
Abstract
Elucidating elementary mechanisms that underlie bacterial diversity is central to ecology1,2 and microbiome research3. Bacteria are known to coexist by metabolic specialization4, cooperation5 and cyclic warfare6,7,8. Many species are also motile9, which is studied in terms of mechanism10,11, benefit12,13, strategy14,15, evolution16,17 and ecology18,19. Indeed, bacteria often compete for nutrient patches that become available periodically or by random disturbances2,20,21. However, the role of bacterial motility in coexistence remains unexplored experimentally. Here we show that—for mixed bacterial populations that colonize nutrient patches—either population outcompetes the other when low in relative abundance. This inversion of the competitive hierarchy is caused by active segregation and spatial exclusion within the patch: a small fast-moving population can outcompete a large fast-growing population by impeding its migration into the patch, while a small fast-growing population can outcompete a large fast-moving population by expelling it from the initial contact area. The resulting spatial segregation is lost for weak growth–migration trade-offs and a lack of virgin space, but is robust to population ratio, density and chemotactic ability, and is observed in both laboratory and wild strains. These findings show that motility differences and their trade-offs with growth are sufficient to promote diversity, and suggest previously undescribed roles for motility in niche formation and collective expulsion–containment strategies beyond individual search and survival.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
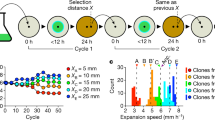


Data availability
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Code availability
Custom-written scripts used in this study are available from the corresponding authors upon reasonable request.
References
Rainey, P. B., Buckling, A., Kassen, R. & Travisano, M. The emergence and maintenance of diversity: insights from experimental bacterial populations. Trends Ecol. Evol. 15, 243–247 (2000).
Hibbing, M. E., Fuqua, C., Parsek, M. R. & Peterson, S. B. Bacterial competition: surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 8, 15–25 (2010).
Koskella, B., Hall, L. J. & Metcalf, C. J. E. The microbiome beyond the horizon of ecological and evolutionary theory. Nat. Ecol. Evol. 1, 1606–1615 (2017).
Turner, P. E., Souza, V. & Lenski, R. E. Tests of ecological mechanisms promoting the stable coexistence of two bacterial genotypes. Ecology 77, 2119–2129 (1996).
Ross-Gillespie, A., Gardner, A., West, S. A. & Griffin, A. S. Frequency dependence and cooperation: theory and a test with bacteria. Am. Nat. 170, 331–342 (2007).
Kerr, B., Riley, M. A., Feldman, M. W. & Bohannan, B. J. Local dispersal promotes biodiversity in a real-life game of rock–paper–scissors. Nature 418, 171–174 (2002).
Reichenbach, T., Mobilia, M. & Frey, E. Mobility promotes and jeopardizes biodiversity in rock–paper–scissors games. Nature 448, 1046–1049 (2007).
Kelsic, E. D., Zhao, J., Vetsigian, K. & Kishony, R. Counteraction of antibiotic production and degradation stabilizes microbial communities. Nature 521, 516–519 (2015).
Berg, H. C. Motile behavior of bacteria. Phys. Today 53, 24–29 (2000).
Parkinson, J. S., Hazelbauer, G. L. & Falke, J. J. Signaling and sensory adaptation in Escherichia coli chemoreceptors: 2015 update. Trends Microbiol. 23, 257–266 (2015).
Alon, U., Surette, M. G., Barkai, N. & Leibler, S. Robustness in bacterial chemotaxis. Nature 397, 168–171 (1999).
Ames, P. & Bergman, K. Competitive advantage provided by bacterial motility in the formation of nodules by Rhizobium meliloti. J. Bacteriol. 148, 728–729 (1981).
Stocker, R., Seymour, J. R., Samadani, A., Hunt, D. E. & Polz, M. F. Rapid chemotactic response enables marine bacteria to exploit ephemeral microscale nutrient patches. Proc. Natl Acad. Sci. USA 105, 4209–4214 (2008).
Roszak, D. B. & Colwell, R. R. Survival strategies of bacteria in the natural environment. Microbiol. Rev. 51, 365–379 (1987).
Vergassola, M., Villermaux, E. & Shraiman, B. I. ‘Infotaxis’ as a strategy for searching without gradients. Nature 445, 406–409 (2007).
Fraebel, D. T. et al. Environment determines evolutionary trajectory in a constrained phenotypic space. eLife 6, e24669 (2017).
