
Abstract
The liberation of energy stores from adipocytes is critical to support survival in times of energy deficit; however, uncontrolled or chronic lipolysis associated with insulin resistance and/or insulin insufficiency disrupts metabolic homeostasis1,2. Coupled to lipolysis is the release of a recently identified hormone, fatty-acid-binding protein 4 (FABP4)3. Although circulating FABP4 levels have been strongly associated with cardiometabolic diseases in both preclinical models and humans4,5,6,7, no mechanism of action has yet been described8,9,10. Here we show that hormonal FABP4 forms a functional hormone complex with adenosine kinase (ADK) and nucleoside diphosphate kinase (NDPK) to regulate extracellular ATP and ADP levels. We identify a substantial effect of this hormone on beta cells and given the central role of beta-cell function in both the control of lipolysis and development of diabetes, postulate that hormonal FABP4 is a key regulator of an adipose–beta-cell endocrine axis. Antibody-mediated targeting of this hormone complex improves metabolic outcomes, enhances beta-cell function and preserves beta-cell integrity to prevent both type 1 and type 2 diabetes. Thus, the FABP4–ADK–NDPK complex, Fabkin, represents a previously unknown hormone and mechanism of action that integrates energy status with the function of metabolic organs, and represents a promising target against metabolic disease.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
Data availability
The datasets generated during the current study are available in the FigShare repository at https://doi.org/10.6084/m9.figshare.16547853. Source data are provided with this paper.
References
Jensen, M. D., Caruso, M., Heiling, V. & Miles, J. M. Insulin regulation of lipolysis in nondiabetic and IDDM subjects. Diabetes 38, 1595–1601 (1989).
Kim, J. Y. et al. Increased lipolysis, diminished adipose tissue insulin sensitivity, and impaired beta-cell function relative to adipose tissue insulin sensitivity in obese youth with impaired glucose tolerance. Diabetes 66, 3085–3090 (2017).
Cao, H. et al. Adipocyte lipid chaperone AP2 is a secreted adipokine regulating hepatic glucose production. Cell Metab. 17, 768–778 (2013).
Prentice, K. J., Saksi, J. & Hotamisligil, G. S. Adipokine FABP4 integrates energy stores and counterregulatory metabolic responses. J. Lipid Res. 60, 734–740 (2019).
Saksi, J. et al. Low-expression variant of fatty acid-binding protein 4 favors reduced manifestations of atherosclerotic disease and increased plaque stability. Circ. Cardiovasc. Genet 7, 588–598 (2014).
Tuncman, G. et al. A genetic variant at the fatty acid-binding protein aP2 locus reduces the risk for hypertriglyceridemia, type 2 diabetes, and cardiovascular disease. Proc. Natl Acad. Sci. USA 103, 6970–6975 (2006).
Zhao, W. et al. Identification of new susceptibility loci for type 2 diabetes and shared etiological pathways with coronary heart disease. Nat. Genet. 49, 1450–1457 (2017).
Ertunc, M. E. et al. Secretion of fatty acid binding protein aP2 from adipocytes through a nonclassical pathway in response to adipocyte lipase activity. J. Lipid Res. 56, 423–434 (2015).
Schlottmann, I., Ehrhart-Bornstein, M., Wabitsch, M., Bornstein, S. R. & Lamounier-Zepter, V. Calcium-dependent release of adipocyte fatty acid binding protein from human adipocytes. Int. J. Obes. 38, 1221–1227 (2014).
Villeneuve, J. et al. Unconventional secretion of FABP4 by endosomes and secretory lysosomes. J. Cell Biol. 217, 649–665 (2018).
Burak, M. F. et al. Development of a therapeutic monoclonal antibody that targets secreted fatty acid-binding protein aP2 to treat type 2 diabetes. Sci. Transl. Med. 7, 319ra205 (2015).
