
Abstract
Osteoarthritis—the most common form of age-related degenerative whole-joint disease1—is primarily characterized by cartilage destruction, as well as by synovial inflammation, osteophyte formation and subchondral bone remodelling2,3. However, the molecular mechanisms that underlie the pathogenesis of osteoarthritis are largely unknown. Although osteoarthritis is currently considered to be associated with metabolic disorders, direct evidence for this is lacking, and the role of cholesterol metabolism in the pathogenesis of osteoarthritis has not been fully investigated4,5,6. Various types of cholesterol hydroxylases contribute to cholesterol metabolism in extrahepatic tissues by converting cellular cholesterol to circulating oxysterols, which regulate diverse biological processes7,8. Here we show that the CH25H–CYP7B1–RORα axis of cholesterol metabolism in chondrocytes is a crucial catabolic regulator of the pathogenesis of osteoarthritis. Osteoarthritic chondrocytes had increased levels of cholesterol because of enhanced uptake, upregulation of cholesterol hydroxylases (CH25H and CYP7B1) and increased production of oxysterol metabolites. Adenoviral overexpression of CH25H or CYP7B1 in mouse joint tissues caused experimental osteoarthritis, whereas knockout or knockdown of these hydroxylases abrogated the pathogenesis of osteoarthritis. Moreover, retinoic acid-related orphan receptor alpha (RORα) was found to mediate the induction of osteoarthritis by alterations in cholesterol metabolism. These results indicate that osteoarthritis is a disease associated with metabolic disorders and suggest that targeting the CH25H–CYP7B1–RORα axis of cholesterol metabolism may provide a therapeutic avenue for treating osteoarthritis.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
Microarray data have been deposited in the Gene Expression Omnibus under accession codes GSE104793 (for IL-1β), GSE104794 (for HIF-2α), and GSE104795 (for ZIP8). Source Data for the Figures and Extended Data Figures are provided in the online version of the paper. Uncropped images of western blots and gels are available in the Supplementary Information.
References
Hunter, D. J., Schofield, D. & Callander, E. The individual and socioeconomic impact of osteoarthritis. Nat. Rev. Rheumatol. 10, 437–441 (2014).
Loeser, R. F., Goldring, S. R., Scanzello, C. R. & Goldring, M. B. Osteoarthritis: a disease of the joint as an organ. Arthritis Rheum. 64, 1697–1707 (2012).
Moon, P. M. & Beier, F. Novel insights into osteoarthritis joint pathology from studies in mice. Curr. Rheumatol. Rep. 17, 50 (2015).
Mobasheri, A. et al. The role of metabolism in the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 13, 302–311 (2017).
Niu, J., Clancy, M., Aliabadi, P., Vasan, R. & Felson, D. T. Metabolic syndrome, its components, and knee osteoarthritis: the Framingham osteoarthritis study. Arthritis Rheumatol. 69, 1194–1203 (2017).
Courties, A., Sellam, J. & Berenbaum, F. Metabolic syndrome-associated osteoarthritis. Curr. Opin. Rheumatol. 29, 214–222 (2017).
Goedeke, L. & Fernández-Hernando, C. Regulation of cholesterol homeostasis. Cell. Mol. Life Sci. 69, 915–930 (2012).
Olkkonen, V. M., Béaslas, O. & Nissilä, E. Oxysterols and their cellular effectors. Biomolecules 2, 76–103 (2012).
Kapoor, M., Martel-Pelletier, J., Lajeunesse, D., Pelletier, J. P. & Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 7, 33–42 (2011).
Yang, S. et al. Hypoxia-inducible factor-2α is a catabolic regulator of osteoarthritic cartilage destruction. Nat. Med. 16, 687–693 (2010).
Kim, J. H. et al. Regulation of the catabolic cascade in osteoarthritis by the zinc–ZIP8–MTF1 axis. Cell 156, 730–743 (2014).