Yi, X. & Dean, A. M. Phenotypic plasticity as an adaptation to a functional trade-off. eLife 5, e19307 (2016).
Grossart, H.-P., Riemann, L. & Azam, F. Bacterial motility in the sea and its ecological implications. Aquat. Microb. Ecol. 25, 247–258 (2001).
Song, H., Payne, S., Gray, M. & You, L. Spatiotemporal modulation of biodiversity in a synthetic chemical-mediated ecosystem. Nat. Chem. Biol. 5, 929–935 (2009).
Lehman, J. T. & Scavia, D. Microscale patchiness of nutrients in plankton communities. Science 216, 729–730 (1982).
Blackburn, N., Fenchel, T. & Mitchell, J. Microscale nutrient patches in planktonic habitats shown by chemotactic bacteria. Science 282, 2254–2256 (1998).
Ochman, H. & Selander, R. K. Standard reference strains of Escherichia coli from natural populations. J. Bacteriol. 157, 690–693 (1984).
Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 31, 343–366 (2000).
Jeltsch, F. et al. Integrating movement ecology with biodiversity research - exploring new avenues to address spatiotemporal biodiversity dynamics. Mov. Ecol. 1, 6 (2013).
Liu, J. T. et al. Metabolic co-dependence gives rise to collective oscillations within biofilms. Nature 523, 550–554 (2015).
Miller, M. B. & Bassler, B. L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 55, 165–199 (2001).
Hallatschek, O., Hersen, P., Ramanathan, S. & Nelson, D. R. Genetic drift at expanding frontiers promotes gene segregation. Proc. Natl Acad. Sci. USA 104, 19926–19930 (2007).
Keller, E. F. & Segel, L. A. Traveling bands of chemotactic bacteria: a theoretical analysis. J. Theor. Biol. 30, 235–248 (1971).
Houchmandzadeh, B. & Vallade, M. Fisher waves: an individual-based stochastic model. Phys. Rev. E 96, 012414 (2017).
Gandhi, S. R., Yurtsev, E. A., Korolev, K. S. & Gore, J. Range expansions transition from pulled to pushed waves as growth becomes more cooperative in an experimental microbial population. Proc. Natl Acad. Sci. USA 113, 6922–6927 (2016).
Ni, B. et al. Evolutionary remodeling of bacterial motility checkpoint control. Cell Rep. 18, 866–877 (2017).
Dobell, C. Antony Van Leeuwenhoek and His Little Animals (John Bale, Sons & Danielsson, 1960).
Levin, S. A. Dispersion and population interactions. Am. Nat. 108, 207–228 (1974).
Sousa, W. P. The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 15, 353–391 (1984).
Taute, K. M., Gude, S., Tans, S. J. & Shimizu, T. S. High-throughput 3D tracking of bacteria on a standard phase contrast microscope. Nat. Commun. 6, 8776 (2015).
Berg, H. C. & Brown, D. A. Chemotaxis in Escherichia coli analyzed by three-dimensional tracking. Antibiot. Chemother. 19, 55–78 (1974).
Boyd, S., Parikh, N., Chu, E., Peleato, B. & Eckstein, J. Distributed optimization and statistical learning via the alternative direction method of multipliers. Found. Trends Mach. Learn. 3, 1–122 (2011).
Leifson, E. Staining, shape and arrangement of bacterial flagella. J. Bacteriol. 62, 377–389 (1951).
Schneider, C. A., Rasband, W. S., & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Edelstein, A. D. et al. Advanced methods of microscope control using μManager software. J. Biol. Methods 1, e10 (2014).
Leinonen, R., Sugawara, H., Shumway, M. & The International Nucleotide Sequence Database Collaboration. The Sequence Read Archive. Nucleic Acids Res. 39, D19–D21 (2010).
Joshi, N. A. & Fass, J. N. Sickle: a sliding-window, adaptive, quality-based trimming tool for FastQ files. v.1.33 https://github.com/najoshi/sickle (2011).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
McKenna, A. et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303 (2010).
Cingolani, P. et al. Using Drosophila melanogaster as a model for genotoxic chemical mutational studies with a new program, SnpSift. Front. Genet. 3, 35 (2012).
Danecek, P. et al. The variant call format and VCFtools. Bioinformatics 27, 2156–2158 (2011).
Cingolani, P. et al. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 6, 80–92 (2012).
Python Software Foundation. Python v.3.6.7 http://www.python.org (2018).