Thumer, L. et al. German new onset diabetes in the young incident cohort study: DiMelli study design and first-year results. Rev. Diabet. Stud. 7, 202–208 (2010).
Warncke, K. et al. Does diabetes appear in distinct phenotypes in young people? Results of the diabetes mellitus incidence Cohort Registry (DiMelli). PLoS ONE 8, e74339 (2013).
Ziegler, A. G., Hummel, M., Schenker, M. & Bonifacio, E. Autoantibody appearance and risk for development of childhood diabetes in offspring of parents with type 1 diabetes: the 2-year analysis of the German BABYDIAB Study. Diabetes 48, 460–468 (1999).
Xiao, Y. et al. Fatty acid binding protein 4 promotes autoimmune diabetes by recruitment and activation of pancreatic islet macrophages. JCI Insight 6, e141814 (2021).
Kralisch, S. et al. Circulating adipocyte fatty acid-binding protein induces insulin resistance in mice in vivo. Obesity 23, 1007–1013 (2015).
Wu, L. E. et al. Identification of fatty acid binding protein 4 as an adipokine that regulates insulin secretion during obesity. Mol. Metab. 3, 465–473 (2014).
Annes, J. P. et al. Adenosine kinase inhibition selectively promotes rodent and porcine islet beta-cell replication. Proc. Natl Acad. Sci. USA 109, 3915–3920 (2012).
Navarro, G. et al. Genetic disruption of adenosine kinase in mouse pancreatic beta-cells protects against high-fat diet-induced glucose intolerance. Diabetes 66, 1928–1938 (2017).
Furuhashi, M. et al. Treatment of diabetes and atherosclerosis by inhibiting fatty-acid-binding protein aP2. Nature 447, 959–965 (2007).
Khan, S. et al. Autocrine activation of P2Y1 receptors couples Ca2+ influx to Ca2+ release in human pancreatic beta-cells. Diabetologia 57, 2535–2545 (2014).
Leon, C. et al. The P2Y1 receptor is an ADP receptor antagonized by ATP and expressed in platelets and megakaryoblastic cells. FEBS Lett. 403, 26–30 (1997).
Detimary, P., Jonas, J. C., & Henquin, J. C. Stable and diffusible pools of nucleotides in pancreatic islet cells. Endocrinology 137, 4671–4676 (1996).
Bauer, C. et al. ATP mediates a negative autocrine signal on stimulus-secretion coupling in mouse pancreatic beta-cells. Endocrine 63, 270–283 (2019).
Baurand, A. et al. Inhibition of platelet function by administration of MRS2179, a P2Y1 receptor antagonist. Eur. J. Pharmacol. 412, 213–221 (2001).
Seo, J. B., Jung, S. R., Hille, B. & Koh, D. S. Extracellular ATP protects pancreatic duct epithelial cells from alcohol-induced damage through P2Y1 receptor-cAMP signal pathway. Cell Biol. Toxicol. 32, 229–247 (2016).
Engin, F. et al. Restoration of the unfolded protein response in pancreatic beta-cells protects mice against type 1 diabetes. Sci. Transl. Med. 5, 211ra156 (2013).
Engin, F., Nguyen, T., Yermalovich, A. & Hotamisligil, G. S. Aberrant islet unfolded protein response in type 2 diabetes. Sci. Rep. 4, 4054 (2014).
Burnstock, G., Vaughn, B. & Robson, S. C. Purinergic signalling in the liver in health and disease. Purinergic Signal. 10, 51–70 (2014).
Burnstock, G. Purinergic signaling in the cardiovascular system. Circ. Res. 120, 207–228 (2017).
Liu, Y. et al. Rapid Elevation in CMPF may act as a tipping point in diabetes development. Cell Rep. 14, 2889–2900 (2016).
Batchuluun, B. et al. Elevated medium-chain acylcarnitines are associated with gestational diabetes mellitus and early progression to type 2 diabetes and induce pancreatic beta-cell dysfunction. Diabetes 67, 885–897 (2018).
