Data Fig. 4a). Using this same prioritization strategy, birds and mammals individually are reasonable surrogates for reptiles, whereas amphibians are poor surrogates (Extended Data Fig. 4a). By contrast, for a complementarity representation strategy that prioritizes individual threatened species with the smallest ranges, birds, mammals and amphibians are not good surrogates for reptiles (species accumulation indices < 0.40), although combined they are reasonable surrogates at both 50-km and 100-km resolutions (median species accumulation indices are 0.44 and 0.64, respectively; Extended Data Fig. 4b). These results indicate that the smallest-ranged threatened reptiles tend to be isolated from threatened birds, mammals and amphibians. In addition, priority areas for threatened birds and mammals independently, and birds, mammals and amphibians combined, showed high spatial congruence with priority sites for threatened reptiles for both strategies (prioritizing either rarity-weighted threatened species or complementarity representation), although correlations among global portfolios of priority areas were lower (Extended Data Tables 3, 4). Although our results for the smallest ranged threatened species are consistent with previous expectations of low surrogacy9, overall, we found reasonably high surrogacy10.
Discussion
Our discovery of broad similarities in the geography and nature of threats between reptiles and other tetrapods was unexpected given previous arguments about the exceptionalism of reptiles for being particularly diverse in arid habitats6. The implications for tetrapod conservation are that geographical prioritizations previously performed for birds, mammals and amphibians overlap broadly with prioritizations for all except the most range-restricted threatened reptiles. The absence of reptiles in many global conservation prioritization analyses to date is unlikely to have left the class less represented than others. Nevertheless, the low surrogacy value of other tetrapods for reptiles with the most restricted ranges suggests that a case-by-case focus is required for these microendemics. Indeed, the ranges of 31 threatened reptiles do not overlap with the ranges of any other threatened tetrapod (among threatened species, 84 birds, 11 mammals and 7 amphibians are similarly isolated from other threatened tetrapods) (Supplementary Table 3).
Researchers have predicted that reptiles are particularly vulnerable to climate change in tropical biomes26 as well as freshwater27 and arid habitats18, although so far no clear geographical signal in reptile declines due to climate change has been detected28. If such vulnerabilities are found, then—as climate change continues to alter the distributions and extinction risk of species—the surrogacy across tetrapods could unravel with, for example, reptiles in specific habitat types declining swiftly and disproportionately (relative to other tetrapods).
Among the conservation strategies needed to prevent reptile extinction, land protection is critically important to buffer many threatened species from the dual threats of agricultural activities and urban development. The hundreds of threatened reptiles that currently occur completely outside protected areas underscore the need for targeted safeguards of important sites. Beyond place-based strategies, conservation policy and practice must halt unsustainable harvest and stem the spread of invasive disease to prevent many more species from becoming threatened23. Furthermore, introduced mammals to islands threaten 257 reptile species (2.8% of all reptiles), calling for continued campaigns to eradicate introduced mammals in those places.
With a comprehensive, global assessment of the extinction risk of reptile species now available, these data can be incorporated into the toolbox of conservation practice and policy. At the species level, they can serve as the starting point for ‘green status’ (formerly ‘green list’) assessments that define, measure and incentivize species recovery29. More generally, they can be integrated into the calculation of species threat abatement and restoration metrics2, the identification of key biodiversity areas30 and resource allocation using systematic conservation planning31, all of which have primarily been dependent on data from birds, mammals and amphibians among animals to date. Future reassessments will allow reptile data to be included in the Red List Index32, a widely used indicator of biodiversity trends1.
Although efforts aimed at protecting other threatened tetrapods probably benefit many of the 1,829 threatened reptiles—especially forest-dwelling species—conservation investments targeted at uniquely occurring reptiles or those requiring tailored policies must also be implemented to prevent extinction. Encouragingly, the First Draft of the Post-2020 Global Biodiversity Framework33 to be agreed by governments in 2022 explicitly targets safeguarding important sites (target 3), complemented by emergency action for individual threatened species (target 4). This political determination to reverse the slide of species toward extinction bodes well for reptiles.
Methods
We used the IUCN Red List criteria34,35 and methods developed in other global status-assessment efforts36,37 to assess 10,078 reptile species for extinction risk. We additionally include recommended Red List categories for 118 turtle species38, for a total of 10,196 species covered, representing 89% of the 11,341 described reptile species as of August 202039.
Data compilation
We compiled assessment data primarily through regional in-person and remote (that is, through phone and email) workshops with species experts (9,536 species) and consultation with IUCN Species Survival Commission Specialist Groups and stand-alone Red List Authorities (442 species, primarily marine turtles, terrestrial and freshwater turtles, iguanas, sea snakes, mainland African chameleons and crocodiles). We conducted 48 workshops between 2004 and 2019 (Supplementary Table 1). Workshop participants provided information to complete the required species assessment fields (geographical distribution, population abundance and trends, habitat and ecological requirements, threats, use and trade, literature) and draw a distribution map. We then applied the Red List criteria34 to this information to assign a Red List category: extinct, extinct in the wild, critically endangered, endangered, vulnerable, near threatened, least concern and data deficient. Threatened species are those categorized as critically endangered, endangered and vulnerable.