Cock, P. J. et al. Biopython: freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 25, 1422–1423 (2009).
Monod, J. The growth of bacterial cultures. Annu. Rev. Microbiol. 3, 371–394 (1949).
Senn, H., Lendenmann, U., Snozzi, M., Hamer, G. & Egli, T. The growth of Escherichia coli in glucose-limited chemostat cultures: a re-examination of the kinetics. Biochim. Biophys. Acta 1201, 424–436 (1994).
Hazel, J. R. & Sidell, B. D. A method for the determination of diffusion coefficients for small molecules in aqueous solution. Anal. Biochem. 166, 335–341 (1987).
Parkinson, J. S. Complementation analysis and deletion mapping of Escherichia coli mutants defective in chemotaxis. J. Bacteriol. 135, 45–53 (1978).
Sourjik, V. & Berg, H. C. Localization of components of the chemotaxis machinery of Escherichia coli using fluorescent protein fusions. Mol. Microbiol. 37, 740–751 (2000).
Zinder, N. D. & Lederberg, J. Genetic exchange in Salmonella. J. Bacteriol. 64, 679–699 (1952).
Zhou, Q., Ames, P., & Parkinson, J. S. Biphasic control logic of HAMP domain signalling in the Escherichia coli serine chemoreceptor. Mol. Microbiol. 80, 596–611 (2011).
Turner, L., Ryu, W. S. & Berg, H. C. Real-time imaging of fluorescent flagellar filaments. J. Bacteriol. 182, 2793–2801 (2000).
Yamashita, I. et al. Structure and switching of bacterial flagellar filaments studied by X-ray fiber diffraction. Nat. Struct. Biol. 5, 125–132 (1998).
Lynch, M. & Marinov, G. K. The bioenergetic costs of a gene. Proc. Natl Acad. Sci. USA 112, 15690–15695 (2015).
Tran, Q. H. & Unden, G. Changes in the proton potential and the cellular energetics of Escherichia coli during growth by aerobic and anaerobic respiration or by fermentation. Eur. J. Biochem. 251, 538–543 (1998).
Ryu, W. S., Berry, R. M. & Berg, H. C. Torque-generating units of the flagellar motor of Escherichia coli have a high duty ratio. Nature 403, 444–447 (2000).
Bremer, H. & Dennis, P. P. in Escherichia coli and Salmonella: Cellular and Molecular Biology (ed. Neidhardt, F. C.) Ch. 97 (ASM, 1996).
Acknowledgements
Work in the groups of S.J.T. and T.S.S. is supported by the Netherlands Organization for Scientific Research (NWO). T.S.S. also acknowledges partial support by the Allen Distinguished Investigator Program (grant 11562) through the Paul G. Allen Frontiers Group. We thank H. C. Berg, J. S. Parkinson and V. Sourjik for providing strains and plasmids, V. Sunderlikova, S. Verstege-Boskamp and Z. Stachova Rychnavskafor for technical assistance, M. Lazova for early ECOR strain experiments, T. Averink and N. Anand for help with plasmid construction, F. Büke for his help with the plate-reader measurements, M. Akman for help with genomics analyses and P. R. ten Wolde, J. van Zon and S. Werner for reading the manuscript.
Author information
Authors and Affiliations
Contributions
S.G., K.M.T., T.S.S. and S.J.T. conceived the research. S.G., E.P., K.M.T., T.S.S. and S.J.T. designed the experiments. S.G., E.P. and A.-B.S. performed the competition experiments, S.G. performed the LAB strain characterizations, E.P. and S.G. performed the wild isolate characterization, E.P. performed the wild isolate three-dimensional tracking, S.G. and E.P. performed the fluorescence calibration measurements, S.G performed the secreted compound tests, K.M.T performed the motility cost and corresponding trade-off experiments. S.G., E.P. and K.M.T. analysed the data, S.G. performed the numerical simulations and sequence analysis. S.G., E.P., K.M.T., T.S.S. and S.J.T. contributed to writing the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Jeff Gore and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Competition and selection functions of cohabitant wild isolate strains.