Prentice, K. J. et al. The furan fatty acid metabolite CMPF is elevated in diabetes and induces beta-cell dysfunction. Cell Metab. 19, 653–666 (2014).
Rezania, A. et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 32, 1121–1133 (2014).
Potel, C. M., Fasci, D. & Heck, A. J. R. Mix and match of the tumor metastasis suppressor Nm23 protein isoforms in vitro and in vivo. FEBS J. 285, 2856–2868 (2018).
Han, B. G., Min, K., Lee, B. I. & Lee, S. Refined structure of human NM23-H1 from a hexagonal crystal. Bull. Korean Chem. Soc. 31, 1397–1399 (2010).
Prentice, K. J. et al. CMPF, a metabolite formed upon prescription omega-3-acid ethyl ester supplementation, prevents and reverses steatosis. EBioMedicine 27, 200–213 (2018).
Basford, C. L. et al. The functional and molecular characterisation of human embryonic stem cell-derived insulin-positive cells compared with adult pancreatic beta-cells. Diabetologia 55, 358–371 (2012).
Acknowledgements
We thank members of the Hotamisligil Laboratory and the Sabri Ülker Center, past and present, for their contributions to our understanding of FABP4 and their helpful discussions. We thank staff at the Islet Core and Clinical Islet Laboratory (Alberta Islet Distribution Program, University of Alberta) for providing us with human islets from review-board-approved donors. UCB generated and purified the antibodies used in this study and independently reproduced the interaction data between NDPK, FABP4 and a-Ab. The Hotamisligil laboratory is supported by grants from the National Institutes of Health (NIH DK123458) and the Juvenile Diabetes Research Foundation (JDRF; 2-SRA-2019-660-S-B). A JDRF Postdoctoral Fellowship (3-PDF-2017-400-A-N) funds K.J.P. The Sigrid Juselius Foundation, the Finnish Foundation for Cardiovascular Research and the Otto A. Malm Foundation support J.S. F.E. was supported by grants from the JDRF (JDRF-5-CDA-2014-184-A-N) and the NIH (NIH 5K01DK102488-03).
Author information
Authors and Affiliations
Contributions
K.J.P. designed and performed the in vitro and in vivo experiments, analysed the data, prepared the figures and wrote and revised the manuscript. F.E., K.E.I., A.L., P.C. and L.T.R. designed and performed the in vivo experiments, analysed data and revised the manuscript. J.S., G.Y.L., K.E., E.O. and E.S.C. designed and performed the in vitro experiments, analysed data and revised the manuscript. O.C. analysed the crystal structure and generated recombinant proteins used in this study, analysed the data and revised the manuscript. P.A. and A.-G.Z. collected and analysed human serum samples and revised the manuscript. G.S.H. conceived, supervised and supported the project, designed experiments, interpreted results and revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The Hotamisligil Lab has generated intellectual property (assigned to Harvard University) related to hormonal FABP4 and its therapeutic targeting and receives funding for this project from Lab1636, LLC, an affiliate of Deerfield Management. G.S.H. is on the Scientific Advisory Board of Crescenta Pharmaceuticals and holds equity. Other authors have no conflicts of interest to declare.
Additional information
Peer review information Nature thanks Matthias Hebrok, Andrew Stewart, Thomas Wieland and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
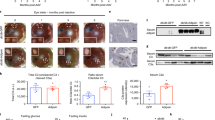
Extended Data Fig. 1 FABP4-/- mice have no difference in alpha cell mass, and FABP4 is not expressed in beta cells.