Taxonomy
We used The Reptile Database39 as a taxonomic standard, diverging only to follow well-justified taxonomic standards from the IUCN Species Survival Commission40. We could not revisit new descriptions for most regions after the end of the original assessment, so the final species list is not fully consistent with any single release of The Reptile Database.
Distribution maps
Where data allowed, we developed distribution maps in Esri shapefile format using the IUCN mapping guidelines41 (1,003 species). These maps are typically broad polygons that encompass all known localities, with provisions made to show obvious discontinuity in areas of unsuitable habitat. Each polygon is coded according to species’ presence (extant, possibly extant or extinct) and origin (native, introduced or reintroduced)41. For some regions covered in workshops (Caucasus, Southeast Asia, much of Africa, Australia and western South America), we collaborated with the Global Assessment of Reptile Distributions (GARD) (http://www.gardinitiative.org/) to provide contributing experts with a baseline species distribution map for review. Although refined maps were returned to the GARD team, not all of these maps have been incorporated into the GARD.
Habitat preferences
Where known, species habitats were coded using the IUCN Habitat Classification Scheme (v.3.1) (https://www.iucnredlist.org/resources/habitat-classification-scheme). Species were assigned to all habitat classes in which they are known to occur. Where possible, habitat suitability (suitable, marginal or unknown) and major importance (yes or no) was recorded. Habitat data were available for 9,484 reptile species.
Threats
All known historical, current and projected (within 10 years or 3 generations, whichever is the longest; generation time estimated, when not available, from related species for which it is known; generation time recorded for 76.3% of the 186 species categorized as threatened under Red List criteria A and C1, the only criteria using generation length) threats were coded using the IUCN Threats Classification Scheme v.3.2 (https://www.iucnredlist.org/resources/threat-classification-scheme), which follows a previously published study42. Where possible, the scope (whole (>90%), majority (50–90%), minority (<50%) of the population; unknown) and severity (causing very rapid (>30%), rapid (>20%), slow but notable (<20%) declines over 10 years or 3 generations, whichever is longer; negligible declines; unknown) of the threat was recorded. Threat data were available for 1,756 of the 1,829 threatened reptile species.
Assessment review
Each assessment underwent two reviews. First, a scientist familiar with the species but not involved in the assessment reviewed the account for biological accuracy and accurate application of the Red List criteria. Once the assessors revised the assessment satisfactorily, staff from the IUCN Red List Unit reviewed the assessment primarily for accurate application of the Red List criteria. The assessors revised the assessment again, if necessary, to satisfy any concerns of the IUCN Red List Unit before the assessment was finalized.
Data limitations
Although we made an extensive effort to complete assessments for all reptiles, some data gaps remain.
Missing species
As of December 2020, 1,145 reptile species, primarily snakes and lizards, were omitted from the present study, including the phylogenetic diversity analyses, because they were described recently and they were described after previous comprehensive assessments from the region. Geographically, they are primarily from tropical regions (as are assessed reptiles) with an underrepresentation of African species (distribution of omitted species: Asia, 41%; Africa, 8%; Australia, 7%; Europe, 3%; North/Central America, 20%; South America, 19%; Caribbean, 5%; Oceania, 4%; percentages sum to greater than 100% because some species occur in two regions). Because they are recently described, many are poorly known, may be rare or occur in a very restricted area, or in poorly surveyed areas that are often subject to high levels of human impacts. As such, recent descriptions are more likely to receive a data-deficient or threatened Red List category than be assigned of least concern41. The net effect on our analyses is a slight underestimate of the number of threatened snakes and lizards, and plausibly a slight overestimate of least concern species. With tetrapod species described in the future likely to be small-ranged, threatened lizards and amphibians43,44, surrogacy levels may decline from those reported here.
Geographical coverage
Although we made extensive efforts to map the current known distribution for each species, this information is incomplete for some species. Where appropriate, and following expert guidance, we interpolated between known localities if the ecological conditions appeared appropriate. In addition, species occurrence is unlikely to be spread evenly or entirely throughout the area depicted in range maps, with gaps expected, for example, in patches of unsuitable habitat.
Data-deficient species
For species assessed to be data deficient (1,507 reptiles, 14.8%), there was inadequate information on the distribution, population status or threats (historical, current or projected future) of the species (both from published sources and expert knowledge) to make a direct, or indirect, assessment of the risk of extinction. All species were assessed according to their recognized taxonomic circumscription at the time of assessment. Taxonomic uncertainty therefore did not result in a data-deficient assignment, although some species were listed as data deficient because they are morphologically indistinguishable from another species and therefore estimates of distribution and abundance are not feasible.