Zeya, B., Arjuman, A. & Chandra, N. C. Lectin-like oxidized low-density lipoprotein (LDL) receptor (LOX-1): a chameleon receptor for oxidized LDL. Biochemistry 55, 4437–4444 (2016).
Cyster, J. G., Dang, E. V., Reboldi, A. & Yi, T. 25-Hydroxycholesterols in innate and adaptive immunity. Nat. Rev. Immunol. 14, 731–743 (2014).
Reboldi, A. et al. Inflammation. 25-Hydroxycholesterol suppresses interleukin-1-driven inflammation downstream of type I interferon. Science 345, 679–684 (2014).
Roughley, P. J. & Mort, J. S. The role of aggrecan in normal and osteoarthritic cartilage. J. Exp. Orthop. 1, 8 (2014).
Troeberg, L. & Nagase, H. Proteases involved in cartilage matrix degradation in osteoarthritis. Biochim. Biophys. Acta 1824, 133–145 (2012).
Blom, A. B. et al. Crucial role of macrophages in matrix metalloproteinase-mediated cartilage destruction during experimental osteoarthritis: involvement of matrix metalloproteinase 3. Arthritis Rheum. 56, 147–157 (2007).
Glasson, S. S. et al. Deletion of active ADAMTS5 prevents cartilage degradation in a murine model of osteoarthritis. Nature 434, 644–648 (2005).
Little, C. B. et al. Matrix metalloproteinase 13-deficient mice are resistant to osteoarthritic cartilage erosion but not chondrocyte hypertrophy or osteophyte development. Arthritis Rheum. 60, 3723–3733 (2009).
Kallen, J. A. et al. X-ray structure of the hRORα LBD at 1.63 Å: structural and functional data that cholesterol or a cholesterol derivative is the natural ligand of RORα. Structure 10, 1697–1707 (2002).
Tuong, Z. K. et al. RORα and 25-hydroxycholesterol crosstalk regulates lipid droplet homeostasis in macrophages. PLoS ONE 11, e0147179 (2016).
Guillemot-Legris, O., Mutemberezi, V. & Muccioli, G. G. Oxysterols in metabolic syndrome: from bystander molecules to bioactive lipids. Trends Mol. Med. 22, 594–614 (2016).
Hamilton, B. A. et al. Disruption of the nuclear hormone receptor RORα in staggerer mice. Nature 379, 736–739 (1996).
Dussault, I., Fawcett, D., Matthyssen, A., Bader, J. A. & Giguère, V. Orphan nuclear receptor RORα-deficient mice display the cerebellar defects of staggerer. Mech. Dev. 70, 147–153 (1998).
Kumar, N. et al. Identification of SR3335 (ML-176): a synthetic RORα selective inverse agonist. ACS Chem. Biol. 6, 218–222 (2011).
de Munter, W., van der Kraan, P. M., van den Berg, W. B. & van Lent, P. L. High systemic levels of low-density lipoprotein cholesterol: fuel to the flames in inflammatory osteoarthritis? Rheumatology 55, 16–24 (2016).
Farnaghi, S., Crawford, R., Xiao, Y. & Prasadam, I. Cholesterol metabolism in pathogenesis of osteoarthritis disease. Int. J. Rheum. Dis. 20, 131–140 (2017).
Garcia-Gil, M. et al. Serum lipid levels and risk of hand osteoarthritis: the Chingford Prospective Cohort Study. Sci. Rep. 7, 3147 (2017).
Ali, S. A. et al. Regulation of cholesterol homeostasis by Hedgehog signaling in osteoarthritic cartilage. Arthritis Rheumatol. 68, 127–137 (2016).
Ryu, J. H. et al. Hypoxia-inducible factor-2α is an essential catabolic regulator of inflammatory rheumatoid arthritis. PLoS Biol. 12, e1001881 (2014).