(a) Dithizone-stained islets in vivo from FABP4−/− mice (Representative image from N = 3). (b) Immunohistochemical staining for glucagon of pancreatic sections from 7-week-old WT or FABP4−/− mice, and (c) quantification of percentage of glucagon positive area per total pancreatic area based on IHC (N = 5/group). (d) Immunofluorescent staining for insulin (green), FABP4 (red) and nuclei (DAPI, blue) in primary isolated mouse islets (N = 15 islets). (e) Western blot for FABP4 and B-Tubulin (B-Tubb) from INS1 cells with and without treatment with FABP4 (N = 3). Baseline matching parameters for 10 week old WT female NOD mice prior to the initiation of dosing showing no difference in (f) body weight, (g) blood glucose, (h) plasma insulin, or (i) plasma FABP4 (N = 46/group). (j) 6 h fasting blood glucose, (k) body weight, and (l) plasma insulin levels among NOD mice that remained non-diabetic for the duration of the treatment period (N = 11 (PBS), 23 (a-Ab), 13 (c-Ab)). (m) Immunofluorescent staining for insulin (green) and glucagon (red) in pancreatic sections from NOD mice treated with a-Ab or c-Ab for 5 wks (N = 4 mice/group). Scale bars are 100um. Data are mean +/− SEM. Two-tailed unpaired t-test (c); One-way ANOVA (f-i); Two-way ANOVA (j-l).
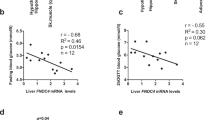
Extended Data Fig. 2 FABP4 exerts differential activity in vivo and in vitro, and only interacts with a-Ab through the light chain.
(a) GSIS from islets from FABP4−/− mice treated with increasing doses of FABP4 (N = 3). (b) Plasma FABP4 levels following acute injection of 10 µg FABP4 (N = 3 (PBS), 6 (FABP4)). (c) Blood glucose (P = 0.0300) and (d) plasma insulin levels 20 min post-injection (P = 0.0160) with FABP4 or PBS (N = 3 (PBS), 6 (FABP4)).(e) Crystal structure showing FABP4 binding through the light chain of a-Ab. *P < 0.05, **P < 0.01, ***P < 0.001. Data are presented as mean +/− SEM. Two-way ANOVA (a,b); Welch’s t-test (c,d).
Extended Data Fig. 3 Determination of the Binding Epitope for a-Ab on NDPK-A.
(a) Amino acid sequence for human NDPK-A, and indication of the location of each peptide generated for epitope mapping (red). Each peptide is 15 amino acids long with an N-terminal 6xHis Tag for peptide labeling. Each peptide has a 5 amino acid overlap with the preceeding peptide sequentially from the N-terminus of the protein.(b) Table summarizing binding affinity as determined by MicroScale Thermophoresis (MST). Low affinity binding was observed between peptide 2 and peptide 3 with a-Ab. High affinity binding was observed between peptide 8 and a-Ab, comparable to full-length NDPK-A protein. No binding was detected (NBD) between a-Ab and any other peptide examined. The lack of binding between peptide 7 or peptide 9 and a-Ab indicated that the central 5 amino acids of peptide 8, non-overlapping in sequence with either peptide 7 or 9, is likely the primary epitope for a-A binding. (c) Example MST binding curves for full-length NDPK-A, NDPK-A peptide 9 and NDPK-A peptide 8 with a-Ab. No binding is observed between NDPK-A peptide 9 and a-Ab. (d) Crystal structure of human NDPK-A in hexamer conformation (source PDB: 3L7U) with the potential binding region of peptide 8 highlighted in pink and peptide 3 highlighted in blue. The protein structure indicates the potential binding region of peptide 8 is on the surface of the protein, amenable for a-Ab interaction. The tertiary folding of NDPK-A places peptide 3 and peptide 8 in close proximity, suggesting binding of a-Ab may be primarily occurring through residues in peptide 8, and partially facilitated through interaction with residues in peptides 2 and 3. EC50 calculated by Hill Slope.
Extended Data Fig. 4 a-Ab interacts with Fabkin.