Time span of assessments
The assessments were completed between 1996 and 2020, with 1,503 assessments completed before 2011. The IUCN Rules of Procedure (https://www.iucnredlist.org/resources/rules-of-procedure) recommend reassessment every 10 years and thus, as of 2020, 15% of the assessments can be considered outdated. Of the species assessed 1996–2010, slightly more species were threatened (23.0%) than the species assessed more recently (20.7%). This difference is largely explained by the greater percentages of crocodiles and turtles with outdated assessments (29% and 35%, respectively) compared with tuatara, lizards and snakes (0%, 12% and 17%, respectively) and the highly threatened nature of crocodiles and turtles (Supplementary Table 2). The continuing deterioration of biodiversity globally1 suggests that the species with outdated assessments are more likely to be in higher threat categories today than when they were when last assessed, causing an underestimation of current reptile threat status.
Analyses
Percentage of species threatened with extinction
To estimate the percentage of species threatened with extinction (categories critically endangered, endangered and vulnerable), we used the following formula, which assumes that data-deficient species have the same proportion of threatened species as species that were not data deficient.
where Propthreat is the best estimate of the proportion of species that are threatened; CR, EN, VU and DD are the number of species in each corresponding Red List category and N is the number of species assessed (excluding extinct and extinct in the wild species).
Data for amphibians, birds and mammals
For all analyses that included data for amphibians, birds and mammals, we used the 2020-1 version3 of the tabular and spatial data downloaded from the IUCN Red List website in May 2020.
Threats
Threats calculations were restricted to species in threatened Red List categories (critically endangered, endangered and vulnerable). Multiple threats can affect a single species. Summaries of threats are for the first level of the IUCN classification scheme. Threats thought to affect only a minority of the global population (<50% of the population) (coded as ‘minority’) were not included. In addition, we removed threats that were assessed to cause ‘no declines’ and ‘negligible declines’ from the analysis (as indicated by the severity coding). We considered all threats without scope or severity scored to be major threats and retained them in the analysis.
Habitat
Analyses of habitat use were restricted to the first level of the IUCN habitat-classification scheme. We excluded habitats for which the major importance to the species was scored ‘no’ and suitability was scored ‘marginal’ and considered all habitats without major importance or suitability scored to be suitable and of major importance and included them in the analyses. We did not consider artificial habitats in the analyses.
Only a small number of reptile species inhabits ‘caves/subterranean’ and ‘marine coastal’ habitats, so they were not included in Fig. 4 but their threat prevalence is summarized in Supplementary Table 4.
Statistics
Statistical tests were designed to avoid inclusion of multiple observations from the same species (because species can occur in multiple habitats and be threatened by multiple threats). To assess whether arid habitat or forest species were more likely to be threatened, we included only species that were restricted to one of these habitat types. For threats analyses, we compared species that occur in forests (including those that occur in forests and other habitats) to those that do not occur in forests. All tests were two-tailed Fisher’s exact tests.
Geographical patterns
The geographical patterns of threat and phylogenetic diversity shown in Fig. 2 are for only terrestrial species (so, for reptiles, excluding 87 species of marine turtles and sea snakes). Tetrapod classes vary widely in the numbers of pelagic marine species and in the methods used to map distributions. Restricting analyses to terrestrial species ensured more-consistent analyses and avoided wide variation in summary values caused by small numbers of species.
Analyses of the distribution maps used polygons either with the following IUCN map code designations or with no codes indicated:
Presence = extant (code 1) and probably extant (code 2)
Origin = native (code 1), reintroduced (code 2) and introduced (code 6)
Seasonality = resident (code 1), breeding season (code 2), non-breeding season (code 3) and passage (code 4).
Ranges for species categorized as critically endangered (possibly extinct) are coded as possibly extinct (code 4) and excluded from the spatial analyses.
All spatial analyses were conducted on a global 0.5° by 0.5 ° latitude–longitude grid (approximately 50 km at the Equator). To explore the influence of spatial resolution, we repeated the surrogacy and phylogenetic diversity analyses at a 100-km resolution. We converted polygon range maps (tagged with the appropriate codes as described above) to these grids. We used a global equal-area pseudocylindrical projection, Goode homolosine.
We mapped the distribution of threatened species as a count of the number of species with ranges overlapping each grid cell.
