
Abstract
The metabolic pathways encoded by the human gut microbiome constantly interact with host gene products through numerous bioactive molecules1. Primary bile acids (BAs) are synthesized within hepatocytes and released into the duodenum to facilitate absorption of lipids or fat-soluble vitamins2. Some BAs (approximately 5%) escape into the colon, where gut commensal bacteria convert them into various intestinal BAs2 that are important hormones that regulate host cholesterol metabolism and energy balance via several nuclear receptors and/or G-protein-coupled receptors3,4. These receptors have pivotal roles in shaping host innate immune responses1,5. However, the effect of this host–microorganism biliary network on the adaptive immune system remains poorly characterized. Here we report that both dietary and microbial factors influence the composition of the gut BA pool and modulate an important population of colonic FOXP3+ regulatory T (Treg) cells expressing the transcription factor RORγ. Genetic abolition of BA metabolic pathways in individual gut symbionts significantly decreases this Treg cell population. Restoration of the intestinal BA pool increases colonic RORγ+ Treg cell counts and ameliorates host susceptibility to inflammatory colitis via BA nuclear receptors. Thus, a pan-genomic biliary network interaction between hosts and their bacterial symbionts can control host immunological homeostasis via the resulting metabolites.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
Data availability
The microarray, RNA-seq and 16S rRNA profiling data are available in the NCBI database under accession numbers GSE68009, GSE137405 and PRJNA573477, respectively. The MS data are available in the MetaboLights database with the identifier MTBLS1276.
References
Postler, T. S. & Ghosh, S. Understanding the holobiont: how microbial metabolites affect human health and shape the immune system. Cell Metab. 26, 110–130 (2017).
Ridlon, J. M., Kang, D. J. & Hylemon, P. B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 47, 241–259 (2006).
Wahlström, A., Sayin, S. I., Marschall, H. U. & Bäckhed, F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab. 24, 41–50 (2016).
Fiorucci, S. & Distrutti, E. Bile acid-activated receptors, intestinal microbiota, and the treatment of metabolic disorders. Trends Mol. Med. 21, 702–714 (2015).
Brestoff, J. R. & Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Immunol. 14, 676–684 (2013).
Tanoue, T., Atarashi, K. & Honda, K. Development and maintenance of intestinal regulatory T cells. Nat. Rev. Immunol. 16, 295–309 (2016).
Panduro, M., Benoist, C. & Mathis, D. Tissue Tregs. Annu. Rev. Immunol. 34, 609–633 (2016).
Sefik, E. et al. Mucosal immunology. Individual intestinal symbionts induce a distinct population of RORγ+ regulatory T cells. Science 349, 993–997 (2015).
Ohnmacht, C. et al. Mucosal immunology. The microbiota regulates type 2 immunity through RORγt+ T cells. Science 349, 989–993 (2015).
Geva-Zatorsky, N. et al. Mining the human gut microbiota for immunomodulatory organisms. Cell 168, 928–943 (2017).
Xu, M. et al. c-MAF-dependent regulatory T cells mediate immunological tolerance to a gut pathobiont. Nature 554, 373–377 (2018).
Yissachar, N. et al. An intestinal organ culture system uncovers a role for the nervous system in microbe–immune crosstalk. Cell 168, 1135–1148 (2017).
Kim, K. S. et al. Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science 351, 858–863 (2016).
Sayin, S. I. et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 17, 225–235 (2013).
Yao, L. et al. A selective gut bacterial bile salt hydrolase alters host metabolism. eLife 7, e37182 (2018).
Devlin, A. S. & Fischbach, M. A. A biosynthetic pathway for a prominent class of microbiota-derived bile acids. Nat. Chem. Biol. 11, 685–690 (2015).
Wexler, A. G. & Goodman, A. L. An insider’s perspective: Bacteroides as a window into the microbiome. Nat. Microbiol. 2, 17026 (2017).
Stevens, A. M., Shoemaker, N. B. & Salyers, A. A. The region of a Bacteroides conjugal chromosomal tetracycline resistance element which is responsible for production of plasmidlike forms from unlinked chromosomal DNA might also be involved in transfer of the element. J. Bacteriol. 172, 4271–4279 (1990).