(a–e) Representative mesoscale thermophoresis experiments showing 1:1 protein interactions between complex components and a-Ab (N = 6/interaction). EC50 was calculated using Hill Slope. (f) Representtive Western blot and (g) quantification showing relative abundance of complex components interacting with GST-NDPK-A (N = 4 replicates; P < 0.0001). Kinase activity of recombinant ADK to generate (h) ATP and (i) ADP in the presence of complex components (N = 3/group). Kinase activity of recombinant NDPK to generate (j) ADP and (k) ADP in the presence of complex components (N = 3/group). Activity of recombinant (l) ADK and (m) NDPK in the presence of a-Ab or c-Ab alone (N = 3/group). *P < 0.05, **P < 0.01, ***P < 0.001. Data are presented as mean +/− SEM. Two-way ANOVA (g-m).
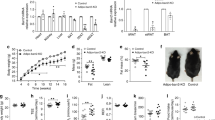
Extended Data Fig. 5 Complex components alone do not alter GSIS.
(a) GSIS from WT mouse islets treated with each of the proposed complex components alone (N = 4/condition). (b) GSIS from INS1 cells treated with NDPK-ADK with FABP4, lipid binding mutant (LBM) FABP4, or FABP4 pre-treated with inhibitor BMS-309403 (N = 4/group). *P < 0.05, ***P < 0.001. Data are mean +/− SEM. One-way ANOVA (a,b).
Extended Data Fig. 6 Treatment with a-Ab preserves beta cell mass and islet number with minimal alterations to the pancreas immune profile.
Cytosolic calcium flux in INS1 cells from (a,b) the extracellular space (N = 2-3 coverslips/treatment; 88 (Control), 100 (NDPK-ADK-FABP4), 47 (MRS2365) cells) and (c,d) the ER in response to thapsigargin as determined by Fura-2 AM staining in control conditions or pretreatment with FABP4-ADK-NDPK with or without MRS2365 (N = 2-3 coverslips/treatment; 50 cells).(e) Cytosolic calcium flux from the ER in INS1 cells under control conditions, or pretreatment with FABP4-ADK-NDPK with or without the adenylyl cyclase inhibitor NKY80 (N = 2-3 coverslips/treatment; 100 (Control, NDPK-ADK-FABP4), 150 (NKY80) cells). (f) Quantification of Western blots from Fig. 4g (N = 4/condition). (g) Gene expression of ER stress markers BIP following 2hr treatment with NDPK-ADK, FABP4-ADK-NDPK, or FABP4-ADK-NDPK with a-Ab in the presence or absence of Tg (N = 3). (h) Cleaved caspase 3/7 activity in INS1 cells treated with increasing concentrations of Tg with or without FABP4-ADK-NDPK (N = 4). (i) Percentage of CD45+ cells (N = 12), (j) regulatory T-cells (N = 6), (k) cytotoxic T-cells (N = 6), (l) T-Helper cells (N = 5), (m) B-cells (N = 6), (n) dendritic cells (N = 6), and (o) granulocytes (N = 6) in whole pancreas from mice treated with PBS, a-Ab or c-Ab for 14 weeks by one-way ANOVA. *P < 0.05, **P < 0.01, ***P < 0.001. Data are presented as mean +/− SEM. One-way ANOVA (b,d,i–o); Two-tailed unpaired t-test (f); Two-way ANOVA (g,h).
Supplementary information
Supplementary Figure 1
This file contains the uncropped blots.
Rights and permissions
About this article

Cite this article
Prentice, K.J., Saksi, J., Robertson, L.T. et al. A hormone complex of FABP4 and nucleoside kinases regulates islet function. Nature 600, 720–726 (2021). https://doi.org/10.1038/s41586-021-04137-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-021-04137-3
This article is cited by
-
PRUNE1 and NME/NDPK family proteins influence energy metabolism and signaling in cancer metastases
Cancer and Metastasis Reviews (2024)
-
Glomerular expression and urinary excretion of fatty acid-binding protein 4 in IgA nephropathy
Journal of Nephrology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