Glasson, S. S., Blanchet, T. J. & Morris, E. A. The surgical destabilization of the medial meniscus (DMM) model of osteoarthritis in the 129/SvEv mouse. Osteoarthritis Cartilage 15, 1061–1069 (2007).
Son, Y. O. et al. Estrogen-related receptor γ causes osteoarthritis by upregulating extracellular matrix-degrading enzymes. Nat. Commun. 8, 2133 (2017).
Glasson, S. S., Chambers, M. G., Van Den Berg, W. B. & Little, C. B. The OARSI histopathology initiative – recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthritis Cartilage 18 (Suppl. 3), S17–S23 (2010).
Krenn, V. et al. Synovitis score: discrimination between chronic low-grade and high-grade synovitis. Histopathology 49, 358–364 (2006).
Das Neves Borges, P., Vincent, T. L. & Marenzana, M. Automated assessment of bone changes in cross-sectional micro-CT studies of murine experimental osteoarthritis. PLoS ONE 12, e0174294 (2017).
Malfait, A. M., Little, C. B. & McDougall, J. J. A commentary on modelling osteoarthritis pain in small animals. Osteoarthritis Cartilage 21, 1316–1326 (2013).
Gosset, M., Berenbaum, F., Thirion, S. & Jacques, C. Primary culture and phenotyping of murine chondrocytes. Nat. Protoc. 3, 1253–1260 (2008).
Rutkowska, A. et al. The EBI2 signalling pathway plays a role in cellular crosstalk between astrocytes and macrophages. Sci. Rep. 6, 25520 (2016).
Acknowledgements
This work was supported by grants from the National Research Foundation of Korea (2016R1A3B1906090 and 2016R1A5A1007318 to J.-S.C., 2012R1A5A2A39671455 and 2018R1A2B2006033 to J.-H.R., and 2017R1A6A3A11034719 to W.-S.C.), the Korea Healthcare Technology R&D project of the Korea Health Industry Development Institute (HI16C0287 to J-.S.C., J.-H.R. and H.A.K., and H114C3484 to J.-S.C. and J.-H.R.), and the GIST Research Institute (GRI) to J.-S.C.
Reviewer information
Nature thanks Benjamin Alman and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
W.-S.C. and G.L. designed, performed and analysed the experiments. W.-H.S., J.Y., J.-S.K., H.-E.K., S.K.K. and Y.-O.S. assisted with the experimental analyses. H.N. and I.J. performed the bioinformatic analysis. Z.-Y.P. and J.K. carried out the liquid chromatography–tandem mass spectrometry analysis. I.Y.P. and J.-I.H. performed the pain assay. H.A.K., J.-T.K. and C.-H.C. evaluated the human samples and critically revised the manuscript. W.-S.C, G.L., J.-H.R. and J.-S.C wrote the manuscript. J.-H.R. and J.-S.C. conceived, planned and oversaw the study.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 OA-associated catabolic signalling increases cholesterol uptake in chondrocytes.
a, Mice fed a regular diet (RD) or HCD were subjected to sham operation or DMM surgery. Left to right, safranin-O staining, scoring of cartilage destruction (OARSI grade), osteophyte maturity, subchondral bone plate (SBP) thickness (n = 10) and serum cholesterol levels (n = 5). b, c, Cholesterol levels in chondrocytes treated with IL-1β or TNFα for 36 h or infected for 36 h with Ad-C (800 MOI), Ad-HIF-2α or Ad-ZIP8 (b). Levels of Hif2a (also known as Epas1) and Zip8 (also known as Slc39a8) mRNA in adenovirus-infected chondrocytes (c). n = 8 each. d, Cholesterol levels in chondrocytes treated with IL-1β (5 ng ml−1) and the cholesterol synthesis inhibitors triparanol (TP) or atorvastatin (AV) for 36 h (n = 9). e, mRNA levels of proteins involved in cholesterol synthesis and efflux in chondrocytes treated with IL-1β or TNFα (n = 5) or in damaged regions of human OA (versus undamaged regions, n = 4) and DMM-operated cartilage (versus sham operation, n = 5). f, Characteristics of individuals with OA from whom cartilage samples were taken. g, Representative images (n = 5) of LOX1 expression, NBD-cholesterol uptake, and DIO-oxLDL uptake in chondrocytes treated with control or Lox1 siRNA with or without IL-1β or TNFα, or infected with 800 MOI of Ad-C, Ad-HIF-2α or Ad-ZIP8. Means ± 95% CI with Mann–Whitney U test for OARSI grade and osteophyte maturity; means ± s.e.m. with two-tailed t-test for SBP thickness and cholesterol levels (a). Means ± s.e.m. with one-way ANOVA and Bonferroni test (b, c, d) and means ± s.e.m. with two-tailed t-test (e). n indicates the number of biologically independent samples or mice per group. Exact P values (for P < 0.001) can be found in the accompanying Source Data. Scale bar, 25 μm.
