
Abstract
The ordered assembly of tau protein into filaments characterizes several neurodegenerative diseases, which are called tauopathies. It was previously reported that, by cryo-electron microscopy, the structures of tau filaments from Alzheimer’s disease1,2, Pick’s disease3, chronic traumatic encephalopathy4 and corticobasal degeneration5 are distinct. Here we show that the structures of tau filaments from progressive supranuclear palsy (PSP) define a new three-layered fold. Moreover, the structures of tau filaments from globular glial tauopathy are similar to those from PSP. The tau filament fold of argyrophilic grain disease (AGD) differs, instead resembling the four-layered fold of corticobasal degeneration. The AGD fold is also observed in ageing-related tau astrogliopathy. Tau protofilament structures from inherited cases of mutations at positions +3 or +16 in intron 10 of MAPT (the microtubule-associated protein tau gene) are also identical to those from AGD, suggesting that relative overproduction of four-repeat tau can give rise to the AGD fold. Finally, the structures of tau filaments from cases of familial British dementia and familial Danish dementia are the same as those from cases of Alzheimer’s disease and primary age-related tauopathy. These findings suggest a hierarchical classification of tauopathies on the basis of their filament folds, which complements clinical diagnosis and neuropathology and also allows the identification of new entities—as we show for a case diagnosed as PSP, but with filament structures that are intermediate between those of globular glial tauopathy and PSP.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others
Data availability
Cryo-EM maps have been deposited in the Electron Microscopy Data Bank (EMDB) under accession numbers EMD-13218 for PSP, EMD-13219 for GGT-I, EMD-13220 for GGT-II, EMD-13221 for GGT-III, EMD-13223 for GPT type 1a, EMD-13224 for GPT type 1b, EMD-13225 for GPT type 2, EMD-13226 for AGD type 1 and EMD-13227 for AGD type 2. Corresponding refined atomic models have been deposited in the Protein Data Bank (PDB) under accession numbers 7P65 for PSP, 7P66 for GGT type 1, 7P67 for GGT type 2, 7P68 for GGT type 3, 7P6A for GPT type 1a, 7P6B for GPT type 1b, 7P6C for GPT type 2, 7P6D for AGD type 1 and 7P6E for AGD type 2. Cryo-EM datasets have been deposited in the Electron Microscopy Public Image Archive (EMPIAR) under accession numbers 10765 for PSP-RS case 1, 10766 for the GGT-I case, 10767 for PSP-F case 2, 10768 for AGD case 1, and 10769 for the +16 case. Any other relevant data are available from the corresponding authors upon request.
References
Fitzpatrick, A. W. P. et al. Cryo-EM structures of tau filaments from Alzheimer’s disease. Nature 547, 185–190 (2017).
Falcon, B. et al. Tau filaments from multiple cases of sporadic and inherited Alzheimer’s disease adopt a common fold. Acta Neuropathol. 126, 699–708
Falcon, B. et al. Structures of filaments from Pick’s disease reveal a novel tau protein fold. Nature 561, 137–140 (2018).
Falcon, B. et al. Novel tau filament fold in chronic traumatic encephalopathy encloses hydrophobic molecules. Nature 568, 420–423 (2019).
Zhang, W. et al. Novel tau filament fold in corticobasal degeneration. Nature 580, 283–287 (2020).
Goedert, M., Spillantini, M. G., Jakes, R., Rutherford, D. & Crowther, R. A. Multiple isoforms of human microtubule-associated protein tau: sequences and localization in neurofibrillary tangles of Alzheimer’s disease. Neuron 3, 519–526 (1989).
Ghetti, B. et al. Frontotemporal dementia caused by microtubule-associated protein tau gene (MAPT) mutations: a chameleon for neuropathology and neuroimaging. Neuropathol. Appl. Neurobiol. 41, 24–46 (2015).
Shi, Y. et al. Cryo-EM structures of tau filaments from Alzheimer’s disease with PET ligand APN-1607. Acta Neuropathol. 141, 697–708 (2021).
Steele, J. C., Richardson, J. C. & Olszewski, J. Progressive supranuclear palsy. a heterogeneous degeneration involving the brainstem, basal ganglia and cerebellum with vertical gaze and pseudobulbar palsy, nuchal dystonia and dementia. Arch. Neurol. 10, 333–359 (1964).