a–e, Fold change of the population ratio R as a function of initial R0 for the five cohabitant pairs of the ECOR strain collection that could be transformed with plasmids for expression of fluorescent-protein labels. Out of the five pairs, three (ECOR18(A)–ECOR19(B) (a), ECOR32–ECOR68 (b) and ECOR36–ECOR8 (c)) clearly demonstrated a negative selection function, and one pair (ECOR36–ECOR8 (b)) showed, in addition, hierarchy inversion. The competition experiments were performed in TB soft-agar assays (see Methods). f, Scatter plot of growth propensity (OD600 at 7.5 h of growth in 96-well plate) versus migration rate for the five tested cohabitant pairs (same data as in Fig. 2b, reproduced here for convenience). g, Reciprocal spatial exclusion observed in cohabitant pairs of wild isolate strains. Tiled fluorescence micrographs show that out of the five cohabitant pairs tested, three (ECOR8–ECOR36, ECOR68–ECOR32 and ECOR18(A)–ECOR19(B)) exhibit spontaneous segregation into proximal and distal territories. The pairs were co-cultured in TB-based soft-agar gels with identical dimensions as plates shown at Fig. 2a, c–f (approximately 4 mm high, 35 mm in diameter). Scale bar, 7.5 mm.
Extended Data Fig. 2 Calibrated tile-scan fluorescence microscopy.
a, b, Calibration curves relating cell density (in OD600 (AU)) to fluorescence intensity was obtained by imaging soft-agar plates into which different known densities of cells were suspended in the molten agar before cooling to form the gel (see Methods). Calibration curves for TB (ECOR18(eYFP) (red) and ECOR19(eCFP) (blue)) (a) and MMH1 (LAB1(eYFP) (red) and LAB5(eCFP) (blue)) (b). Similar calibration curves were obtained for LAB1(eYFP) and LAB2(eCFP) in MMH1. c, Calibrated tile-scan fluorescence microscopy. Petri dishes (35 mm diameter) containing soft-agar gel medium were imaged by scanning an 11 × 11 grid of image fields (tiles) for each channel (YFP, CFP and bright field) at each time point. Image post-processing was performed for each tile individually. Subsequently, tile images were assembled into montages. Montages were converted and population size and colony front position were extracted.
Extended Data Fig. 3 Edge inoculation in minimal medium and colony expansion in rich medium.
a, Reciprocal spatial exclusion was confirmed in both centre- and edge-inoculated competitions in the soft-agar gel environment. ECOR18 and ECOR19 labelled with YFP and CFP, respectively (strains A and B in the main text) were pre-grown on TB and glycerol minimal medium as described in the Methods. The OD600 of both strains was adjusted to 10−2. Strains were mixed 1:1 and subsequently mixed with twofold concentrated molten agar. The final mixture was kept at room temperature for 1.5 h to transition into a gel state. The gel, containing a mixture of ECOR18 and ECOR19, was transferred onto soft-agar plates using a 1-ml pipette, either around the edge of the plate (plates 1–5 and 8) or into the centre (plates 6 and 7). All eight plates were placed at 33.5 °C in a humid environment for approximately 4 days and subsequently imaged by fluorescence microscopy. When inoculated around the edge, ECOR18 occupies the edge region, while ECOR19 is seen closer to the centre of the dish. Conversely, when inoculated in the centre ECOR18 occupies the central region of the dish, while ECOR19 is seen further away from the centre. Hence, spatial exclusion is seen for both centre and edge inoculation. Petri dish diameter: 35 mm. b, Migration phenotypes of all six cohabitant pairs (that is, isolated from the same host) in the ECOR strain collection on LB soft-agar plates at 37 °C. Petri dish diameter: 90 mm. The images of culture plates were acquired at different time points (see top-left inset of each sub-image). Scale bar, 25 mm.
Extended Data Fig. 4 Test for secreted compounds that inhibit growth.