Extended Data Fig. 2 Upregulation of CH25H and CYP7B1 in OA chondrocytes.
a–c, Microarray analysis of cholesterol metabolism-related genes in chondrocytes treated with IL-1β, Ad-C (control), Ad-HIF-2α or Ad-ZIP8. a, Fold changes in mRNA levels versus those in vehicle or Ad-C-infected cells (n = 4). b, Heat maps (left, IL-1β; centre, Ad-HIF-2α; right, Ad-ZIP8) of genes involved in cholesterol metabolism and metabolic pathways of bile acid synthesis in the liver. c, PPI network based on CH25H and CYP7B1. Nodes and links represent protein-encoding genes and their interactions, respectively. Genes shown in red and blue indicate those annotated as upregulated and downregulated, respectively, in the PPI network. d, e, mRNA levels (d) and representative RT–PCR gel images (e) in chondrocytes treated with IL-1β or TNFα, or infected with Ad-C (800 MOI; left), Ad-HIF-2α (middle) or Ad-ZIP8 (right; n = 5). f, Safranin-O staining and immunostaining of the indicated proteins in mice that underwent intra-articular injection of Ad-HIF-2α or Ad-ZIP8 (n = 7). Means ± s.e.m. with one-way ANOVA and Bonferroni test. n indicates the number of biologically independent samples or mice per group. Exact P values (for P < 0.001) can be found in the accompanying Source Data. For gel source data, see Supplementary Fig. 1. Scale bars, 25 μm.
Extended Data Fig. 3 Production of oxysterols in chondrocytes stimulated with OA-associated catabolic signalling.
a, b, Chondrocytes were treated with IL-1β or TNFα, or infected with 800 MOI of Ad-C, Ad-CH25H, Ad-CYP7B1, Ad-HIF-2α or Ad-ZIP8. The amounts of 25-HC (a) and 7α,25-HC (b) were measured using HPLC–MS/MS (n = 4 biologically independent samples). Representative peak areas of endogenous oxysterols and corresponding internal standards are shown.
Extended Data Fig. 4 CH25H regulates OA pathogenesis in mice.
