
Abstract
The role of gene expression during learning and in short-term memories has been studied extensively1,2,3, but less is known about remote memories, which can persist for a lifetime4. Here we used long-term contextual fear memory as a paradigm to probe the single-cell gene expression landscape that underlies remote memory storage in the medial prefrontal cortex. We found persistent activity-specific transcriptional alterations in diverse populations of neurons that lasted for weeks after fear learning. Out of a vast plasticity-coding space, we identified genes associated with membrane fusion that could have important roles in the maintenance of remote memory. Unexpectedly, astrocytes and microglia also acquired persistent gene expression signatures that were associated with remote memory, suggesting that they actively contribute to memory circuits. The discovery of gene expression programmes associated with remote memory engrams adds an important dimension of activity-dependent cellular states to existing brain taxonomy atlases and sheds light on the elusive mechanisms of remote memory storage.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Data availability
The accession number for the single-cell RNA sequencing data reported in this paper is GSE152632.
Code availability
Custom scripts can be found at https://github.com/mbchen-424/memory-sc-rnaseq.
References
Kandel, E. R. The molecular biology of memory storage: a dialogue between genes and synapses. Science 294, 1030–1038 (2001).
Flexner, L. B. & Flexner, J. B. Effect of acetoxycycloheximide and of an acetoxycycloheximide–puromycin mixture on cerebral protein synthesis and memory in mice. Proc. Natl Acad. Sci. USA 55, 369–374 (1966).
Alberini, C. M. & Kandel, E. R. The regulation of transcription in memory consolidation. Cold Spring Harb. Perspect. Biol. 7, a021741 (2014).
Squire, L. R. Mechanisms of memory. Science 232, 1612–1619 (1986).
Kitamura, T. et al. Engrams and circuits crucial for systems consolidation of a memory. Science 356, 73–78 (2017).
DeNardo, L. & Luo, L. Genetic strategies to access activated neurons. Curr. Opin. Neurobiol. 45, 121–129 (2017).
DeNardo, L. A. et al. Temporal evolution of cortical ensembles promoting remote memory retrieval. Nat. Neurosci. 22, 460–469 (2019).
McKenzie, S. & Eichenbaum, H. Consolidation and reconsolidation: two lives of memories? Neuron 71, 224–233 (2011).
Guenthner, C. J., Miyamichi, K., Yang, H. H., Heller, H. C. & Luo, L. Permanent genetic access to transiently active neurons via TRAP: targeted recombination in active populations. Neuron 78, 773–784 (2013).
Zeisel, A. et al. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 347, 1138–1142 (2015).
O’Sullivan, C. N., Sheridan, G. & Murphy, K. in Transcription Factors CREB and NF-κB: Involvement in Synaptic Plasticity and Memory Formation 43–65 (Bentham Science, 2012).
Suzuki, A. et al. Upregulation of CREB-mediated transcription enhances both short- and long-term memory. J. Neurosci. 31, 8786–8802 (2011).
Kida, S. et al. CREB required for the stability of new and reactivated fear memories. Nat. Neurosci. 5, 348–355 (2002).
Inuzuka, M., Hayakawa, M. & Ingi, T. Serinc, an activity-regulated protein family, incorporates serine into membrane lipid synthesis. J. Biol. Chem. 280, 35776–35783 (2005).
Zhang, X., Rizo, J. & Südhof, T. C. Mechanism of phospholipid binding by the C2A-domain of synaptotagmin I. Biochemistry 37, 12395–12403 (1998).
Bélanger, M., Allaman, I. & Magistretti, P. J. Brain energy metabolism: focus on astrocyte–neuron metabolic cooperation. Cell Metab. 14, 724–738 (2011).
Williamson, L. L., Sholar, P. W., Mistry, R. S., Smith, S. H. & Bilbo, S. D. Microglia and memory: modulation by early-life infection. J. Neurosci. 31, 15511–15521 (2011).
Ramilowski, J. A. et al. A draft network of ligand–receptor-mediated multicellular signalling in human. Nat. Commun. 6, 7866 (2015).
Südhof, T. C. Synaptic neurexin complexes: a molecular code for the logic of neural circuits. Cell 171, 745–769 (2017).