Makishima, M. et al. Vitamin D receptor as an intestinal bile acid sensor. Science 296, 1313–1316 (2002).
Makishima, M. et al. Identification of a nuclear receptor for bile acids. Science 284, 1362–1365 (1999).
DiSpirito, J. R. et al. Molecular diversification of regulatory T cells in nonlymphoid tissues. Sci. Immunol. 3, eaat5861 (2018).
Yang, B. H. et al. Foxp3+ T cells expressing RORγt represent a stable regulatory T-cell effector lineage with enhanced suppressive capacity during intestinal inflammation. Mucosal Immunol. 9, 444–457 (2016).
Britton, G. J. et al. Microbiotas from humans with inflammatory bowel disease alter the balance of gut Th17 and RORγt+ regulatory T cells and exacerbate colitis in mice. Immunity 50, 212–224 (2019).
Staley, C., Weingarden, A. R., Khoruts, A. & Sadowsky, M. J. Interaction of gut microbiota with bile acid metabolism and its influence on disease states. Appl. Microbiol. Biotechnol. 101, 47–64 (2017).
Xue, L. N. et al. Associations between vitamin D receptor polymorphisms and susceptibility to ulcerative colitis and Crohn’s disease: a meta-analysis. Inflamm. Bowel Dis. 19, 54–60 (2013).
Chen, S. et al. Cardiomyocyte-specific deletion of the vitamin D receptor gene results in cardiac hypertrophy. Circulation 124, 1838–1847 (2011).
Persson, E. et al. Simultaneous assessment of lipid classes and bile acids in human intestinal fluid by solid-phase extraction and HPLC methods. J. Lipid Res. 48, 242–251 (2007).
Sumner, L. W. et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 3, 211–221 (2007).
Song, X. et al. Growth factor FGF2 cooperates with interleukin-17 to repair intestinal epithelial damage. Immunity 43, 488–501 (2015).
Wirtz, S., Neufert, C., Weigmann, B. & Neurath, M. F. Chemically induced mouse models of intestinal inflammation. Nat. Protocols 2, 541–546 (2007).
Atarashi, K. et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331, 337–341 (2011).
Haber, A. L. et al. A single-cell survey of the small intestinal epithelium. Nature 551, 333–339 (2017).
Trapnell, C. et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protocols 7, 562–578 (2012).
Li, Y. C. et al. Targeted ablation of the vitamin D receptor: an animal model of vitamin D-dependent rickets type II with alopecia. Proc. Natl Acad. Sci. USA 94, 9831–9835 (1997).
Acknowledgements
We thank D. Gardner, Y. Li and T. Hla for providing mouse strains, and C. Fu for help with microscope. We also thank T. Sherpa and J. Ramos for help with GF mice and J. McCoy for manuscript editing. This work was supported in part by a Sponsored Research Agreement from UCB Pharma and Evelo Biosciences. S.F.O. was supported by NIH K01 DK102771.
Author information
Authors and Affiliations
Contributions
D.L.K. and X. Song designed the experiments and wrote the manuscript. X. Song, X. Sun, S.F.O., M.W., Y.Z., W.Z. and N.G.-Z. conducted or helped with the experiments. X. Song, X. Sun and Y.Z. analysed the data. R.J., D.M. and C.B. were involved in data discussions and edited the manuscript. D.L.K. supervised the study.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Pieter Dorrestein, Hiroshi Ohno and the, other, anonymous reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
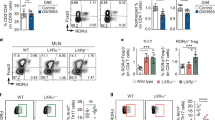
Extended Data Fig. 1 Both dietary and microbial factors control the number of colonic RORγ+ Treg cells.