Williams, D. R. et al. Characteristics of two distinct clinical phenotypes in pathologically proven progressive supranuclear palsy: Richardson’s syndrome and PSP-parkinsonism. Brain 128, 1247–1258 (2005).
Höglinger, G. U. et al. Clinical diagnosis of progressive supranuclear palsy: the Movement Disorder Society criteria. Mov. Disord. 32, 853–864 (2017).
Kovacs, G. G. et al. Distribution patterns of tau pathology in progressive supranuclear palsy. Acta Neuropathol. 140, 99–119 (2020).
Sanders, D. W. et al. Distinct tau prion strains propagate in cells and mice and define different tauopathies. Neuron 82, 1271–1288 (2014).
Molina, J. A. et al. Primary progressive aphasia with glial cytoplasmic inclusions. Eur. Neurol. 40, 71–77 (1998).
Ahmed, Z. et al. Globular glial tauopathies (GGT) presenting with motor neuron disease or frontotemporal dementia: an emerging group of 4-repeat tauopathies. Acta Neuropathol. 122, 415–428 (2011).
Ahmed, Z. et al. Globular glial tauopathies (GGT): consensus recommendations. Acta Neuropathol. 126, 537–544 (2013).
Clark, C. N. et al. Temporal variant frontotemporal dementia is associated with globular glial tauopathy. Cogn. Behav. Neurol. 28, 92–97 (2015).
Tanaka, H. et al. Globular glial tauopathy type II: clinicopathological study of two autopsy cases. Neuropathology 39, 111–119 (2019).
Fu, Y. J. et al. Sporadic, four-repeat tauopathy with frontotemporal lobar degeneration, Parkinsonism, and motor neuron disease: a distinct clinicopathological and biochemical disease entity. Acta Neuropathol. 120, 21–32 (2010).
Miki, Y., Mori, F., Hori, E., Kaimori, M. & Wakabayashi, K. Hippocampal sclerosis with four repeat tau-positive round inclusions in the dentate gyrus: a new type of four-repeat tauopathy. Acta Neuropathol. 117, 713–718 (2009).
Kovacs, G. G. et al. Tauopathy with hippocampal 4-repeat tau immunoreactive spherical inclusions: a report of three cases. Brain Pathol. 28, 274–283 (2018).
Borrego-Écija, S. et al. Tauopathy with hippocampal 4-repeat tau immunoreactive spherical inclusions in a patient with PSP. Brain Pathol. 28, 284–286 (2018).
Nakane, T. et al. Single-particle cryo-EM at atomic resolution. Nature 587, 152–156 (2020).
Braak, H. & Braak, E. Argyrophilic grains: characteristic pathology of cerebral cortex in cases of adult-onset dementia without Alzheimer changes. Neurosci. Lett. 76, 124–127 (1987).
Braak, H. & Braak, E. Cortical and subcortical argyrophilic grains characterize a disease associated with adult-onset dementia. Neuropathol. Appl. Neurobiol. 15, 13–26 (1989).
Tolnay, M. & Clavaguera, F. Argyrophilic grain disease: a late-onset dementia with distinctive features among tauopathies. Neuropathology 24, 269–283 (2004).
Saito, Y. et al. Staging of argyrophilic grains: an age-associated tauopathy. J. Neuropathol. Exp. Neurol. 63, 911–918 (2004).
Ferrer, I., Santpere, G. & Van Leeuwen, F. W. Argyrophilic grain disease. Brain 131, 1416–1432 (2008).
Kovacs, G. G. et al. Aging-related tau astrogliopathy (ARTAG): harmonized evaluation strategy. Acta Neuropathol. 131, 87–102 (2016).
Klotz, S. et al. Multiple system aging-related tau astrogliopathy with complex proteinopathy in an oligosymptomatic octogenarian. Neuropathology 41, 72–83 (2021).
Spillantini, M. G. et al. Familial multiple system tauopathy with presenile dementia: a disease with abundant neuronal and glial tau filaments. Proc. Natl Acad. Sci. USA 94, 4113–4118 (1997).
Hutton, M. et al. Association of missense and 5’-splice-site mutations in tau with the inherited dementia FTDP-17. Nature 393, 702–705 (1998).
Spillantini, M. G. et al. Mutation in the tau gene in familial multiple system tauopathy with presenile dementia. Proc. Natl Acad. Sci. USA 95, 7737–7741 (1998).