Estimating the spatial distribution of disproportionate threat and phylogenetic diversity loss
We identified global areas in which each tetrapod class is disproportionately threatened compared with all other classes by comparing the species-richness-adjusted level of threat among the four tetrapod classes. First, for each grid cell, we identified the proportional threat level of each class by dividing the number of species in threatened Red List categories (vulnerable, endangered and critically endangered) by the total number of species for the class found in that cell. Second, for all grid cells in which at least five tetrapod species are present, we compared proportional threat values across the four classes and identified a grid cell as having a disproportionate threat level for a given class if: (1) the grid cell had a proportional level of threat equal to 10% or higher for the class; and (2) the grid cell had a proportional level of threat for the class at least twice as high as the proportional level of threat for the next class. We assessed the sensitivity of disproportionate threat patterns to our definition of disproportionate threat by varying the degree of difference in proportional threat level between the highest and second highest class. We identified the number of grid cells with disproportionate threat for each class when the class had a proportional threat level: (1) higher than any other class; (2) 25% or more higher than any other class; (3) 50% or more higher than any other class; (4) 100% or more higher than any other class; and (5) 200% or more higher than any other class. In the main text, we report results for the 100% or more threat level. Results for all thresholds are included in Extended Data Tables 1, 2.
Conservation strategies
We identified global conservation priorities for each tetrapod class using two alternative strategies: strategy 1 prioritized areas containing many threatened species with relatively highly restricted ranges, whereas strategy 2 prioritized areas core to the most range-restricted threatened species globally. We implemented both conservation strategies within the spatial conservation-planning software Zonation45 and the R package zonator46, using the additive benefit function and the core-area Zonation algorithms for strategies 1 and 2, respectively, at 50-km and 100-km resolutions for threatened reptiles.
The additive benefit function algorithm prioritizes areas by the sum of the proportion of the global range size of all species included in a given grid cell—a quantity similar to weighted species endemism (as defined previously47) and endemism richness (as defined48). On the basis of this algorithm, cells with many species occurring only in that cell or few other cells receive the highest priority. The core-area Zonation algorithm prioritizes areas by the maximum proportion of the global range size of all species included in a given grid cell: cells including the highest proportions of the ranges of the most range-restricted species are given the highest priority.
Therefore, comparing the two strategies, strategy 1 gives more importance to the number of species within grid cells (that is, more species = a higher summed proportion), potentially at the expense of the single most-range-restricted species globally, which are instead prioritized directly by strategy 2.
Because complementary representation problems such as these spatial prioritizations often have multiple solutions, we ran five iterations of each algorithm used and summarized variation across those.
Estimating surrogacy
To assess the degree to which conserving the diversity of threatened species of birds, mammals and amphibians (individually or combined) serves as a surrogate for conserving threatened reptile diversity, we calculated a species accumulation index (SAI) of surrogate effectiveness. The SAI is derived from the comparison of three curves: (1) the ‘optimal curve’ represents the accumulation of the diversity of threatened reptile species when conservation is planned using data for threatened reptiles directly; (2) the ‘surrogacy curve’ represents the accumulation of the diversity of threatened reptile species when conservation is planned using the diversity of threatened species diversity of a different class as a surrogate; and (3) the ‘random curve’ represents the accumulation of the diversity of threatened reptile species when conservation areas are selected at random. We estimated optimal, surrogate and random curves based on each reptile-surrogate combination (birds, mammals and amphibians individually and combined). Using 100 sets of approximately random terrestrial grid-cell sequences allowed us to generate 95% confidence intervals around a median ‘random curve’. In addition, because we ran five iterations of each spatial prioritization algorithm for each tetrapod class, optimal and surrogate curves were also summarized using the median and 95% confidence intervals across the five iterations.
We then derived the quantitative measure of surrogacy as SAI = (s − r)/(o − r), where s is the area under the surrogate curve, r is the area under the random curve and o is the area under the optimal curve. SAI = 1 when the optimal and surrogate curves are the same (perfect surrogacy). It is between 1 and 0 when the surrogate curve lies above the random curve (positive surrogacy), zero when the surrogate and random curves coincide (no surrogacy) and negative when the surrogate curve lies below the random curve (negative surrogacy). We calculated the SAI using R code modified from a previous study49. For each reptile–surrogate combination, we report median and 95% confidence intervals across all combinations of optimal, surrogate and random curves (5 target and surrogate curve iterations and 100 random curve iterations).
Although not strictly a measure of surrogacy25, we also calculated the spatial congruence (Spearman’s rank correlation, analogous to a previously published approach9) of Zonation priorities for each conservation strategy and spatial resolution.
Coverage by protected areas
We overlayed protected areas (polygons, categorized as IUCN I–VI from the World Database of Protected Areas50) over the ranges of all threatened tetrapods and classified species with ranges completely outside any protected area as unprotected.
Phylogenetic diversity
To calculate phylogenetic diversity15, we used published time trees of mammals51, birds52 and amphibians53. For reptiles, we combined two time trees: a comprehensive squamate time tree containing 9,755 squamate species, including the species Sphenodon punctatus16, and a turtle and crocodilian tree containing 384 species17. The time trees contain some species lacking genetic data, added by taxonomic interpolation54 to maximize taxonomic coverage. In total, we analysed 32,722 tetrapod species including 10,139 reptiles, 5,364 mammals, 9,879 birds and 7,239 amphibians. For squamates, and for turtles and crocodiles, 10,000 fully resolved trees were available. For each group, we randomly sampled 100 trees and combined them to obtain 100 fully resolved reptile time trees, to accommodate for uncertainty. Similarly, we randomly sampled 100 amphibian and 100 mammal time trees over the 10,000 available.
