
Abstract
Gigantopithecus blacki was a giant hominid that inhabited densely forested environments of Southeast Asia during the Pleistocene epoch1. Its evolutionary relationships to other great ape species, and the divergence of these species during the Middle and Late Miocene epoch (16–5.3 million years ago), remain unclear2,3. Hypotheses regarding the relationships between Gigantopithecus and extinct and extant hominids are wide ranging but difficult to substantiate because of its highly derived dentognathic morphology, the absence of cranial and post-cranial remains1,3,4,5,6, and the lack of independent molecular validation. We retrieved dental enamel proteome sequences from a 1.9-million-year-old G. blacki molar found in Chuifeng Cave, China7,8. The thermal age of these protein sequences is approximately five times greater than that of any previously published mammalian proteome or genome. We demonstrate that Gigantopithecus is a sister clade to orangutans (genus Pongo) with a common ancestor about 12–10 million years ago, implying that the divergence of Gigantopithecus from Pongo forms part of the Miocene radiation of great apes. In addition, we hypothesize that the expression of alpha-2-HS-glycoprotein, which has not been previously observed in enamel proteomes, had a role in the biomineralization of the thick enamel crowns that characterize the large molars in Gigantopithecus9,10. The survival of an Early Pleistocene dental enamel proteome in the subtropics further expands the scope of palaeoproteomic analysis into geographical areas and time periods previously considered incompatible with the preservation of substantial amounts of genetic information.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

Data availability
All the mass spectrometry proteomics data have been deposited in the ProteomeXchange Consortium (http://proteomecentral.proteomexchange.org) via the PRIDE partner repository with the dataset identifier PXD013838. Generated ancient protein consensus sequences for G. blacki can be found in Supplementary Data 1. Full thermal age calculations can be found in Supplementary Data 2.
References
Zhang, Y. & Harrison, T. Gigantopithecus blacki: a giant ape from the Pleistocene of Asia revisited. Am. J. Phys. Anthropol. 162 (Suppl. 63), 153–177 (2017).
Harrison, T. Apes among the tangled branches of human origins. Science 327, 532–534 (2010).
Begun, D. R. How to identify (as opposed to define) a homoplasy: examples from fossil and living great apes. J. Hum. Evol. 52, 559–572 (2007).
Kelley, J. in The Primate Fossil Record (ed. Hartwig, W. C.) 369–384 (Cambridge Univ. Press, 2002).
Miller, S. F., White, J. L. & Ciochon, R. L. Assessing mandibular shape variation within Gigantopithecus using a geometric morphometric approach. Am. J. Phys. Anthropol. 137, 201–212 (2008).
Grehan, J. R. & Schwartz, J. H. Evolution of the second orangutan: phylogeny and biogeography of hominid origins. J. Biogeogr. 36, 1823–1844 (2009).
Shao, Q. et al. ESR, U-series and paleomagnetic dating of Gigantopithecus fauna from Chuifeng Cave, Guangxi, southern China. Quat. Res. 82, 270–280 (2014).
Wang, W. New discoveries of Gigantopithecus blacki teeth from Chuifeng Cave in the Bubing Basin, Guangxi, south China. J. Hum. Evol. 57, 229–240 (2009).
Bartlett, J. D. et al. Protein–protein interactions of the developing enamel matrix. Curr. Top. Dev. Biol. 74, 57–115 (2006).
Dean, M. C. & Schrenk, F. Enamel thickness and development in a third permanent molar of Gigantopithecus blacki. J. Hum. Evol. 45, 381–388 (2003).
Von Koenigswald, G. H. R. Eine fossile Saugetierfauna mit Simia aus Sudchina. Proc. K. Ned. Akad. Wet. 38, 872–879 (1935).
Zhang, Y. et al. New 400–320 ka Gigantopithecus blacki remains from Hejiang Cave, Chongzuo City, Guangxi, South China. Quat. Int. 354, 35–45 (2014).