a, Tile-scanned snapshots of Petri dishes (diameter 35 mm) in bright-field (BF) and fluorescence (CFP and YFP) after approximately 3 days. Images were not corrected for background or autofluorescence of the cells. TB hard-agar plates (1.5% (w/v)) were prepared in four different ways; either pre-inoculated with a drop of 10 μl saturated TB culture of unlabelled A or B, left empty (blanks), or addition of a drop of 10 μl tetracycline (5 mM) after 2 days (tet). Plates inoculated with A or B were placed at 33.5 °C in a humid environment for 2 days. Plates not pre-inoculated with A or B were stored at 4 °C. Subsequently all gels were covered with a layer of TB soft agar (0.26% (w/v)) containing diluted, saturated cultures of either A (labelled with YFP, A-YFP), B (labelled with CFP, B-CFP) or neither of the two (blank). The soft-agar gel contained the necessary antibiotics and inducers (see Methods). All plates were placed on 33.5 °C in a humid environment for 18 h and were then imaged. b, c, Fluorescence intensities extracted along vertical lines through the snapshots shown in a, for the CFP channel (b) and YFP channel (c). Lines were manually centred on the colonies of A and B as seen in the bright-field image. Each group indicates the hard agar/soft agar combinations. For example, A/A-YFP indicates a TB hard-agar gel pre-inoculated with unlabelled A, covered with TB soft-agar gel containing A (labelled with YFP). The presence of pre-grown B does not negatively affect the growth of A-YFP as its profile is indistinguishable from the profile of A-YFP when grown in the presence of pre-grown A (c, marker 1). The presence of tetracycline affects the growth of A-YFP negatively, as is clearly indicated by the central dip in its profile compared to the blank (c, marker 3). In the same way, the presence of pre-grown A does not negatively affect the growth of B-CFP as its profile shows higher rather than lower intensities compared to the profile of B-CFP when grown in the presence of pre-grown B (b, marker 2). The presence of tetracycline affects the growth of B-CFP negatively, as is indicated by the central dip in its profile compared to the blank (b, marker 4). Hence, we do not observe evidence that either A or B secretes growth-inhibitory compounds such as antibiotics or toxins.
Extended Data Fig. 5 Additional strains and simulation-experiment comparison.
a, b, Simulations of repeated competitive migration into a virgin nutrient patch. Top, growth and migration phenotypes of the strains are schematically illustrated. Bottom, each point is one round of competitive migration into a virgin nutrient patch. Resulting population ratios are the starting point for the next round. The populations thus can spatially segregate during rounds, and are re-mixed in between rounds. Relative population sizes are indicated as a function of a round number. a, Four populations and one common nutrient. The A–B pair (not to be confused with the strains A and B in the main text) exhibits a growth–migration trade-off and coexists (data not shown). In the presence of two additional strains, C and D, we find that the coexistence of A–B is not broken. Not all four populations coexist, as C and D become extinct. Parameters: population A (red, μ = 0.8 dbl h−1, v = 0.55 mm h−1, χ = 1.15 mm2 (h (AU))−1, D = 0.02 mm2 h−1), population B (light blue, μ = 0.6 dbl h−1, v = 0.8 mm h−1, χ = 2.25 mm2 (h (AU))−1, D = 0.02 mm2 h−1), population C (magenta, μ = 0.6 dbl h−1, v = 0.3 mm h−1, χ = 0.55 mm2 (h (AU))−1, D = 0.02 mm2 h−1) and population D (dark blue, μ = 0.4 dbl h−1, v = 0.55 mm h−1, χ = 2.0 mm2 (h (AU))−1, D = 0.02 mm2 h−1). b, The same four populations, but now A–B and C–D have their own metabolic niche, as they consume different nutrients. All four strains adopt a stable equilibrium value, illustrating how multiple coexistence mechanisms can work together to stabilize complex communities. Populations were transferred as 1/25,000 dilutions into habitats with replenished nutrients after 4 days. Nutrients are exhausted at all transfers. Habitat size: 17.5 mm. Simulation performed using the MKS model with parameters as described in the Methods. c, d, Simulations of ECOR18 and ECOR19 in gut dimensions. c, Simulations of ECOR18 (red) and ECOR19 (blue) alone (leftmost and rightmost images, respectively) and as mixed population at initial ratio of 1 (two middle images). Spatial patterns are reported after 40 h in cylindrical habitats of radius 37.5 mm and edge inoculation. Scale bar, 10 mm. Simulation performed using the MKS model with parameters as described in the Methods. d, Illustration of the cylindrical habitat. e–l, Simulations of (e–h) and experimentally measured (i–l) spatial patterns of LAB strains as monocultures (leftmost and rightmost images, respectively) and as mixed populations at initial ratio of 1 (two middle images). Experimental data as shown in Fig. 2c–f are repeated here for convenience. Spatial patterns are reported after 4 days in circular habitats of radius 17.5 mm and centre inoculation. Scale bars, 10 mm (e–h) and 7.5 mm (i–l). Simulation performed using MKS model with parameters as described in the Methods.