We thoroughly compared the species name mismatches between geographical and phylogenetic data to match synonyms and correct misspelled names. We also imputed species for which the genus (but not the species) was already present in the tree, for example newly described species (262 amphibian, 1,694 bird, 236 mammal and 777 reptile species). Imputed species were randomly attached to a node within the genus subtree. Because polytomies can result in an overestimation of the phylogenetic diversity, we randomly resolved all polytomies using a previously published method54 implemented in R code. This procedure was performed 100 times for birds, and one time for each of the 100 amphibian, 100 mammal and 100 reptile time trees. We included 30,778 tetrapod species, each with geographical and phylogenetic data, in the phylogenetic diversity analyses. This total included 6,641 amphibians, 8,758 birds, 5,550 mammals and 9,829 reptiles. For each class, we estimated phylogenetic diversity14 for all species and after the removal of threatened species, at 50-km and 100-km resolution. To consider phylogenetic uncertainty (that is, the placement of interpolated species) in phylogenetic diversity calculation for each of the 100 fully resolved trees for each class, we conducted a sensitivity analysis using a previously described method55. This method calculates an evolutionary distinctiveness score that (1) increases the total phylogenetic diversity of the clade when including interpolated species and (2) corrects the evolutionary distinctiveness score of species in genera with interpolated species (missing relatives). Following this method, we calculated evolutionary distinctiveness scores56 for each cell from the subtree including all species present in the focal cell with the R package caper57. For genera with interpolated species, the mean evolutionary distinctiveness score of non-interpolated species was assigned to interpolated species of that genus. For those genera, we computed a second evolutionary distinctiveness score corresponding to the mean evolutionary distinctiveness score of the focal genera (including interpolated species). For species belonging to genera with no interpolated species, the first and second evolutionary distinctiveness scores were identical. Next, we calculated the mean of the two evolutionary distinctiveness scores and reported this value as the evolutionary distinctiveness score of each species. Finally, we computed phylogenetic diversity as the sum of evolutionary distinctiveness scores. Therefore, phylogenetic diversity corresponds to Crozier’s version of phylogenetic diversity58, that is, the sum of the branch lengths connecting all members of a species assemblage without the root. Next, we reported median phylogenetic diversity, computed over 100 fully resolved trees for each class. In the figures, cells with fewer than five species were excluded to avoid outliers.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this paper.
Data availability
Taxonomic data for reptiles were from the Reptile Database (http://www.reptile-database.org/index.html). All spatial and tabular data for the tetrapod analyses are permanently available at https://transfer.natureserve.org/download/Publications/Global_Reptiles/. Trees used for the phylogenetic diversity analyses are available at Zenodo (https://zenodo.org/record/5974891). In addition, assessment data, including range maps, for all tetrapods are publicly available on the IUCN Red List of Threatened Species website (https://iucnredlist.org). Occasionally, where a species may be threatened because of over-collection, sensitive distribution information is not publicly available. Protected areas boundaries were from the World Database of Protected Areas (https://www.protectedplanet.net). Source data are provided with this paper.
Code availability
Python scripts used for the spatial analyses are permanently available at https://transfer.natureserve.org/download/Publications/Global_Reptiles/. No code was used for the Fisher’s exact tests, which were performed in Excel and available with the tabular data at https://transfer.natureserve.org/download/Publications/Global_Reptiles/. Code used for the phylogenetic diversity, areas of disproportionate threat and surrogacy analyses are available at Zenodo (https://zenodo.org/record/5974891).
References
IPBES. Global Assessment Report on Biodiversity and Ecosystem Services (eds Brondizio, E. S. et al.) (2019).
Mair, L. et al. A metric for spatially explicit contributions to science-based species targets. Nat. Ecol. Evol. 5, 836–844 (2021).
IUCN. The IUCN Red List of Threatened Species v.2020-1 https://www.iucnredlist.org/ (2020).
Tilman, D. et al. Future threats to biodiversity and pathways to their prevention. Nature 546, 73–81 (2017).
Mason, N., Ward, M., Watson, J. E. M., Venter, O. & Runting, R. K. Global opportunities and challenges for transboundary conservation. Nat. Ecol. Evol. 4, 694–701 (2020).
Roll, U. et al. The global distribution of tetrapods reveals a need for targeted reptile conservation. Nat. Ecol. Evol. 1, 1677–1682 (2017).
Harfoot, M. B. J. et al. Using the IUCN Red List to map threats to terrestrial vertebrates at global scale. Nat. Ecol. Evol. 5, 1510–1519 (2021).