a, Beginning at 3 weeks of age, three groups of mice were fed special diets for 4 weeks. SPF mice were fed either a nutrient-rich or a minimal diet, and GF mice were fed the nutrient-rich diet. Colonic Treg cells were analysed, and absolute numbers of RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population are shown. b, c, Three-week-old SPF mice were fed as in a, and Treg cells in different tissues were analysed after 4 weeks. Representative plots (b) and frequencies of RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population (c) are shown. iLN, inguinal lymph node. d–f, SPF mice were fed a nutrient-rich or a minimal diet at birth and were either maintained on that diet or switched to the opposite diet at 3 weeks of age. Colonic Treg cells were analysed after 4 weeks. Representative plots of RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population (d), and the frequencies of FOXP3+ in the CD4+TCRβ+ cell population (e) and RORγ+Helios− in the colonic FOXP3+CD4+TCRβ+ Treg cell population (f) are shown. g–i, LC–MS quantification of faecal acetate (g), propionate (h) and butyrate (i) from SPF mice fed a nutrient-rich or a minimal diet, and from GF mice fed a nutrient-rich diet. j, Three-week-old SPF mice were fed a nutrient-rich diet, a minimal diet, or a minimal diet supplemented with individual or mixed SCFAs in drinking water. Colonic Treg cells were analysed after 4 weeks. Frequencies of RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population are shown. Data are representative of two independent experiments. n represents biologically independent animals. Data are mean ± s.e.m. (a, c and e–j). *P < 0.05, **P < 0.01, ***P < 0.001, one-way ANOVA followed by the Bonferroni post hoc test (a, e–i) or two-tailed Student’s t-test (c).
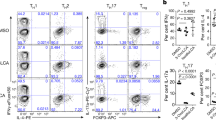
Extended Data Fig. 2 Intestinal BAs regulate the number of colonic Treg cells.
a, b, Absolute numbers of RORγ+Helios− in the colonic FOXP3+CD4+TCRβ+ Treg cell population (a) and of FOXP3+ Treg cells in the CD4+TCRβ+ population (b) in SPF mice fed a nutrient-rich diet, a minimal diet, or a minimal diet supplemented with mixtures of primary or secondary BAs in drinking water. The primary BAs were CA, CDCA and UDCA (2 mM of each). The secondary BAs were 3-oxo-CA, 3-oxo-LCA, 7-oxo-CA, 7-oxo-CDCA, 12-oxo-CA, 12-oxo-DCA, DCA and LCA (1 mM of each). c, Three-week-old SPF mice were fed a nutrient-rich diet, a minimal diet, or a minimal diet supplemented with one or more primary or secondary BAs in drinking water. Colonic TH17 cells were analysed after 4 weeks. CA, CDCA, UDCA, DCA, LCA, 3-oxo-CA, 3-oxo-LCA, 7-oxo-CA, 7-oxo-CDCA, 12-oxo-CA, 12-oxo-DCA and the indicated BA combinations were tested. Frequencies of RORγ+FOXP3− in the CD4+TCRβ+ cell population are shown. d, e, Three-week-old SPF mice were fed a nutrient-rich diet, a minimal diet, or a minimal diet supplemented with the indicated primary BAs (CA/CDCA/UDCA, 2 mM of each) or the secondary BAs (oxo-BAs/LCA/DCA, 1 mM of each) in drinking water. Treg cells and TH17 cells in the spleen, mesenteric lymph node and ileum were analysed after 4 weeks. Frequencies of RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population (d) and of RORγ+FOXP3− in the CD4+TCRβ+ cell population (e) are shown. Data are pooled from two or three independent experiments. n represents biologically independent animals. Data are mean ± s.e.m. **P < 0.01, ***P < 0.001, one-way ANOVA followed by the Bonferroni post hoc test.