Extended Data Fig. 6 Different modes of motility can support coexistence.
a, Departure of fast movers depends on growth rate of slow mover. Simulated colony front position of fast movers versus time, for different growth rates of slow movers (yellow to black: μS = 0.4, 0.5, 0.6, 0.7, 0.8 dbl h−1. Parameters for slow movers: DS = 0.02 mm2 h−1, χs = 0 mm2 (h (AU))−1. Fast movers: μF = 0.5 dbl h−1, DF = 0.02 mm2 h−1, χF = 0.875 mm2 (h (AU))−1. Habitat radius: 17.5 mm. Simulations were performed using the MKS model with parameters as described in the Methods. b–d, Phase diagrams show the coexistence for chemotactic and non-chemotactic strains. Parameters are fixed for strain Y, and varied for strain X: growth rate μX (in units of μY), diffusive mobility DX (c, in units of DY; d, in mm2 h−1) and chemotactic coefficient χX (in units of χY). Blue, Y excludes X (X has low growth and motility). Red, X excludes Y (X has high growth and motility). White, stable coexistence (trade-off, with X faster grower in bottom right, X faster mover in top left). Black, unstable coexistence. b, Both X and Y are chemotactic (Y = LAB1). c, Both X and Y are non-chemotactic (Y = LAB3). d, Y is chemotactic, while X is not (Y = LAB1). Same as Fig. 4e. Simulations performed using the MKS model with parameters of indicated reference strains and DX = 0.02 mm2 h−1 in b. χx = 0 mm2 (h (AU))−1 in c and d. Habitat size in all panels: 17.5 mm. Simulation performed using MKS model with parameters as described in t he Methods. e–h, Chemotaxis is not essential for coexistence but does give an advantage. Top, snapshots of spatial density profiles at initial ratio R (fast growers (red), fast movers (blue)) of 1. Bottom, fold change versus initial ratio. Monocultures in solid lines, mixed populations as shaded regions. Chemotactic populations: both (e), only fast movers (f), only slow movers (g), neither of the two (h). e–h, Reciprocal spatial exclusion and coexistence occur, but the details differ. Specifically, when fast movers are chemotactic (e, f), they form more narrowly distributed travelling waves, spatially exclude slow movers better and increase invasion capacity (for example, at high R the fold change is higher in e than h). These results indicate a competitive advantage of chemotaxis over active random motion alone. e–h, Parameters: red fast growers (μ = 0.75 dbl h−1, v = 0.4 mm h−1), blue fast movers (μ = 0.5 dbl h−1, v = 0.8 mm h−1). e, Specific parameters: blue: χ = 3 mm2 (h (AU))−1, D = 0.02 mm2 h−1, red: χ = 0.775 mm2 (h (AU))−1, D = 0.02 mm2 h−1. f, Specific parameters: blue: χ = 3 mm2 (h (AU))−1, D = 0.02 mm2 h−1, red: χ = 0 mm2 (h (AU))−1, D = 0.0775 mm2 h−1. g, Specific parameters: blue: χ = 0 mm2 (h (AU))−1, D = 0.365 mm2 h−1, red: χ = 0.775 mm2 (h (AU))−1, D = 0.02 mm2 h−1. h, Specific parameters: blue: χ = 0 mm2 (h (AU))−1, D = 0.365 mm2 h−1, red: χ = 0 mm2 (h (AU))−1, D = 0.0775 mm2 h−1. Habitat size in all panels: 17.5 mm. Simulation was performed using the MKS model with parameters as described in the Methods. i, j, A poor competitor (i, blue: slow growth, weak random motion without chemotaxis) can become a competitive invader that can coexist with faster growers (red) when it becomes chemotactic (j). i, Parameters: blue and red are non-chemotactic and have identical expansion rates (v = 0.4 mm h−1). Red: μ = 0.75 dbl h−1, χ = 0 mm2 (h (AU))−1, D = 0.0775 mm2 h−1. Blue: μ = 0.5 dbl h−1, χ = 0 mm2 (h (AU))−1, D = 0.115 mm2 h−1. j, Parameters: red: μ = 0.75 dbl h−1, v = 0.4 mm h−1, χ = 0 mm2 (h (AU))−1, D = 0.0775 mm2 h−1. Blue: μ = 0.5 dbl h−1, v = 0.8 mm h−1, χ = 2.4 mm2 (h (AU))−1, D = 0.115 mm2 h−1. Habitat size in all panels: 17.5 mm. Simulation was performed using the MKS model with parameters as described in the Methods.