Saunders, A. et al. Molecular diversity and specializations among the cells of the adult mouse brain. Cell 174, 1015–1030.e16 (2018).
Lacar, B. et al. Nuclear RNA-seq of single neurons reveals molecular signatures of activation. Nat. Commun. 7, 11022 (2016).
Cho, J.-H., Huang, B. S. & Gray, J. M. RNA sequencing from neural ensembles activated during fear conditioning in the mouse temporal association cortex. Sci. Rep. 6, 31753 (2016).
Hrvatin, S. et al. Single-cell analysis of experience-dependent transcriptomic states in the mouse visual cortex. Nat. Neurosci. 21, 120–129 (2018).
Rao-Ruiz, P. et al. Engram-specific transcriptome profiling of contextual memory consolidation. Nat. Commun. 10, 2232 (2019).
Zhou, M. et al. A central amygdala to zona incerta projection is required for acquisition and remote recall of conditioned fear memory. Nat. Neurosci. 21, 1515–1519 (2018).
Picelli, S. et al. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protoc. 9, 171–181 (2014).
Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420 (2018).
Acknowledgements
We thank L. Chen, M. Zhou and J. Li for discussion of the experimental design; S. Kolluru and D. Henderson for assistance in library preparation; N. Neff and J. Okamoto for assistance with sequencing; J. Lui for advice on brain dissociation; L. Denardo, J. Lui and L. Luo for the gift and help with the TRAP2 line; and W. Wang, G. Stanley and F. Horns for helpful discussions and computational assistance. S.R.Q. is a Chan Zuckerberg Investigator. This work is supported by a grant from the NIH (MH115999 to T.C.S.)
Author information
Authors and Affiliations
Contributions
X.J. and T.C.S. designed the animal experiments. M.B.C. and S.R.Q. designed the single-cell RNA sequencing experiments. X.J. performed the animal experiments, brain dissection, and in situ hybridization and imaging. M.B.C. performed the brain dissociation, flow cytometry, single-cell library preparation and sequencing pipelines. M.B.C. performed all single-cell RNA sequencing data and image analysis, with input from X.J., S.R.Q. and T.C.S. M.B.C. wrote the manuscript with substantial contributions from X.J., S.R.Q. and T.C.S. T.C.S. and S.R.Q. oversaw the project.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Steve Ramirez and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Fidelity of the TRAP2; Ai14 line and sequencing quality metrics.
a, Fidelity with which the tdTomato reporter in TRAP2; Ai14 mice captures endogenous cFos expression during fear memory encoding by in situ hybridization of cFos and iCre in the mPFC directly following fear conditioning. Left, percentage of cFos+ cells in either fear conditioned (FC) or homecage (HC) mice as seen by in situ staining (mean ± s.d.). Percentage of cFos+ cells that are also iCre+(mean ± s.e.m). Right, representative in situ hybridization images of FC mice (mPFC). b, Representative images of regions of the mPFC analysed in this study (anterior cingulate cortex (ACC), the prelimbic (PrL) and the infralimbic cortex (IL)). Scale bars are 1mm (and 0.5 mm in the insets). c, Violin plots of the number of reads and number of genes per biological replicates (m1–m4), per cell. d, Scatterplot depicting the lack of strong relationship between number of genes detected and number of reads obtained per cell. e, Scaled expression of canonical markers in non-neuronal cells (Cldn5-BECs, Pdgfra-OPCs, Cx3cr1-microglia, Aqp4-astrocytes).
Extended Data Fig. 2 Distribution of cell numbers and neuronal subtypes across various training conditions.
a, Representative flow cytometry plot of the amount of tdT + events per training condition. In scatter plot, each point represents one mouse (mean ± s.d.). b, Number of cells from each biological replicate that were annotated as one of 7 defined neuron subtypes (C0–C6). c, Scaled expression of the top marker genes for each neuron subtype. d, Number of TRAPed and Inactive cells (as defined by non-zero expression of tdT mRNA) collected per neuron subtype, in either fear-recall (FR) or no-fear (NF) mice. e, Representative images of subtype marker genes (C0–C6) in the Allen Brain ISH atlas. f, Neuron subtype composition of FR and NF populations. Number of TRAPed (tdT mRNA+) cells collected in each experimental condition that fall in one of 7 neuronal subtype categories.