Extended Data Fig. 3 Colonic microbial profiling of rich-diet mice versus minimal-diet mice.
a–d, Three-week-old SPF mice were fed a nutrient-rich or a minimal diet, and the microbial compositions in the colonic lumen were analysed after 4 weeks by 16S rRNA sequencing. Observed operational taxonomic units (OTUs) (a), Shannon index (b), principal coordinates analysis (PCoA) (c) and the relative abundance of bacteria at the phylum and family levels (d) are shown. e, Quantitative PCR analysis of 16S rDNA of Clostridium cluster IV and Clostridium cluster XIVα in colonic luminal specimens from SPF mice fed a nutrient-rich diet, a minimal diet, or a minimal diet supplemented with the indicated primary BAs (CA/CDCA/UDCA, 2 mM of each) or the secondary BAs (oxo-BAs/LCA/DCA, 1 mM of each) in drinking water. f, Four-week-old GF mice or GF mice receiving transferred faecal materials (FMTs) from minimal-diet or rich-diet SPF mice were fed a nutrient-rich diet or a minimal diet, and colonic Treg cells were analysed after 2 weeks. Frequencies of colonic RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population are shown. Data are pooled from three independent experiments in a–d. Data are representative of two independent experiments in e and f. n represents biologically independent animals. Data are mean ± s.e.m. ***P < 0.001, one-way ANOVA followed by the Bonferroni post hoc test.
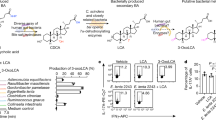
Extended Data Fig. 4 Generation of BA metabolic pathway mutants in Bacteroides.
a, Schematic diagram of pNJR6 suicide vector-mediated BA gene deletion in Bacteroides. b, Genotyping of B. thetaiotaomicron and B. fragilis BA metabolic pathway mutants by PCR. PCR primers were designed to target the flanking regions of an intact gene. PCR of an untouched gene plus its flanking regions generated a PCR product of around 1,150–1,500 bp, while deletion of an interested BA metabolic gene resulted in only an approximately 350–450-bp PCR amplicon of its two flanking regions. c, d, Bacterial load (measured as colony-forming unit (CFU) per gram of faeces) of B. thetaiotaomicron (c) and B. fragilis (d) BA metabolic pathway mutants and their wild-type control strains in monocolonized GF mice. e, f, LC–MS quantification of faecal conjugated primary BAs (e) and deconjugated primary BAs (f) in GF mice monocolonized with B. thetaiotaomicron or B. fragilis BA metabolic pathway mutants and their wild-type control strains. Data are representative of two independent experiments in b, e and f. Data are pooled from three independent experiments in c and d. n represents biologically independent animals. Data are mean ± s.e.m. (c–f).
Extended Data Fig. 5 Gut bacteria modulate colonic RORγ+ Treg cells via their BA metabolic pathways.
a, b, Each of four groups of GF mice was colonized with one of the following microorganisms: (1) a wild-type strain of B. thetaiotaomicron; (2) a BSH-mutant strain; (3) a 7α-HSDH-mutant strain; or (4) a triple-mutant (TKO) strain. Colonic Treg cells were analysed after 2 weeks. Absolute numbers of RORγ+Helios− in the colonic FOXP3+CD4+TCRβ+ Treg cell population (a) and of FOXP3+ Treg cells in the CD4+TCRβ+ population (b) are shown. c, d, Each of four groups of GF mice was colonized with one of the following microorganisms: (1) a wild-type strain of B. fragilis; (2) a BSH-KO strain; (3) a 7α-HSDH-KO strain; or (4) a double-mutant (DKO) strain. Absolute numbers of colonic Treg cells are shown as in a and b. e, f, GF mice were colonized with B. thetaiotaomicron BA metabolic pathway mutants or their wild-type control strains. Colonic Treg cells and TH17 cells were analysed after 2 weeks. Frequencies of FOXP3+ in the CD4+TCRβ+ cell population (e) or RORγ+FOXP3− in the CD4+TCRβ+ cell population (f) are shown. g, h, GF mice were colonized with B. fragilis BA metabolic pathway mutants or their wild-type control strains. Frequencies of colonic Treg cells and TH17 cells are shown as in e and f. i, j, GF mice were colonized with B. thetaiotaomicron BA metabolic pathway mutants or their wild-type control strains. Treg cells and TH17 cells in the spleen, mesenteric lymph node and ileum were analysed after 2 weeks. Frequencies of RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population (i) and RORγ+FOXP3− in the CD4+TCRβ+ cell population (j) are shown. k, l, GF mice were colonized with B. fragilis BA metabolic pathway mutants or their wild-type control strains. Frequencies of Treg cells and TH17 cells in the spleen, mesenteric lymph node and ileum are shown as in i and j. Data are pooled from two or three independent experiments. n represents biologically independent animals. Data are mean ± s.e.m. *P < 0.05, **P < 0.01, ***P < 0.001, one-way ANOVA followed by the Bonferroni post hoc test.