Böhm, M. et al. The status of the world’s reptiles. Biol. Conserv. 157, 372–385 (2013).
Grenyer, R. Global distribution and conservation of rare and threatened vertebrates. Nature 444, 93–96 (2006).
Rodrigues, A. S. L. Effective global conservation strategies. Nature 450, E19 (2007).
Gumbs, R. et al. Global priorities for conservation of reptilian phylogenetic diversity in the face of human impacts. Nat. Commun. 11, 2616 (2020).
Pincheira-Donoso, D., Bauer, A. M., Meiri, S. & Uetz, P. Global taxonomic diversity of living reptiles. PLoS ONE 8, e59741 (2013).
Hay, J. M., Sarre, S. D., Lambert, D. M., Allendorf, F. W. & Daugherty, C. H. Genetic diversity and taxonomy: a reassessment of species designation in tuatara (Sphenodon: Reptilia). Conserv. Genet. 11, 1063–1081 (2010).
Hedges, S. B. & Conn, C. E. A new skink fauna from Caribbean islands (Squamata, Mabuyidae, Mabuyinae). Zootaxa 3288, 1–244 (2012).
Faith, D. P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 61, 1–10 (1992).
Tonini, J. F. R., Beard, K. H., Ferreira, R. B., Jetz, W. & Pyron, R. A. Fully-sampled phylogenies of squamates reveal evolutionary patterns in threat status. Biol. Conserv. 204, 23–31 (2016).
Colston, T. J., Kulkarni, P., Jetz, W. & Pyron, R. A. Phylogenetic and spatial distribution of evolutionary diversification, isolation, and threat in turtles and crocodilians (non-avian archosauromorphs). BMC Evol. Biol. 20, 81 (2020).
Sinervo, B. et al. Erosion of lizard diversity by climate change and altered thermal niches. Science 328, 894–899 (2010).
Valenzuela, N. et al. Extreme thermal fluctuations from climate change unexpectedly accelerate demographic collapse of vertebrates with temperature-dependent sex determination. Sci. Rep. 9, 4254 (2019).
Diele-Viegas, L. M. & Rocha, C. F. D. Unraveling the influences of climate change in Lepidosauria (Reptilia). J. Therm. Biol. 78, 401–414 (2018).
Trull, N., Böhm, M. & Carr, J. Patterns and biases of climate change threats in the IUCN Red List. Conserv. Biol. 32, 135–147 (2018).
Lorch, J. M. et al. Snake fungal disease: an emerging threat to wild snakes. Phil. Trans. R. Soc. B 371, 20150457 (2016).
Marshall, B. M., Strine, C. & Hughes, A. C. Thousands of reptile species threatened by under-regulated global trade. Nat. Commun. 11, 4738 (2020).
Powney, G. D., Grenyer, R., Orme, C. D. L., Owens, I. P. F. & Meiri, S. Hot, dry and different: Australian lizard richness is unlike that of mammals, amphibians and birds. Glob. Ecol. Biogeogr. 19, 386–396 (2010).
Rapacciuolo, G. et al. Species diversity as a surrogate for conservation of phylogenetic and functional diversity in terrestrial vertebrates across the Americas. Nat. Ecol. Evol. 3, 53–61 (2019).
Huey, R. B. et al. Predicting organismal vulnerability to climate warming: roles of behaviour, physiology and adaptation. Phil. Trans. R. Soc. B 36, 1665–1679 (2012).
Stanford, C. B. et al. Turtles and tortoises are in trouble. Curr. Biol. 30, R721–R735 (2020).
Diele-Viegas, L. M., Figueroa, R. T., Vilela, B. & Rocha, C. F. D. Are reptiles toast? A worldwide evaluation of Lepidosauria vulnerability to climate change. Clim. Change 159, 581–599 (2020).
Grace, M. K. et al. Testing a global standard for quantifying species recovery and assessing conservation impact. Conserv. Biol. 35, 1833–1849 (2021).
IUCN. A Global Standard for the Identification of Key Biodiversity Areas v.1.0 (2016).
Montesino Pouzols, F. et al. Global protected area expansion is compromised by projected land-use and parochialism. Nature 516, 383–386 (2014).
Butchart, S. H. M. et al. Improvements to the Red List Index. PLoS ONE 2, e140 (2007).
Convention on Biological Diversity. First Draft of the Post-2020 Global Biodiversity Framework, CBD/WG2020/3/3 https://www.cbd.int/conferences/post2020/wg2020-03/documents (2021).
IUCN. IUCN Red List Categories and Criteria v.3.1 https://www.iucnredlist.org/resources/categories-and-criteria (2012).
IUCN Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria v.14 https://www.iucnredlist.org/resources/redlistguidelines (IUCN, 2019).
Stuart, S. N. et al. Status and trends of amphibian declines and extinctions worldwide. Science 306, 1783–1786 (2004).