Extended Data Fig. 3 Differential gene expression in distinct neuronal subtypes (FR over NF TRAPed populations).
a, DEGs in each neuron subtype when inactive (tdT-) neurons are compared between FR and NF mice. b, Left, volcano plots of DEGs in FR vs NF mice for each neuron subtype (C0 to C4). DEGs found when all replicates are pooled analysed in a combined manner are shown in red. Recall-dependent DEGs (defined as being differentially expressed in at least 3 out of 4 replicates, when analysed replicates are analysed individually) are labelled in black. Right, permutations are performed for every recall-dependent DEG for each neuronal population. Upregulated DEGs lying above the y = x line (red) and downregulated DEGs lying below the y = x line (blue) are considered to be above the 99th percentile of the permuted distribution. c, GO enrichment analysis of all up- and downregulated DEGs (941 DEGs up, 384 DEGs down, all neuron subtypes combined) when all replicates are pooled. Bars show the enrichment scores (GeneAnalytics) for the GO pathway and dot indicates the number of DEGs involved in that pathway. d, GO enrichment analysis of only the upregulated remote-memory-specific DEGs (from Fig. 3c) from all neuron subtypes combined. Bar indicates enrichment score (Gene Analytics) and number indicates number of recall-dependent DEGs involved in that pathway.
Extended Data Fig. 4 Analysis of TRAPed ensembles in food salience (S) versus no salience (NS) mice.
a, Schematic of experimental paradigm for generating TRAPed neuronal ensembles as a result of food salience (food deprivation followed by food return (salience) or no food return (no salience)). b, Percentage of events in flow cytometry that were tdT +, by experience (mean ± s.d.). c, tSNE of the merge of data from fear-recall experiments and food-salience experiments coloured by neuron subtype (left) and experimental paradigm (right). d, Subtype composition differences between TRAPed fear-recalled ensembles and TRAPed food salience ensembles, as compared to background Inactive ensembles. e, Heat map of the average log2 fold change of DEGs in each neuron subtype when comparing fear-recall vs no-fear, and salience vs no salience. Only DEGs with FDR <0.01 are shown.
Extended Data Fig. 5 DEGs when comparing ensembles from food salience (S) to no salience (NS) mice.
Volcano plots showing the log2 fold change and adjusted P values (in log10 scale) of genes when comparing food salience over no salience groups within each neuron subtype (C0 through C4). Top DEGs per neuron subtype are labelled in red. Positive log2 fold change indicates upregulation in food salience (S) group.
Extended Data Fig. 6 Neuron subtype-specific activation programs, hypothesized protein–protein interactions and upstream regulatory motifs.
a, Fraction of cells in each neuron subtype that are induced with the transcriptional program (that is, DEGs) from a neuron subtype. Overall, the activation program of each TRAPed neuron subtype is found to be more specific to it than the inactive population, or other subtypes. b, Left, de novo regulator motif discovery: analysis was performed using HOMER on the subset of 99 remote-memory-associated DEGs by looking at the sequences -400 to +100 bp from the TSS. 12 de novo and 2 known motifs were found (only motifs with an enrichment P value <10−2) were kept). Heat map depicts the ‘motif score’ of each DEG for each motif, and genes and motifs were clustered via the ward.D method. Right, bar graph depicting the percentage of the DEGs (target sequences) that possess a match for the motif within -400 to +100 bp from the TSS, vs the percentage of background sequences. For de novo motifs, the best match gene is listed on the right. HIF1b and HIF1a are matches to known motifs. c, Left, hypothesized protein–protein interactions of a subset of recall-dependent DEGs (TRAPed FR/NF) using the STRING database (https://string-db.org/). Only genes that are connected at a confidence level of 0.4 (medium) are shown. Connections indicate a possible existence of an interaction between two proteins. Genes are coloured by up of downregulation in FR/NF. Right, same network plot, with nodes coloured by the neuron subtype which differentially regulates the DEG.
Extended Data Fig. 7 In situ validation of tdT levels, neuronal subtype compositions and remote-memory-specific DEGs in the mPFC.