Forrest, S. L. et al. Retiring the term FTDP-17 as MAPT mutations are genetic forms of sporadic frontotemporal tauopathies. Brain 141, 521–534 (2018).
Arai, T. et al. Identification of amino-terminally cleaved tau fragments that distinguish progressive supranuclear palsy from corticobasal degeneration. Ann. Neurol. 55, 72–79 (2004).
Taniguchi-Watanabe, S. et al. Biochemical classification of tauopathies by immunoblot, protein sequence and mass spectrometric analyses of sarkosyl-insoluble and trypsin-resistant tau. Acta Neuropathol. 131, 267–280 (2016).
Miki, T. et al. Factors associated with development and distribution of granular/fuzzy astrocytes in neurodegenerative diseases. Brain Pathol. 30, 811–830 (2020).
Vidal, R. et al. A stop-codon mutation in the BRI gene associated with familial British dementia. Nature 399, 776–781 (1999).
Vidal, R. et al. A decamer duplication in the 3’ region of the BRI gene originates an amyloid peptide that is associated with dementia in a Danish kindred. Proc. Natl Acad. Sci. USA 97, 4920–4925 (2000).
Holton, J. L. et al. Regional distribution of amyloid-Bri deposition and its association with neurofibrillary degeneration in familial British dementia. Am. J. Pathol. 158, 515–526 (2001).
Holton, J. L. et al. Familial Danish dementia: a novel form of cerebral amyloidosis associated with deposition of both amyloid-Dan and amyloid-beta. J. Neuropathol. Exp. Neurol. 61, 254–267 (2002).
Dan, A. et al. Extensive deamidation at asparagine residue 279 accounts for weak immunoreactivity of tau with RD4 antibody in Alzheimer’s disease brain. Acta Neuropathol. Commun. 1, 54 (2013).
Duyckaerts, C. et al. PART is part of Alzheimer disease. Acta Neuropathol. 129, 749–756 (2015).
Hallihan, G. I. et al. Structure of tau filaments in prion protein amyloidoses. Acta Neuropathol. 142, 227–241 (2021).
Kametani, F. et al. Comparison of common and disease-specific post-translational modifications of pathological tau associated with a wide range of tauopathies. Front. Neurosci. 14, 581936.
Grinberg, L. T. et al. Argyrophilic grain disease differs from other tauopathies by lacking tau acetylation. Acta Neuropathol. 125, 581–593 (2013).
Kovacs, G. G. et al. MAPT S305I mutation: implications for argyrophilic grain disease. Acta Neuropathol. 116, 103–118 (2008).
Rönnbäck, A. et al. Neuropathological characterization of two siblings carrying the MAPT S305S mutation demonstrates features resembling argyrophilic grain disease. Acta Neuropathol. 127, 297–298 (2014).
Varani, L. et al. Structure of tau exon 10 splicing regulatory element RNA and destabilization by mutations of frontotemporal dementia and parkinsonism linked to chromosome 17. Proc. Natl Acad. Sci. USA 96, 8229–8234 (1999).
Poorkaj, P. et al. Tau is a candidate gene for chromosome 17 frontotemporal dementia. Ann. Neurol. 43, 815–825 (1998).
Bugiani, O. et al. Frontotemporal dementia and corticobasal degeneration in a family with a P301S mutation in tau. J. Neuropathol. Exp. Neurol. 58, 667–677 (1999).
Lladó, A. et al. A novel MAPT mutation (P301T) associated with familial frontotemporal dementia. Eur. J. Neurol. 14, e9–e10 (2007).
Lewis, J. et al. Neurofibrillary tangles, amyotrophy and progressive motor disturbance in mice expressing mutant (P301L) tau protein. Nat. Genet. 25, 402–405 (2000).
Allen, B. et al. Abundant tau filaments and nonapoptotic neurodegeneration in transgenic mice expressing human P301S tau protein. J. Neurosci. 22, 9340–9351 (2002).
Coughlin, D. G., Dickson, D. W., Josephs, K. A. & Litvan, I. Progressive supranuclear palsy and corticobasal degeneration. Adv. Exp. Med. Biol. 1281, 151–176 (2021)
Josephs, K. A. et al. Atypical progressive supranuclear palsy with corticospinal tract degeneration. J. Neuropathol. Exp. Neurol. 65, 396–405 (2006).