Extended Data Fig. 6 The effect of BAR deficiency on Treg cells or TH17 cells in gut and peripheral lymphoid organs.
a, Protein expression of VDR, FXR (also known as NR1H4) and GPBAR1 in the colonic tissue of SPF C57BL/6J mice was analysed by western blot. The red asterisks indicate the corresponding molecular weight of VDR (53 kDa), FXR (69 kDa) and GPBAR1 (33 kDa). For gel source data, see Supplementary Fig. 1. b, c, Absolute numbers of RORγ+Helios− in the colonic FOXP3+CD4+TCRβ+ Treg cell population (b) and of FOXP3+ Treg cells in the CD4+TCRβ+ population (c) from mice deficient in nuclear receptors (Nr1i2−/−Nr1i3−/−, Nr1h3−/−, Vdr−/−, Nr1h4−/− and Vdr−/−Nr1h4−/−) and their littermate controls. d, e, Frequencies of FOXP3+ in the colonic CD4+TCRβ+ cell population from mice deficient in G-protein-coupled receptors (Gpbar1−/−, Chrm2−/−, Chrm3−/− and S1pr2−/−) and their littermate controls (d) and from mice deficient in nuclear receptors (Nr1i2−/−Nr1i3−/−, Nr1h3−/−, Vdr−/−, Nr1h4−/− and Vdr−/−Nr1h4−/−) and their littermate controls (e). f, g, Frequencies of RORγ+FOXP3− in the colonic CD4+TCRβ+ cell population from mice described in d and e. h–n, Treg cells in the spleen, mesenteric lymph node and ileum from the indicated mice were analysed. Frequencies of RORγ+Helios− in the FOXP3+CD4+TCRβ+ Treg cell population from Gpbar1−/− (h), Chrm2−/− (i), Chrm3−/− (j), S1pr2−/− (k), Nr1i2−/−Nr1i3−/− (l), Nr1h3−/− (m), and Vdr−/−, Nr1h4−/− and Vdr−/−Nr1h4−/− (n) mice and their littermate controls are shown. Data are representative of two or three independent experiments in a and d–n, or are pooled from two or three independent experiments in b and c. n represents biologically independent animals. Data are mean ± s.e.m. **P < 0.01, one-way ANOVA followed by the Bonferroni post hoc test.
Extended Data Fig. 7 Dietary vitamin D3 does not alter the frequency of colonic RORγ+ Treg cells.
a, b, Beginning at 3 weeks of age, three groups of mice were fed special diets for 4 weeks. SPF mice were fed either a nutrient-rich or a minimal diet, and GF mice were fed the nutrient-rich diet. The levels of 1,25-dihydroxyvitamin D3 in serum (a) and the colon (b) of these mice were determined by ELISA. c, SPF mice were fed a nutrient-rich diet, or a rich diet deficient in vitamin D3 (VitD3) or vitamin A (VitA) at birth. Colonic Treg cells were analysed after 7 weeks. Frequencies of RORγ+Helios− in the colonic FOXP3+CD4+TCRβ+ Treg cell population are shown. d, SPF mice were fed a nutrient-rich diet at birth and were either maintained on that diet or switched to a rich diet deficient in vitamin D3 or vitamin A at 3 weeks of age. Colonic Treg cells were analysed after 4 weeks. Frequencies of RORγ+Helios− in the colonic FOXP3+CD4+TCRβ+ Treg cell population are shown. Data are representative of two independent experiments. n represents biologically independent animals. Data are mean ± s.e.m. ***P < 0.001, one-way ANOVA followed by the Bonferroni post hoc test.