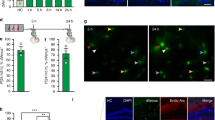
a, Ratio of Nuclei that are tdT+ (mRNA level) per training condition. Each data point represents one region of interest. (mean ± s.d.) b, Ratio of TRAPed cells that are positive for a neuronal subtype marker obtained either via the RNA-scope method, or by scRNA-seq (mean ± s.e.m.) (see Fig. 2). TRAPed cells are defined as DAPI+/tdT+ in RNAscope quantification, and as tdT mRNA count >1 in scRNA-seq (post-QC). No significant differences are found between FR and NF within either RNAscope or scRNA-methods, indicating no major changes in neuronal subtype composition of active populations in different training conditions. c, in situ hybridization of key remote-memory specific DEGs including Stx1b in Rprm+/tdT + cells, Syt13 in Tnfaip8l3+/tdT + cells, Vamp2 in Tesc+/tdT + cells. Scale bars, 100 μm.
Extended Data Fig. 8 DEGs and potential cell–cell interactions in non-neuronal cells during memory consolidation.
a, Volcano plots of non-neuronal cell types when comparing cells in FR over NF nice. DEGs (FDR >0.01, log2FC >1) are labelled in red, and exemplary DEGs (high log2FC and log10FDR) are labelled in black. b, Number of non-neuronal cells collected in this study, for each cell type and experimental condition. c, Heat map of a subset of neuronal ligands and glial receptors that are found to be differentially perturbed upon memory consolidation. Only receptors and ligands which were found to be (differentially) expressed are shown. d, Left, heat map of the log2FC of DEGs (FR over NF) in neurons that are classified as ligands. Middle and right, Sankey plot of known ligand-receptor pairs and heat map of the average scaled expression level of the corresponding receptors in each cell type. e, Left, heat map of the log2FC of DEGs (FR over NF) in neurons that are classified as receptors. Middle and right, Sankey plot of known ligand-receptor pairs and heat map of the average scaled expression level of the corresponding ligands in each cell type.
Extended Data Fig. 9 Comparison of remote-memory DEGs with previously published datasets of experience-dependent transcriptional activity.
a, Left, heat map of the log2FC of all 1,292 DEGs (FDR <0.01, FR over NF, all cells pooled) in this manuscript, and their log2FC values in previously published datasets of experience-dependent DEGs in activated neurons during: recent fear memory retrieval24, associative fear-learning22, post-visual stimulus23, or novel environment exposure21. A value of zero log2FC indicates the gene was not differentially expressed in a dataset. Right, same as left, but now DEGs are filtered down to the ‘Recall-dependent DEG’ set derived from this manuscript. Only genes differentially expressed in three out of four replicates are remaining. b, Left, log2 fold change heat map of the recall-dependent DEGs between tdT + vs tdT − neurons in FR mice (genes are differentially expressed in >3/4 replicates) undergoing remote fear memory consolidation. Right, the log2FC values of these genes if they are found in previously published datasets of experience-dependent DEGs (see a). A value of zero log2FC indicates the gene was not differentially expressed in that dataset.
Extended Data Fig. 10 Comparison of remote-memory-specific DEGs and fear-experience-related DEGs.
Left, log2 fold change heatmap of the union of DEGs from comparing FR vs NF (remote-memory specific) and NR vs NF (fear-related). Sustained transcriptional changes from the fear-experience itself is shown in the yellow highlighted columns. Right, zoomed in view of the portion of the heat map within the green boxes where most fear experience-related DEGs are located.
Supplementary information
Rights and permissions
About this article

Cite this article
Chen, M.B., Jiang, X., Quake, S.R. et al. Persistent transcriptional programmes are associated with remote memory. Nature 587, 437–442 (2020). https://doi.org/10.1038/s41586-020-2905-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2905-5
This article is cited by
-
q-Diffusion leverages the full dimensionality of gene coexpression in single-cell transcriptomics
Communications Biology (2024)
-
Alternative splicing of latrophilin-3 controls synapse formation
Nature (2024)
-
Context memory formed in medial prefrontal cortex during infancy enhances learning in adulthood
Nature Communications (2024)
-
Spatial transcriptomics reveal neuron–astrocyte synergy in long-term memory
Nature (2024)
-
Noteworthy perspectives on microglia in neuropsychiatric disorders
Journal of Neuroinflammation (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