Clavaguera, F. et al. Brain homogenates from human tauopathies induce tau inclusions in mouse brain. Proc. Natl Acad. Sci. USA 110, 9535–9540 (2013).
Evans, W. et al. The tau H2 haplotype is almost exclusively Caucasian in origin. Neurosci. Lett. 369, 183–185 (2004).
Farlow, J. L. et al. Whole-exome sequencing in familial Parkinson’s disease. JAMA Neurol. 73, 68–75 (2016).
Spina, S. et al. The tauopathy associated with mutation +3 in intron 10 of Tau: characterization of the MSTD family. Brain 131, 72–89 (2008).
Braak, H., Braak, E., Ohm, T. & Bohl, J. Silver impregnation of Alzheimer’s neurofibrillary changes counterstained for basophilic material and lipofuscin pigment. Stain Technol. 69, 197–200 (1988).
Guo, H. et al. Electron-event representation data enable efficient cryoEM file storage with full preservation of spatial and temporal resolution. IUCrJ 7, 860–869 (2020).
Zivanov, J. et al. A Bayesian approach to beam-induced motion correction in cryo-EM single-particle analysis. IUCrJ 6, 5–17 (2020).
Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221.
He, S. & Scheres, S. H. W. Helical reconstruction in RELION. J. Struct. Biol. 198, 163–176 (2017).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. Elife 7, e42166 (2018).
Wagner, T. et al. SPHIRE-crYOLO is a fast and accurate fully automated particle picker for cryo-EM. Commun. Biol. 2, 218 (2019).
Scheres, S. H. W. Amyloid structure determination in RELION-3.1. Acta Crystallogr. D 76, 94–101 (2020).
Zivanov, J. et al. Estimation of high-order aberrations and anisotropic magnification from cryo-EM data sets in RELION-3.1. IUCrJ 7, 253–267 (2020).
Chen, S. et al. High-resolution noise substitution to measure overfitting and validate resolution in 3D structure determination by single particle electron cryomicroscopy. Ultramicroscopy 135, 24–35 (2013).
Casañal, A. et al. Current developments in Coot for macromolecular model building of electron cryo-microscopy and crystallographic data. Protein Sci. 29, 1069–1078 (2020).
Williams, C. J. et al. MolProbity: more and better reference data for improved all-atom structure validation. Protein Sci. 27, 293–315 (2018).
Brown, A. et al. Tools for macromolecular model building and refinement into electron cryo-microscopy reconstructions. Acta Crystallogr. D 71, 136–153 (2015).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D 74, 531–544 (2018).
Pettersen, E. F. et al. UCSF ChimeraX: structure visualization for researchers, educators, and developers. Protein Sci. 30, 70–82 (2021).
Acknowledgements
We thank the families of the patients for donating brain tissues; U. Kuederli, M. Jacobsen, F. Epperson and R. M. Richardson for human brain collection and technical support; E. Gelpi for preparing brain samples from the ARTAG case; E. de Jong, B. van Knippenberg, L. Yu and E. Ioannou for support with the Krios G4 microscope; T. Darling and J. Grimmett for help with high-performance computing; and S. Lövestam, T. Nakane, R. A. Crowther, F. Clavaguera, K. Del Tredici, H. Braak, Z. Ahmed and M. G. Spillantini for helpful discussions. We acknowledge Diamond Light Source for access to and support with the cryo-EM facilities at the UK’s national Electron Bio-imaging Centre (eBIC) (under proposals BI23268-49 and BI23268-19), funded by the Wellcome Trust, Medical Research Council (MRC) and Biotechnology and Biological Sciences Research Council (BBSRC). This study was supported by the MRC Laboratory of Molecular Biology Electron Microscopy facility. W.Z. was supported by a foundation that prefers to remain anonymous. M.G. is an Honorary Professor in the Department of Clinical Neurosciences of the University of Cambridge. This work was supported by the UK MRC (MC_U105184291 to M.G. and MC_UP_A025_1013 to S.H.W.S.); the EU/EFPIA/Innovative Medicines Initiative (2) Joint Undertaking IMPRiND project 116060 to M.G.; the Japan Agency for Science and Technology (CREST) (JPMJCR18H3 to M.H.); the Japan Agency for Medical Research and Development (AMED) (JP20dm0207072 to M.H. and JP21dk0207045 and JP21ek0109545 to T.I.); JP20ek0109392, JP20ek0109391 and an Intramural Research Grant (number 30–8) for Neurological and Psychiatric Disorders of NCNP to M.Y.; the US National Institutes of Health (P30-AG010133, UO1-NS110437 and RF1-AG071177) to R.V., B.G. R.V. and B.G. were supported by the Department of Pathology and Laboratory Medicine, Indiana University School of Medicine. G.G.K. was supported by the Safra Foundation and the Rossy Foundation. T.R. is supported by the National Institute for Health Research Queen Square Biomedical Research Unit in Dementia. T.L. holds an Alzheimer’s Research UK Senior Fellowship. The Queen Square Brain Bank is supported by the Reta Lila Weston Institute for Neurological Studies.