Extended Data Fig. 7 Coexistence under different inoculation and patch geometries.
a–f, Inoculation of mixed strain co-culture at the edge (a) and centre (b) of the patch, and of each strain separately at a distance of 1, 7, 13 and 19 mm relative to the centre (c–f). Inoculation regions are 1 mm wide, so they are in direct contact in c. Simulations are for a linear (one-dimensional) patch geometry. Fast-mover LAB1 (blue) and fast-grower LAB5 (red). Top, spatial pattern for initial ratio (fast grower/fast mover) of 1. Bottom, selection functions. For all cases the selection function crosses a fold change of unity (black line), indicating hierarchy inversion. The stability of coexistence, as assessed by the distance of the selection function from the unity fold change line at low and high initial ratios, is higher for separate inoculations, and increases with inoculation distance (c–f). These findings indicate that separated inoculation corresponds to having (more) pre-defined spatial niches, which require less active spatial competition, and hence yield the most robust coexistence. However, populations typically become spatially mixed, and can then still coexist by active spatial segregation and exclusion (a, b). Spatial patterns and fold changes are reported after 4 days in habitats of 20 mm linear extension. Simulation was performed using the MKS model with parameters as described in the Methods. g, h, Inoculation of mixed co-culture (LAB1 and LAB5, initial ratio: 1) in the centre (g) and around the edge (h) of a patch of circular geometry. i, j, Inoculation of mixed co-culture (LAB1 and LAB5, initial ratio: 1) in the centre (i) and edge (j) of a patch with spherical geometry. Both circular and spherical geometries also show coexistence. Note that other parameters such as nutrient concentration and initial cell density shift the selection curves (Fig. 4). Spatial patterns and fold changes are reported after 4 days in habitats of 20 mm radius. Simulation was performed using the MKS model with parameters as described in the Methods. Scale bars, 10 mm. k–m, Competition experiments with fast movers (blue, LAB2) and fast growers (red, LAB3), inoculated as either a mixed co-culture or unmixed at separate spots (close together or farther apart), at three initial ratios (fast grower/fast mover) = 10−2, 1 and 102. The measured spatial patterns and selection functions are shown. When inoculated separately, the fast movers still encircle the fast growers and reach a higher abundance and density. The selection function crosses the unity fold change line, indicating coexistence. Consistent with the simulations in c–f, the fold change of the fast movers is higher at large R0 when inoculated more distantly. These findings may be explained as follows: for separated inoculation, the fast movers have a (more) pre-defined spatial niche (location), which further limits their competition with the fast growers, allows them to reach a higher abundance, and hence increases their competitive advantage. Such a rationale is consistent with classical niche theory, which indicates that spatial niches limit between-organism competition. Taken together, both mixed and separated cultures can coexist in these spatial habitats. Scale bars, 10 mm.
Extended Data Fig. 8 Average swimming speed of cells correlates with collective migration rate.
a, Relation between average swimming speed of wild isolates in liquid rich medium (1%(w/v) TB) with their expansion rate in soft-agar motility assay (0.26% (w/v) agar). The trajectory and speed of each bacterial cell were extracted using a high-throughput three-dimensional cell-tracking method, from videos recorded at 15 Hz sampling rate35. Average swimming speeds were calculated from the full populations (that is, swimming and non-swimming). The cells were grown to their exponential phase in overnight TB culture and diluted into fresh TB medium. Open circles indicate expansion rates and mean swimming speeds (across all recorded trajectories) obtained from a single acquisition of an individual experiment. Filled circles indicate weighted-mean swimming speed across all repeated experiments (with the error-estimate weights) and mean expansion rate across two repeats (see ‘Statistics and reproducibility’ in the Methods). Cohabitant pairs bear the identical colour code. Red and black data points (open and filled circles) show the mean swimming speeds and expansion rates of flagella induction strain and its wild-type control (Extended Data Fig. 9). b, Trade-off between growth propensity (OD600 after 7.5 h, data shown in Fig. 2b and Extended Data Fig. 1f) and mean swimming speed in ECOR pairs. The observed growth–speed trade-off follows the pattern of the growth–migration trade-off shown in Figs. 2b, 4 out of 5 pairs show a difference in swimming speed and growth. Open and filled circles are defined as indicated in a. c, d, A subset of three-dimensional tracks of natural isolates A (ECOR18) (c) and B (ECOR19) (d) with a minimum duration of 20 s and a median speed larger than 10 μm s−1 are illustrated. The tracks were sampled from the full population shown in e and f. The starting point of each individual trajectory was marked by a black dot. Speed colour map shows the instantaneous speed at each point in space. e, f, Three-dimensional tracks of the total population of natural isolates A (e) and B (f). A total of 891 (A) and 1,019 (B) cell trajectories were tracked and recorded.