Schipper, J. et al. The status of the world’s land and marine mammals: diversity, threat, and knowledge. Science 322, 225–230 (2008).
Rhodin, A. G. J. et al. Global conservation status of turtles and tortoises (order Testudines). Chelonian Conserv. Biol. 17, 135–161 (2018).
Uetz, P., Freed, P., Aguilar, R. & Hošek, J. The Reptile Database http://reptile-database.org/ (2020).
Turtle Taxonomy Working Group. Turtles of the World: Annotated Checklist and Atlas of Taxonomy, Synonymy, Distribution, and Conservation Status. 8th edn Chelonian Research Monograph No.7 (Chelonian Research Foundation and Turtle Conservancy, 2017).
IUCN. Documentation Standards and Consistency Checks for IUCN Red List Assessments and Species Accounts. v.2 https://www.iucnredlist.org/resources/supporting-information-guidelines (2019).
Salafsky, N. et al. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conserv. Biol. 22, 897–911 (2008).
Meiri, S. Small, rare and trendy: traits and biogeography of lizards described in the 21st century. J. Zool. 299, 251–261 (2016).
Moura, M. R. & Jetz, W. Shortfalls and opportunities in terrestrial vertebrate species discovery. Nat. Ecol. Evol. 5, 631–639 (2021).
Moilanen, A., Montesino Pouzols, F., Meller, L. & Veach, V. Spatial Conservation Planning Methods and Software Zonation. User manual v.4 https://github.com/cbig/zonation-core/releases (2014).
Lehtomaki, J. zonator. R package v.0.6.0 GitHub https://github.com/cbig/zonator (2020).
Crisp, M. D., Laffan, S., Linder, H. P. & Monro, A. Endemism in the Australian flora. J. Biogeogr. 28, 183–198 (2001).
Kier, G. & Barthlott, W. Measuring and mapping endemism and species richness: a new methodological approach and its application on the flora of Africa. Biodivers. Conserv. 10, 1513–1529 (2001).
Sabatini, F. M. et al. One taxon does not fit all: herb-layer diversity and stand structural complexity are weak predictors of biodiversity in Fagus sylvatica forests. Ecol. Indic. 69, 126–137 (2016).
UNEP-WCMC & IUCN. Protected Planet: the World Database on Protected Areas (WDPA) https://www.protectedplanet.net (2021).
Upham, N. S., Esselstyn, J. A. & Jetz, W. Inferring the mammal tree: species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS Biol. 17, e3000494 (2019).
Hedges, S. B., Marin, J., Suleski, M., Paymer, M. & Kumar, S. Tree of life reveals clock-like speciation and diversification. Mol. Biol. Evol. 32, 835–845 (2015).
Jetz, W. & Pyron, R. A. The interplay of past diversification and evolutionary isolation with present imperilment across the amphibian tree of life. Nat. Ecol. Evol. 2, 850–858 (2018).
Rangel, T. F. et al. Phylogenetic uncertainty revisited: implications for ecological analyses. Evolution 69, 1301–1312 (2015).
Gumbs, R., Gray, C. L., Wearn, O. R. & Owen, N. R. Tetrapods on the EDGE: overcoming data limitations to identify phylogenetic conservation priorities. PLoS ONE 13, e0194680 (2018).
Redding, D. W. & Mooers, A. Ø. Incorporating evolutionary measures into conservation prioritization. Conserv. Biol. 20, 1670–1678 (2006).
Orme, D. et al. caper: comparative analyses of phylogenetics and evolution in R. R package v.0.5.2 https://rdrr.io/cran/caper/man/caper-package.html (2013).
Tucker, C. M. et al. Assessing the utility of conserving evolutionary history. Biol. Rev. Camb. Philos. Soc. 94, 1740–1760 (2019).
Tolley, K. A. et al. Clinging to survival: critically endangered Chapman’s pygmy chameleon Rhampholeon chapmanorum persists in shrinking forest patches. Oryx https://doi.org/10.1017/S0030605320000952 (2021).