a, b, Representative images of CH25H immunostaining (a) and safranin-O staining (b) in mice that underwent intra-articular injection of Ad-C or Ad-CH25H once weekly for 3 weeks (n = 12 mice per group). c, Representative immunostaining of CH25H in cartilage sections of 20-week-old wild-type and Ch25h−/− (KO) mice (n = 12 per group). d, Staining of whole skeleton, hindlimbs and forelimbs, and the length of the indicated bones, in wild-type (n = 6) and Ch25h−/− (n = 12) mice. e, Staining of growth plates in metatarsal bones, along with the lengths of the indicated zones, from Ch25h−/− mice and wild-type littermates (n = 20 mice per group). f, Representative immunostaining images of CH25H, DIPEN and NITEGE in Ch25h−/− and wild-type littermates subjected to sham or DMM surgery (n = 8 mice per group). g, OA parameters quantified 8 and 12 weeks after Ch25h−/− and wild-type littermates were subjected to sham operation (n = 8 mice) or DMM surgery (n = 12 mice). h, Wild-type and Ch25h−/− mice fed regular diet or HCD were subjected to sham or DMM operations, with OA parameters quantified at 4 (n = 8 mice for sham and 6 mice for DMM) and 6 weeks (n = 9 mice for sham and 9 mice for DMM). i, von Frey (n = 10 mice per group) and hot-plate (n = 15 mice per group) pain assays in DMM-operated Ch25h−/− and wild-type mice at the indicated times after operation. Means ± s.e.m. with two-tailed t-test (d, e). Means ± 95% CI with Mann–Whitney U test for OARSI grade, osteophyte maturity and von Frey assay; means ± s.e.m. with two-tailed t-test for SBP thickness and hot-plate assay (g−i). Exact P values (for P < 0.001) can be found in the accompanying Source Data. Scale bars, 25 μm.
Extended Data Fig. 5 CYP7B1 regulates OA pathogenesis in mice.
a, b, Wild-type mice underwent intra-articular injection of Ad-C or Ad-CYP7B1 and were killed three weeks later. Representative images of CYP7B1 immunostaining (a) and safranin-O staining (b). n = 9 mice per group. c, d, Sham- or DMM-operated mice underwent intra-articular injection of Ad-shControl or Ad-shCyp7b1, and were killed 8 or 12 weeks later. Representative immunostaining images of CYP7B1 in cartilage sections (c) and quantification of OA parameters (d). n = 10 mice for sham and 13 mice for DMM. e, Sham- or DMM-operated mice fed regular diet or HCD underwent intra-articular injection of Ad-shControl or Ad-shCyp7b1. The mice were killed four or six weeks after the operation, and OA parameters were quantified (n = 10 mice for sham and 11 mice for DMM). f, g, Pain assays. Sham- or DMM-operated mice underwent intra-articular injection of Ad-shControl or Ad-shCyp7b1 (n = 9 mice per group) and were subjected to von Frey (f) and hot-plate (g) assays. Means ± 95% CI with Mann–Whitney U test for OARSI grade, osteophyte maturity and von Frey assay; means ± s.e.m. with two-tailed t-test for SBP thickness and hot-plate assay (d–g). Exact P values (for P < 0.001) can be found in the accompanying Source Data. Scale bars, 25 μm.
Extended Data Fig. 6 Cholesterol metabolism regulates the expression of matrix-degrading enzymes in chondrocytes.
a, b, Mouse articular chondrocytes in primary culture were treated for 36 h with MβCD-cholesterol, 25-HC or 7α,25-HC. Alternatively, chondrocytes were infected with 800 MOI of Ad-C, Ad-CH25H or Ad-CYP7B1. a, Relative mRNA levels (fold change against control) of the indicated molecules were quantified by qRT–PCR analysis (n = 6 biologically independent samples). b, Western blots of secreted MMP3 and MMP13 in the culture supernatants of chondrocytes (n = 5 biologically independent samples). c, Representative immunostaining of MMP3 and MMP13 in cartilage from sham- or DMM-operated wild-type and Ch25h−/− mice (left) and DMM-operated wild-type mice that received intra-articular injection of Ad-shControl or Ad-shCyp7b1 (right; n = 8 mice per group). d, e, Detection of aggrecan neoepitopes in the cartilage of mice that received intra-articular injection of Ad-C, Ad-CH25H or Ad-CYP7B1 (d; n = 8 mice per group) and sham- or DMM-operated mice fed regular diet or HCD (e, n = 10 mice per group). Means ± s.e.m. with two-tailed t-test. Exact P values (for P < 0.001) can be found in the accompanying Source Data. Scale bars, 25 μm.
Extended Data Fig. 7 RORα is a downstream mediator of cholesterol metabolism in chondrocytes.