Author information
Authors and Affiliations
Contributions
T.L., Y. Saito, S.M., M.Y., H.T., A.K., A.C.R., D.M.A.M., G.G.K., T.R. and B.G. identified patients and performed neuropathology. R.V., H.J.G., G.I.H. and T.I. performed genetic analysis. A.T., F.K., M.H., Y. Shi, W.Z., Y.Y. and B.F. prepared tau filament samples and performed biochemical analyses. Y. Shi, W.Z., Y.Y, B.F., A.K. and M.v.B. performed cryo-EM data acquisition. Y. Shi, W.Z., Y.Y., A.G.M. and S.H.W.S. performed cryo-EM structure determination. M.G. and S.H.W.S. supervised the project. All authors contributed to writing the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Gil Rabinovici, Henning Stahlberg and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
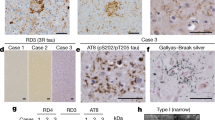
Extended Data Fig. 1 Tau immunohistochemistry.
Representative tau staining of the brain regions used for cryo-EM structure determination (see Methods), using antibody AT8 (pS202/pT205 tau). Scale bars are 50 μm, except for GGT Type I, FBD and FDD, for which they are 25 μm. For PSP-RS case 3, both the thalamus (Tha) and the entorhinal cortex (EC) are shown. Similar results were obtained using a minimum of six additional stained sections for each case.
Extended Data Fig. 2 Cryo-EM reconstructions.
a, Cryo-EM maps for tau filaments from six cases of PSP. For each map, a sum of the reconstructed density for several XY-slices is shown, corresponding to approximately 4.7 Å. The disease cases are referenced at the bottom of each image, the filament types at the top left and the percentages of a given filament type among the tau filaments in the dataset at the top right. Scale bar 5 nm. The same scale applies to all panels, except d–g. b–h, As in a, but a case of GGT-I and a case of GGT-II (b); a case diagnosed as PSP-F, but that contains filaments with the PGT fold (c); two cases of AGD (d); scale bar 5nm, the same scale applies to panels d–g; entorhinal cortex of PSP-RS case 3 (e); a case of ARTAG (f); three cases with mutations +16 or +3 in intron 10 of MAPT (g); a case of FBD and a case of FDD (h). Panels d–f contain blank squares to indicate the absence of AGD type I filaments in some cases. The inset with dashed lines shows 2D class average images of tau filaments from a case of GGT-III without apparent twist.
Extended Data Fig. 3 Cryo-EM resolution estimates.
Fourier Shell Correlation (FSC) curves for cryo-EM maps and atomic structures of PSP filaments (from PSP-RS case 1); GGT filament types 1-3 (from GGT-I); GPT filament types 1a, 1b and 2 (from PSP-F case 2); and AGD filament types 1 and 2 (from AGD case 1 and the +16 case, repectively). FSC curves are shown for two independently refined cryo-EM half-maps (black); for the final refined atomic model against the final cryo-EM map (red); for the atomic model refined in the first half-map against that half-map (blue); and for the refined atomic model in the first half-map against the other half-map (yellow).
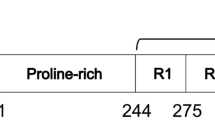
Extended Data Fig. 4 Schematics of tau filament folds.
a–d, Schematics of the tau folds for PSP (a), GGT (b), GPT (c) and AGD (d). Negatively charged residues are shown in red, positively charged residues in blue, polar residues in green, apolar residues in white, sulfur-containing residues in yellow, prolines in purple and glycines in pink. Thick connecting lines with arrow heads are used to indicate β-strands; additional densities are shown in grey.