Zhao, L. X. & Zhang, L. Z. New fossil evidence and diet analysis of Gigantopithecus blacki and its distribution and extinction in South China. Quat. Int. 286, 69–74 (2013).
Pei, W. C. Excavation of Liucheng Gigantopithecus cave and exploration of other caves in Kwangsi. Memoir of the Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica 7, 1–54 (1965).
Bocherens, H. et al. Flexibility of diet and habitat in Pleistocene South Asian mammals: implications for the fate of the giant fossil ape Gigantopithecus. Quat. Int. 434, 148–155 (2017).
Ciochon, R. et al. Dated co-occurrence of Homo erectus and Gigantopithecus from Tham Khuyen Cave, Vietnam. Proc. Natl Acad. Sci. USA 93, 3016–3020 (1996).
Cappellini, E. et al. Early Pleistocene enamel proteome from Dmanisi resolves Stephanorhinus phylogeny. Nature 574, 103–107 (2019).
Meyer, M. et al. Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins. Nature 531, 504–507 (2016).
Wadsworth, C. & Buckley, M. Proteome degradation in fossils: investigating the longevity of protein survival in ancient bone. Rapid Commun. Mass Spectrom. 28, 605–615 (2014).
Stewart, N. A. et al. The identification of peptides by nanoLC-MS/MS from human surface tooth enamel following a simple acid etch extraction. RSC Advances 6, 61673–61679 (2016).
Castiblanco, G. A. et al. Identification of proteins from human permanent erupted enamel. Eur. J. Oral Sci. 123, 390–395 (2015).
Cristobal, A. et al. Toward an optimized workflow for middle-down proteomics. Anal. Chem. 89, 3318–3325 (2017).
Cox, J. & Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 26, 1367–1372 (2008).
Tagliabracci, V. S. et al. Secreted kinase phosphorylates extracellular proteins that regulate biomineralization. Science 336, 1150–1153 (2012).
Prado-Martinez, J. et al. Great ape genetic diversity and population history. Nature 499, 471–475 (2013).
Nater, A. et al. Morphometric, behavioral, and genomic evidence for a new orangutan species. Curr. Biol. 27, 3487–3498 (2017).
Tang, N. & Skibsted, L. H. Calcium binding to amino acids and small glycine peptides in aqueous solution: toward peptide design for better calcium bioavailability. J. Agric. Food Chem. 64, 4376–4389 (2016).
Heiss, A. et al. Structural basis of calcification inhibition by alpha 2-HS glycoprotein/fetuin-A. Formation of colloidal calciprotein particles. J. Biol. Chem. 278, 13333–13341 (2003).
Demarchi, B. et al. Protein sequences bound to mineral surfaces persist into deep time. eLife 5, e17092 (2016).
Price, P. A., Toroian, D. & Lim, J. E. Mineralization by inhibitor exclusion: the calcification of collagen with fetuin. J. Biol. Chem. 284, 17092–17101 (2009).
Kono, R. T., Zhang, Y., Jin, C., Takai, M. & Suwa, G. A 3-dimensional assessment of molar enamel thickness and distribution pattern in Gigantopithecus blacki. Quat. Int. 354, 46–51 (2014).
Mackie, M. et al. Palaeoproteomic profiling of conservation layers on a 14th century Italian wall painting. Angew. Chem. Int. Ed. 57, 7369–7374 (2018).
Jin, C. et al. Chronological sequence of the early Pleistocene Gigantopithecus faunas from cave sites in the Chongzuo, Zuojiang River area, South China. Quat. Int. 354, 4–14 (2014).
Huang, W. C. R. et al. Early Homo and associated artefacts from Asia. Nature 378, 275–278 (1995).
Pei, W. Discovery of Gigantopithecus mandibles and other material in Liucheng district of central Kwangsi in South China. Vertebrata PalAsiatica 1, 65–71 (1957).