Extended Data Fig. 8 Comparison of RORγ+ Treg cell signature genes of colonic Treg cells from Vdr+/+ and Vdr−/− mice.
Volcano plots comparing transcriptomes of colonic Treg cells from Vdr+/+Foxp3mRFP and Vdr−/−Foxp3mRFP mice (n = 3). Colonic RORγ+ Treg cell signature genes are highlighted in red (upregulated) or blue (downregulated). The number of genes from each signature preferentially expressed by one or the other population is shown at the bottom. Data are pooled from two independent experiments. n represents biologically independent animals. To determine the enrichment of certain gene signatures in RNA-seq datasets, a χ2 test was used. P < 0.05 was considered statistically significant.
Extended Data Fig. 9 BA supplementation does not cause gut inflammation and cannot ameliorate gut inflammation after the development of colitis.
a–c, Three-week-old SPF mice were fed a nutrient-rich diet, a minimal diet, or a minimal diet supplemented with mixtures of primary BAs (CA/CDCA/UDCA, 2 mM of each) or secondary BAs (oxo-BAs/LCA/DCA, 1 mM of each) in drinking water. Initial body weights were recorded before DSS challenge (a). Clinical scores (b) and haematoxylin and eosin histology (c) for representative colons from mice not challenged with DSS are shown. d, e, Three-week-old SPF mice fed a nutrient-rich or a minimal diet for 4 weeks were then challenged in the DSS-induced colitis model. After the development of colitis at day 5 of the model, the DSS containing drinking water was switched to regular drinking water or to drinking water supplemented with mixtures of primary or secondary BAs. The primary BAs were CA, CDCA and UDCA (2 mM of each). The secondary BAs were 3-oxo-CA, 3-oxo-LCA, 7-oxo-CA, 7-oxo-CDCA, 12-oxo-CA, 12-oxo-DCA, DCA and LCA (1 mM of each). Daily weight loss (d) of mice during the course of DSS-induced colitis and clinical scores (e) on day 10 of colitis are shown. Data are representative of two independent experiments. n represents biologically independent animals. Data are mean ± s.e.m. in a, b, d and e.
Extended Data Fig. 10 VDR signalling controls gut inflammation.
a–c, Daily weight loss (a) of Vdr+/+ and Vdr−/− mice during the course of DSS-induced colitis. Clinical scores (b) and haematoxylin and eosin histology (c) of representative colons on day 10 of colitis are shown. d, Schematic representation of the T cell-adaptive transfer model of colitis. Either Vdr+/+ or Vdr−/− naive T cells are transferred to Rag1−/− mice. e–g, Weight loss (e) of Rag1−/− mice in d during the course of T cell-adaptive transfer-induced colitis. Clinical scores (f) and haematoxylin and eosin histology (g) of representative colons on day 67 of colitis are shown. Data are representative of two independent experiments. n represents biologically independent animals. Data are mean ± s.e.m. in a, b, e and f. *P < 0.05, **P < 0.01, ***P < 0.001, two-way ANOVA followed by the Bonferroni post hoc test (a and e) or two-tailed Student’s t-test (b and f).
Supplementary information
Supplementary Information
This file contains Supplementary Table 1-2 and Supplementary Figure 1
Source data
Rights and permissions
About this article

Cite this article
Song, X., Sun, X., Oh, S.F. et al. Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis. Nature 577, 410–415 (2020). https://doi.org/10.1038/s41586-019-1865-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-019-1865-0
This article is cited by
-
The gut-liver axis in hepatobiliary diseases
Inflammation and Regeneration (2024)
-
The microbial landscape of colorectal cancer
Nature Reviews Microbiology (2024)
-
Neuropilin-1high monocytes protect against neonatal inflammation
Cellular & Molecular Immunology (2024)
-
The maternal gut microbiome in pregnancy: implications for the developing immune system
Nature Reviews Gastroenterology & Hepatology (2024)
-
Microbiota–gut–brain axis and its therapeutic applications in neurodegenerative diseases
Signal Transduction and Targeted Therapy (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