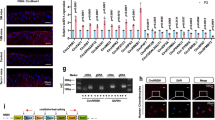
a, mRNA levels of putative oxysterol-binding nuclear receptors in chondrocytes treated with IL-1β, TNFα, cholesterol, 25-HC or 7α,25-HC for 36 h (n = 5), in damaged regions of cartilage from humans with OA (versus undamaged regions; n = 4), and in cartilage from DMM-operated mice (versus sham-operated mice; n = 5). b, RT–PCR and qRT–PCR (n = 5) analyses of ROR isoforms in chondrocytes treated with IL-1β or TNFα for 36 h. c, d, Representative immunostaining images of RORα and RORγ in damaged and undamaged regions of cartilage from the same patient (c; n = 10) and sham- or DMM-operated mice (d; n = 7). e, Left, saturation fluorescent polarization assays assessing the binding of 22-NBD cholesterol (200 nM) to increasing amounts of full-length recombinant RORα protein. Right, competition fluorescent polarization assays to determine the competition of the indicated concentrations of cholesterol, 25-HC, and 7α,25-HC with 22-NBD cholesterol (200 nM) for binding to full-length recombinant RORα protein (5 nM; n = 4). f, qRT–PCR analysis of the indicated molecules in chondrocytes infected with Ad-RORα (n = 5). g, ChIP assays for RORα binding to the promoter of each target gene in chondrocytes infected with 800 MOI of Ad-C (C) or Ad-RORα (R) (n = 5). Means ± s.e.m. with two-tailed t-test (a) and with one-way ANOVA and Bonferroni test (b, f). n indicates the number of biologically independent samples, mice per group, or human specimens. Exact P values (for P < 0.001) can be found in the accompanying Source Data. For gel source data, see Supplementary Fig. 1. Scale bars, 25 μm.
Extended Data Fig. 8 RORα overexpression enhances experimental OA in mice.
a, Representative immunostaining images of RORα and the aggrecan neoepitopes DIPEN and NITEGE in cartilage of mice that received intra-articular injection of Ad-C or Ad-RORα (n = 5 mice per group). b, Representative immunostaining images of RORα in knee joint sections of transgenic and wild-type mice (n = 4 mice each). c, Transgenic and wild-type mice were subjected to sham operation or DMM surgery. Representative safranin-O staining images of joint sections (left) and quantification of OARSI grade, osteophyte maturity and SBP thickness (right; n = 6 mice per group), 6 weeks after sham operation or DMM surgery. Means ± 95% CI with Mann–Whitney U test for OARSI grade and osteophyte maturity; means ± s.e.m. with two-tailed t-test for SBP thickness. Exact P values (for P < 0.001) can be found in the accompanying Source Data. Scale bars, 25 μm.
Extended Data Fig. 9 Regulation of OA pathogenesis by RORα.
a–c, Whole-skeleton staining (a), staining of hindlimbs and forelimbs and lengths of the indicated bones on postnatal day 1 in RoraSg/+ and wild-type mice (b; n = 6 each), and representative microCT images of the tibia and the length of the indicated region in 4-month-old RoraSg/+ and wild-type mice (c; n = 8 each). d, Representative immunostaining images of RORα in cartilage sections from DMM-operated RoraSg/+ and wild-type mice (n = 5 each). e, Quantification of OA manifestations 8 (n = 6 mice for sham and 7 mice for DMM) or 12 (n = 5 each) weeks after sham or DMM surgery in RoraSg/+ and wild-type mice. f, RoraSg/+ and wild-type mice fed regular diet or HCD were subjected to sham or DMM surgery, and OA parameters were quantified after 4 (n = 6 each) or 6 (n = 5 each) weeks. g, h, Safranin-O staining and scoring of OARSI grade in mice that underwent intra-articular injection of Ad-RORα with vehicle or SR3335, or sham- or DMM-operated mice that underwent intra-articular injection of vehicle or SR3335 (n = 5 mice each). i, mRNA levels of indicated molecules in chondrocytes treated with cholesterol for 36 h or infected with Ad-C, Ad-CH25H or Ad-CYP7B1 for 36 h in the presence of the indicated concentrations of SR3335 (n = 6 mice per group). Means ± s.e.m. with two-tailed t-test (b, c). Means ± 95% CI with Mann–Whitney U test for OARSI grade and osteophyte maturity; means ± s.e.m. with two-tailed t-test for SBP thickness (e–h). Means ± s.e.m. with one-way ANOVA and Bonferroni test (i). Exact P values (for P < 0.001) can be found in the accompanying Source Data. Scale bars, 25 μm.