Extended Data Fig. 5 Tau pathology in limbic-predominant neuronal inclusion body 4R tauopathy (LNT, PSP-F case 2).
a, Low power view of hippocampus stained with antibody AT8 (pS202/pT205tau). b, Low power view of frontal cortex stained with AT8. c, Higher power view of hippocampus stained with AT8. d, AT8-positive globular astrocyte in hippocampus. e, AT8-positive neurons in hippocampus. f, Hippocampus stained with antibody RD4 (specific for 4R tau). g, Gallyas-Braak silver-positive neurons and glial cells in hippocampus. h, Hippocampus stained with antibody RD3 (specific for 3R tau). i, Higher-power view of frontal cortex stained with AT8. j, AT8-positive globular astrocyte in frontal cortex. k, AT8-positive neurons in frontal cortex. l, Frontal cortex stained with RD4. m, Gallyas-Braak silver-positive neurons and glial cells in frontal cortex. n, Frontal cortex stained with RD3. Representative images are shown. Similar results were obtained using a minimum of six additional stained sections for each panel. Scale bars: 400 μm in (a); 200 μm in (b), 50 μm in (c, f, g, h, i, l, m, n); 10 μm in (d, e, j, k).
Extended Data Fig. 6 Structural comparisons.
a, Comparison of two different main-chain conformations for GPT type 1 filaments (type 1a in purple; type 1b in green) and the main-chain conformation of GPT type 2 filaments (red). b, Comparison of the PSP (orange), GGT (blue) and GPT (purple) folds. c, Comparison of the inter-protofilament interfaces of GGT type 3 and GPT type 2 filaments. d, Comparison of the AGD type 1 (light blue), AGD type 2 (pink) and CBD (grey) folds.
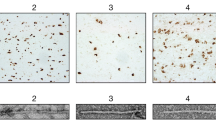
Extended Data Fig. 7 Argyrophilic grains in the entorhinal cortex.
Representative tau staining with antibodies RD4 (4R tau), RD3 (3R tau), AT8 (pS202/pT205 tau), as well as Gallyas-Braak silver, of the entorhinal cortex from cases 1 and 2 with mutation +3 in intron 10 of MAPT and from case 3 of PSP-RS. Similar results were obtained using a minimum of six additional stained sections for each case. Scale bar, 50 μm.
Extended Data Fig. 8 Immunoblot analysis of 4R tauopathies.
Hyperphosphorylated full-length tau (64 and 68 kDa) and C-terminal fragments (33 kDa and 37 kDa) were detected in sarkosyl-insoluble fractions from the brain regions used for cryo-EM by anti-tau antibody T46. A prominent 33-kDa band was characteristic of PSP and GGT; strong 37-kDa bands were in evidence in AGD, ARTAG, cases with intron 10 mutations in MAPT (+3 and +16) and in CBD. PSP-RS case 3 had a strong 33-kDa band in thalamus (Tha) and strong 37-kDa bands in entorhinal cortex (EC), consistent with AGD co-pathology. Similar results were obtained in three independent experiments. The original, uncropped image is available in Supplementary Fig. 1.
Supplementary information
Supplementary Information
This file contains Supplementary Tables 1–3 and Supplementary Figure 1.
Rights and permissions
About this article

Cite this article
Shi, Y., Zhang, W., Yang, Y. et al. Structure-based classification of tauopathies. Nature 598, 359–363 (2021). https://doi.org/10.1038/s41586-021-03911-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-021-03911-7
This article is cited by
-
Brain clearance of protein aggregates: a close-up on astrocytes
Molecular Neurodegeneration (2024)
-
Shaping the future of preclinical development of successful disease-modifying drugs against Alzheimer's disease: a systematic review of tau propagation models
Acta Neuropathologica Communications (2024)
-
Misfolded protein oligomers: mechanisms of formation, cytotoxic effects, and pharmacological approaches against protein misfolding diseases
Molecular Neurodegeneration (2024)
-
Astrocytic accumulation of tau fibrils isolated from Alzheimer’s disease brains induces inflammation, cell-to-cell propagation and neuronal impairment
Acta Neuropathologica Communications (2024)
-
Synthetic β-sheets mimicking fibrillar and oligomeric structures for evaluation of spectral X-ray scattering technique for biomarker quantification
Cell & Bioscience (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