Sun, L. et al. Magnetochronological sequence of the Early Pleistocene Gigantopithecus faunas in Chongzuo, Guangxi, southern China. Quat. Int. 354, 15–23 (2014).
Blain, H.-A., Agustí, J., Lordkipanidze, D., Rook, L. & Delfino, M. Paleoclimatic and paleoenvironmental context of the Early Pleistocene hominins from Dmanisi (Georgia, Lesser Caucasus) inferred from the herpetofaunal assemblage. Quat. Sci. Rev. 105, 136–150 (2014).
Hendy, J. et al. A guide to ancient protein studies. Nat. Ecol. Evol. 2, 791–799 (2018).
Welker, F. Elucidation of cross-species proteomic effects in human and hominin bone proteome identification through a bioinformatics experiment. BMC Evol. Biol. 18, 23 (2018).
Welker, F. Palaeoproteomics for human evolution studies. Quat. Sci. Rev. 190, 137–147 (2018).
Hanson-Smith, V. & Johnson, A. PhyloBot: a web portal for automated phylogenetics, ancestral sequence reconstruction, and exploration of mutational trajectories. PLOS Comput. Biol. 12, e1004976 (2016).
Zhang, J. et al. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteomics 11, M111.010587 (2012).
Welker, F. et al. Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte du Renne. Proc. Natl Acad. Sci. USA 113, 11162–11167 (2016).
de Manuel, M. et al. Chimpanzee genomic diversity reveals ancient admixture with bonobos. Science 354, 477–481 (2016).
Mallick, S. et al. The Simons Genome Diversity Project: 300 genomes from 142 diverse populations. Nature 538, 201–206 (2016).
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
Katoh, K. & Frith, M. C. Adding unaligned sequences into an existing alignment using MAFFT and LAST. Bioinformatics 28, 3144–3146 (2012).
Guindon, S. et al. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 59, 307–321 (2010).
Ronquist, F. et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542 (2012).
Miller, M. A., Pfeiffer, W. & Schwartz, T. in Gateway Computing Environments Workshop (GCE) 1–8 (New Orleans, 2010).
Schliep, K. P. phangorn: phylogenetic analysis in R. Bioinformatics 27, 592–593 (2011).
Le, S. Q. & Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 25, 1307–1320 (2008).
Besenbacher, S., Hvilsom, C., Marques-Bonet, T., Mailund, T. & Schierup, M. H. Direct estimation of mutations in great apes reconciles phylogenetic dating. Nat. Ecol. Evol. 3, 286–292 (2019).
Müller, T. & Vingron, M. Modeling amino acid replacement. J. Comput. Biol. 7, 761–776 (2000).
Langergraber, K. E. et al. Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution. Proc. Natl Acad. Sci. USA 109, 15716–15721 (2012).
Katoh, S. et al. New geological and palaeontological age constraint for the gorilla–human lineage split. Nature 530, 215–218 (2016).
Senut, B. et al. First hominid from the Miocene (Lukeino Formation, Kenya). Comptes Rendus Acad. Sci. IIA Earth Planet. Sci. 332, 137–144 (2001).
Brunet, M. et al. A new hominid from the Upper Miocene of Chad, central Africa. Nature 418, 145–151 (2002).
Haile-Selassie, Y. Late Miocene hominids from the Middle Awash, Ethiopia. Nature 412, 178–181 (2001).