Extended Data Fig. 9 Cost of motility and the growth–motility trade-off.
We first estimated the synthesis and operating costs of the flagella that power motility. A single cell produces about 62,000 flagellin units (assuming 3.4 flagella per cell, each 7.3 μm long with helical pitch and radius of 2.5 μm and 0.25 μm57, respectively, composed of 11 protofilaments with flagellin repeat distances of 5.2 nm58). For flagellin synthesis, we estimate the cost as 7.7 × 108 P per cell cycle (using 25 phosphate cleavages (P) per amino acid59, 498 amino acids per flagellin). Using 47.7 kJ mol−1 for one phosphate cleavage60, the minimum power for flagella synthesis then ranges from 1 × 10−14 W (slow growth, doubling time 100 min) to 4.3 × 10−14 W (fast growth, doubling time 24 min). The operating power of one flagellar motor is estimated at 1.5 × 105 pN nm s−1 = 1.5 × 10−16 W61, resulting in 5.1 × 10−16 W for the whole cell. Thus, the cost of flagella expression is dominant. To estimate the corresponding growth penalty, we compute a cellular flagellin mass of 51 kg mol−1 × 62,000/R = 5.3 fg, given a flagellin molecular weight of 51 kDa and Avogadro’s number, R = 6 × 1023. Assuming a total cellular protein mass of 100 fg (slow growth, doubling time 100 min) to 450 fg (fast growth, 24 min)62, flagellin constitutes approximately 1.2% to 5.3% of the total protein mass of the cell. Assuming that growth costs are determined by protein synthesis, these numbers indicate that the growth penalty is significant, and provide a mechanistic rationale for a growth–motility trade-off. Other costs such as reduced membrane integrity and operating costs further increase this penalty. We investigated this issue experimentally, using new laboratory strains to allow expression control of flagella. a, Leifson stains showing flagellation for a wild-type control strain (TSS1709, 50 μM IPTG, black, middle) and a flagella induction strain (TSS1410; left, 10 μM IPTG, right, 100 μM IPTG). TSS1709 is a vector control, while TSS1410 carries an IPTG-inducible plasmid expressing the flagella master regulator FlhDC (pSJAB50) in an flhDC deletion background. In both strains, all chemoreceptors (tar, tsr, tap, trp and aer) and chemotaxis signalling genes (cheAWRBYZ) have been deleted and replaced by a plasmid expressing cheAW, tar, tsr, cheRBYZ (pTA6) to decouple them from flhDC control. b, Growth-rate dependence on IPTG induction for both strains. c, Average swimming speed of the full population (circles with solid lines) and of the motile population (crosses with dotted lines) as a function of IPTG induction. d, Colony expansion rate in soft-agar plates as a function of IPTG induction. The red square reflects the lack of expansion in the flagella induction strain in the first 24 h at 0 μM IPTG. We observed asymmetric expansion at later time points, which we attribute to mutations. In b-d, lines connect data points to guide the eye, and the grey area indicates the IPTG range in which the flagella induction strain mimics the wild-type properties. Small symbols show raw data, large symbols show averages of raw data, and error bars indicating the standard error of the mean are shown whenever n > 2. e, Soft-agar plates for various IPTG concentrations 21 h after inoculation with the wild-type control strain (bottom-right colony), the flagella induction strain (top colony) and a flagella overexpression strain that contains the inducible flhDC plasmid in addition to wild-type chromosomal flhDC (TSS1699, left colony).
Supplementary information
Supplementary Tables
This file contains Supplementary Tables 1-4.
Rights and permissions
About this article

Cite this article
Gude, S., Pinçe, E., Taute, K.M. et al. Bacterial coexistence driven by motility and spatial competition. Nature 578, 588–592 (2020). https://doi.org/10.1038/s41586-020-2033-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2033-2
This article is cited by
-
The collapse of cooperation during range expansion of Pseudomonas aeruginosa
Nature Microbiology (2024)
-
Virulence factors and mechanisms of paediatric pneumonia caused by Enterococcus faecalis
Gut Pathogens (2023)
-
Bacterial motility can govern the dynamics of antibiotic resistance evolution
Nature Communications (2023)
-
Synergistic biocontrol of Bacillus subtilis and Pseudomonas fluorescens against early blight disease in tomato
Applied Microbiology and Biotechnology (2023)
-
CRISPR evolves among the winners
Nature Ecology & Evolution (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