Extended Data Fig. 10 Regulation of cholesterol metabolism by OA-associated catabolic signalling in FLSs and Raw264.7 cells.
a, Total cholesterol levels in FLSs treated with IL-1β (1 ng ml−1, 36 h) or TNFα (50 ng ml−1, 36 h) or infected with 800 MOI of Ad-C, Ad-HIF-2α or Ad-ZIP8 for 36 h (n = 6). b, mRNA levels in FLSs treated with IL-1β or TNFα or infected with Ad-HIF-2α or Ad-ZIP8 (n = 6). c, mRNA levels of the indicated molecules in FLSs treated with cholesterol or infected with Ad-CH25H, Ad-CYP7B1 or Ad-RORα (n = 6). d, Total cholesterol levels in Raw264.7 cells treated as in a (n = 7). e, mRNA levels of the indicated molecules in Raw264.7 cells treated with IL-1β or TNFα or infected with Ad-C, Ad-HIF-2α or Ad-ZIP8 (n = 7). f, RT–PCR analysis of the indicated molecules in Raw264.7 cells treated with IL-1β or TNFα or infected with Ad-HIF-2α or Ad-ZIP8 (n = 7). g, mRNA levels in Raw264.7 cells treated with cholesterol or infected with Ad-CH25H (n = 6). h, Total cholesterol levels and mRNA levels (n = 6) of the indicated molecules in human normal chondrocytes treated with IL-1β (2.5 ng ml−1, 36 h) or TNFα (50 ng ml−1, 36 h). i, Representative immunostaining images of CH25H, CYP7B1 and RORα in synovial tissue from humans with OA or rheumatoid arthritis (n = 4). Means ± s.e.m. with two-tailed t-test (a, d, h) or with one-way ANOVA and Bonferroni test (b, c, e, g). n indicates the number of biologically independent samples or human specimens. Exact P values (for P < 0.001) can be found in the accompanying Source Data. For gel source data, see Supplementary Fig. 1. Scale bars, 25 μm.
Supplementary information
Supplementary Information
This file contains Supplementary Figure 1: Uncropped scans with size marker indications.
Source data
Rights and permissions
About this article

Cite this article
Choi, WS., Lee, G., Song, WH. et al. The CH25H–CYP7B1–RORα axis of cholesterol metabolism regulates osteoarthritis. Nature 566, 254–258 (2019). https://doi.org/10.1038/s41586-019-0920-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-019-0920-1
This article is cited by
-
Identification and validation of biomarkers related to lipid metabolism in osteoarthritis based on machine learning algorithms
Lipids in Health and Disease (2024)
-
Genetic evidence of the causal relationship between chronic liver diseases and musculoskeletal disorders
Journal of Translational Medicine (2024)
-
Metabolic regulation of skeletal cell fate and function
Nature Reviews Endocrinology (2024)
-
5-aminosalicylic acid suppresses osteoarthritis through the OSCAR-PPARγ axis
Nature Communications (2024)
-
A Novel Hybrid of Telmisartan and Borneol Ameliorates Neuroinflammation and White Matter Injury in Ischemic Stroke Through ATF3/CH25H Axis
Translational Stroke Research (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