Acknowledgements
We thank the 961 Red List assessors (listed in Supplementary Note 1); the organizers of workshops in United Arab Emirates (D. Mallon), Fiji (H. Pippard) and Colombia (B. Bock and V. Páez); the co-principal investigators of NSF award 1136586 (G. Costa, C. Graham and V. Radeloff); and J. Chanson, D. Bennett, N. Garcia, H. Hamilton, D. Hole, D. Juhn, A. Kenlan, M. Mascia, R. Mittermeier, M. Ormes, J. P. Ross, J. P. Rodriguez, J. Rodriguez, J. Smart, S. Stuart, P. Uetz and J.-C. Vié. Funding was provided by Agencia Nacional de Promoción Científica y Tecnológica-Fondo para la Investigación Científica y Tecnológica RC 2014-0116 (L.J.A.); the Australian Department of Agriculture, Water and the Environment; the Australian Research Council; B. and G. Moore; Charles Darwin University; Conselho Nacional de Desenvolvimento Científico e Tecnológico; Coordenação de Aperfeiçoamento de Pessoal de Nível Superior; the Environment Agency Abu Dhabi; Environment and Protected Areas Authority of Sharjah; Fundação de Amparo à Pesquisa do Estado de São Paulo grants 2015/20215-7 (C.d.C.N.) and 2020/12658-4 (M.M.); Fundação de Apoio à Pesquisa do Distrito Federal; Global Protected Area Friendly System; IBAT—The Integrated Biodiversity Assessment Tool; Instituto Chico Mendes de Conservação da Biodiversidade; IUCN; Ministério do Meio Ambiente; the Mohamed bin Zayed Species Conservation Fund; Monash University; Museum für Naturkunde; National Science Foundation grants 1136586 (B.E.Y. and T.M.B.), 1455761 (S.B.H.) and 1932765 (S.B.H.); the Rainforest Trust; the Regina Bauer Frankenberg Foundation; the Rufford Foundation; San Diego Zoo Wildlife Alliance; Secretaría de Educación Superior, Ciencia, Tecnología e Innovación; the Society of Entrepreneurs and Ecology Foundation; the South African National Biodiversity Institute; Species Survival Commission; Toyota Motor Corporation through the IUCN-Toyota Red List Partnership; US Fish and Wildlife Service; Universidad del Bío-Bío; Universidad San Francisco de Quito; University of Western Australia; Wildlife Reserves Singapore; World Wildlife Fund; Zoological Institute, St Petersburg theme 122031100282-2; and Zoological Society of London.
Author information
Authors and Affiliations
Contributions
N.C. and B.E.Y. conceived the original idea. C.H.-T. curated the data. M.F., J.M., G.R. and B.E.Y. ran the analyses. T.M.B., N.C., S.B.H., R.K.B.J. and B.E.Y. secured the principal funding. N.C. and B.E.Y. wrote the first draft and all authors reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Céline Bellard, Arne Mooers, Mario Moura and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Locations of critically endangered (possibly extinct) reptiles.
Colours depict individual species’ ranges. These species were classified as critically endangered (possibly extinct) (CR(PE)) at the time of their assessments. Fortunately, a few species such as Rhampholeon chapmanorum59 have been recorded subsequent to their assessment publication dates and will no longer be classified as CR(PE) once they are reassessed in the future.
Extended Data Fig. 2 Richness of threatened (CR, EN, VU) tetrapods (50-km resolution).
As in Fig. 2a, only threatened species occurring in terrestrial areas are shown.
Extended Data Fig. 3 Areas with the top 15% of PD loss if all threatened reptiles become extinct.
a, 50-km resolution, b, 100-km resolution.
Extended Data Fig. 4 Species Accumulation Index (SAI) values and curves for surrogacy. analysis.
Effectiveness of threatened birds, mammals, and amphibians – individually or combined – as a surrogate for the conservation of threatened reptile species diversity for two alternative strategies – a, Conservation Strategy 1: Maximize rarity-weighted richness, b, Conservation Strategy 2: Maximize inclusion of most range-restricted species. See Methods for full explanation of each strategy. Surrogate effectiveness is measured using the Species Accumulation Index (SAI): values range from −∞ to 1, with 1 indicating perfect surrogacy, values between 1 and 0 indicate positive surrogacy, 0 indicating no surrogacy, and values less than 0 indicating negative surrogacy. In each panel, median SAI values are in bold, with lower and upper confidence intervals in brackets. Blue lines are the optimal curves (accumulation of target diversity based on target priority areas); red lines are the surrogate curves (accumulation of target diversity based on surrogate priority areas); and grey lines are the random curves (accumulation of target diversity based on random selection of areas). Confidence intervals (95%, based on 100 randomizations) shown in lighter shading around curves; most are too small to be visible.
Supplementary information
Supplementary Information
This file contains Supplementary Note 1 and Supplementary Tables 1–3.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cox, N., Young, B.E., Bowles, P. et al. A global reptile assessment highlights shared conservation needs of tetrapods. Nature 605, 285–290 (2022). https://doi.org/10.1038/s41586-022-04664-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-04664-7
This article is cited by
-
Global conservation status of the jawed vertebrate Tree of Life
Nature Communications (2024)
-
Modelling reveals the effect of climate and land use change on Madagascar’s chameleons fauna
Communications Biology (2024)
-
Global evaluation of current and future threats to drylands and their vertebrate biodiversity
Nature Ecology & Evolution (2024)
-
Wildlife ecotoxicology of plant protection products: knowns and unknowns about the impacts of currently used pesticides on terrestrial vertebrate biodiversity
Environmental Science and Pollution Research (2024)
-
Invasive black rats menacing endangered lizards
Biodiversity and Conservation (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