Acknowledgements
E.C. and F.W. are supported by VILLUM FONDEN (17649) and by the European Commission through a Marie Skłodowska-Curie (MSCA) Individual Fellowship (795569). T.M.-B. is supported by BFU2017-86471-P (MINECO/FEDER, UE), NIHM grant U01 MH106874, Howard Hughes International Early Career grant, Obra Social ‘La Caixa’ and Secretaria d’Universitats i Recerca and CERCA Programme del Departament d’Economia i Coneixement de la Generalitat de Catalunya (GRC 2017 SGR 880). E.C., J.C., J.V.O., D.S. and P.G. are supported by the Marie Skłodowska-Curie European Training Network (ETN) TEMPERA, a project funded by the European Union’s EU Framework Program for Research and Innovation Horizon 2020 under grant agreement no. 722606. M.J.C. and M.M. are supported by the Danish National Research Foundation award PROTEIOS (DNRF128). Work at the Novo Nordisk Foundation Center for Protein Research is funded in part by a donation from the Novo Nordisk Foundation (NNF14CC0001). Research at Chuifeng Cave is made possible by support from the National Natural Science Foundation of China (41572023) and by a grant from the Bagui Scholar of Guangxi. M.K. was supported by a Deutsche Forschungsgemeinschaft (DFG) fellowship (KU 3467/1-1) and the Postdoctoral Junior Leader Fellowship Programme from ‘la Caixa’ Banking Foundation (LCF/BQ/PR19/11700002). M.E.A. is supported by the Independent Research Fund Denmark (7027-00147B). We thank E. Willerslev for critical reading of the manuscript, scientific support and guidance.
Author information
Authors and Affiliations
Contributions
F.W., E.C., F.D. and T.M.-B. designed the study. W.W. conduced excavation of Chuifeng Cave. W.W. and W.L. carried out faunal analysis of Chuifeng Cave. W.W., W.L. and M.E.A. provided ancient samples. F.W., J.R.-M., M.K., P.G., D.S., M.M., J.V.O. and M.d.M. performed data generation and analysed data with support from A.-M.B., M.J.C., J.C., C.L.-F., F.D. and T.M.-B. F.W., E.C. and T.M.-B. wrote the manuscript with contributions from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Peer review information Nature thanks Benedikt Kessler and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Extended data figures and tables
Extended Data Fig. 1 The current environment of Chuifeng Cave.
a, Landscape outside Chuifeng Cave. b, Elevated altitude of Chuifeng Cave (arrow points to the entrance). Photo credit: W.W.
Extended Data Fig. 2 Excavations in Chuifeng Cave.
a, Main entrance of Chuifeng Cave. b, Well-preserved deposits before excavation. c, The stratigraphic profile (1.3 m in height) of area D. d, W.W. excavating in area D. e, Excavated channel. f, g, In situ G. blacki teeth (scale bars, 3 cm). Photo credit: W.W.
Extended Data Fig. 3 Total ion current chromatograms of the analysed samples.
a, HCl extract. b, TFA extract. Note differences in maximum total ion currents on the y axes. Each extract was analysed only once.
Extended Data Fig. 4 Comparison of HCl and TFA extraction protocols.
a, Summed and normalized peptide intensities for each combination of peptide length and number of acidic residues (aspartic acid, glutamic acid, deamidated asparagine, deamidated glutamine). Circle sizes are proportional to the percentage of the total intensity, for each combination of peptide length and number of acidic residues. b, Summed and normalized intensities by peptide length. c, Summed and normalized peptide intensities across peptide hydrophobicity (GRAVY index values calculated using the R package Peptides, scale ‘KyteDoolittle’). Insets show peptide count distribution across peptide hydrophobicity. d, Extraction performance for various data categories. Values scaled to one and compared to the best-performing extraction method for each category independently. SAPs refer to those SAPs informative within Hominoidea. e, Proportional Venn diagram of unique peptide sequences identified in the two demineralization methods. All comparisons based on MaxQuant LFQ data only. N, number of peptides.
Extended Data Fig. 5 Damage characteristics of HCl and TFA extraction protocols.
a, Comparison of peptide lengths, showing an identical distribution for the TFA (n = 305) and HCl (n = 191) extractions (two-sided t-test, t394 = −0.599, P = 0.5495). b, Comparison of asparagine deamidation. c, Comparison of glutamine deamidation. b, c, Violin plots describe the distribution of bootstrap replicates (n = 1,000) of intensity-based peptide deamidation32. The number of peptides used for the calculation are shown at the bottom. For some proteins, only deamidated asparagines or glutamines were observed (for example, AMBN), while DCD is included as an example of a non-deamidated contaminant. All comparisons based on MaxQuant data only. For a–c, box plots define the range of the data (whiskers extending to 1.5× the interquartile range), outliers (beyond 1.5× the interquartile range), 25th and 75th percentiles (boxes) and medians (dots).
Extended Data Fig. 6 Unconstrained phylogenetic analysis of G. blacki.
a, Pairwise distances between groups of selected Hominoids and Pongo estimated using the concatenated protein alignments and the phangorn R package. n shows number of pairwise comparisons. b, Maximum-likelihood tree computed on a distance matrix using pml R function. Support values were obtained from 1,000 bootstrap replicates. c, Rooted phylogenetic tree obtained using MrBayes. For each bipartition, we show the posterior probability (0–1) obtained from the Bayesian approach and the support values obtained from 100 non-parametric bootstrap replicates in a PHyML maximum-likelihood (0–100) tree. PHyML and MrBayes recover the same topology. b and c are based on the same concatenated alignment of the five proteins retrieved from Gigantopithecus, and resulted in the same tree topology.
Extended Data Fig. 7 Bayesian phylogenetic tree of G. blacki including 162 modern genomes of great apes and a single hylobatid (N. leucogenys).
Tree obtained from the concatenated alignment using MrBayes. M. mulatta was used as an outgroup. The internal nodes corresponding to the Gorilla, Pan and Homo clades are collapsed for visualization purposes. The number of individuals in each of these nodes is indicated in parentheses.
Extended Data Fig. 8 Sequence conservation and structural relevance of retrieved AHSG peptides.
All AHSG-specific peptides, identified by PEAKS and MaxQuant, derive from a single sequence region bridging cystatin domains 1 and 2. The surviving sequence region is evolutionarily conserved across Catarrhini. It contains a regular repeat of acidic amino acid residues (aspartic acid, D, on positions 133, 137 and 141) that enable binding of basic calcium phosphate (residues highlighted in green), similarly to a conserved region just N-terminal (glutamic acid, E, on position 111, and aspartic acid, D, on position 113 and 115). At the bottom, a fragment ion alignment is given of MaxQuant-identified AHSG peptides. The serine is phosphorylated in all matching spectra. The majority-based amino acid consensus sequence for the alignment of Catarrhini is shown at the top for amino acid positions 100–149 (amino acid coordinates following UniProt accession P02765 [FETUA_HUMAN]).
Supplementary information
Supplementary Information
Supplementary Information containing SGDP human individual accessions, 3 supplementary tables, and 12 supplementary figures.
Supplementary Data 1
File in .fasta format containing recovered Gigantopithecus protein sequences. Sequence coverage obtained after combining the results from MaxQuant and PEAKS analysis.
Supplementary Data 2
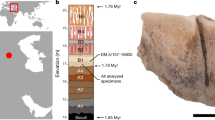
Excel file containing all numerical values used for the calculation of the thermal ages presented in Fig. 1.
Rights and permissions
About this article

Cite this article
Welker, F., Ramos-Madrigal, J., Kuhlwilm, M. et al. Enamel proteome shows that Gigantopithecus was an early diverging pongine. Nature 576, 262–265 (2019). https://doi.org/10.1038/s41586-019-1728-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-019-1728-8
This article is cited by
-
Deep-time phylogenetic inference by paleoproteomic analysis of dental enamel
Nature Protocols (2024)
-
The demise of the giant ape Gigantopithecus blacki
Nature (2024)
-
Bone collagen from subtropical Australia is preserved for more than 50,000 years
Communications Earth & Environment (2023)
-
Did some extinct South American native ungulates arise from an afrothere ancestor? A critical appraisal of Avilla and Mothé’s (2021) Sudamericungulata – Panameridiungulata hypothesis
Journal of Mammalian Evolution (2023)
-
SPIN enables high throughput species identification of archaeological bone by proteomics
Nature Communications (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
