
Abstract
The sensory cells that are responsible for hearing include the cochlear inner hair cells (IHCs) and outer hair cells (OHCs), with the OHCs being necessary for sound sensitivity and tuning1. Both cell types are thought to arise from common progenitors; however, our understanding of the factors that control the fate of IHCs and OHCs remains limited. Here we identify Ikzf2 (which encodes Helios) as an essential transcription factor in mice that is required for OHC functional maturation and hearing. Helios is expressed in postnatal mouse OHCs, and in the cello mouse model a point mutation in Ikzf2 causes early-onset sensorineural hearing loss. Ikzf2cello/cello OHCs have greatly reduced prestin-dependent electromotile activity, a hallmark of OHC functional maturation, and show reduced levels of crucial OHC-expressed genes such as Slc26a5 (which encodes prestin) and Ocm. Moreover, we show that ectopic expression of Ikzf2 in IHCs: induces the expression of OHC-specific genes; reduces the expression of canonical IHC genes; and confers electromotility to IHCs, demonstrating that Ikzf2 can partially shift the IHC transcriptome towards an OHC-like identity.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
The RiboTag OHC RNA-seq, P8 cello cochlea RNA-seq, and P8 Anc80-Ikzf2 and Anc80-eGFP injected cochlea scRNA-seq data have been submitted to the Gene Expression Omnibus (GEO) database under accessions GSE116703, GSE116702 and GSE120462, and are also available for viewing through the gEAR portal (https://umgear.org/).
References
Dallos, P. et al. Prestin-based outer hair cell motility is necessary for mammalian cochlear amplification. Neuron 58, 333–339 (2016).
Ehret, G. Development of absolute auditory thresholds in the house mouse (Mus musculus). J. Am. Audiol. Soc. 1, 179–184 (1976).
Bielefeld, E. C., Tanaka, C., Chen, G. D. & Henderson, D. Age-related hearing loss: is it a preventable condition? Hear. Res. 264, 98–107 (2010).
World Health Organization. Deafness and Hearing Loss; available at: http://www.who.int/mediacentre/factsheets/fs300/en/ (2018).
Mittal, R. et al. Recent advancements in the regeneration of auditory hair cells and hearing restoration. Front. Mol. Neurosci. 10, 236 (2017).
Fang, J. et al. Outer hair cell-specific prestin-CreERT2 knockin mouse lines. Genesis 50, 124–131 (2012).
Sanz, E. et al. Cell-type-specific isolation of ribosome-associated mRNA from complex tissues. Proc. Natl Acad. Sci. USA 106, 13939–13944 (2009).
Liu, H. et al. Characterization of transcriptomes of cochlear inner and outer hair cells. J. Neurosci. 34, 11085–11095 (2014).
Elkon, R. et al. RFX transcription factors are essential for hearing in mice. Nat. Commun. 6, 8549 (2015).
Hertzano, R. et al. Cell type-specific transcriptome analysis reveals a major role for Zeb1 and miR-200b in mouse inner ear morphogenesis. PLoS Genet. 7, e1002309 (2011).
Janky, R. et al. iRegulon: from a gene list to a gene regulatory network using large motif and track collections. PLOS Comput. Biol. 10, e1003731 (2014).
Potter, P. K. et al. Novel gene function revealed by mouse mutagenesis screens for models of age-related disease. Nat. Commun. 7, 12444 (2016).
Ashmore, J. F. A fast motile response in guinea-pig outer hair cells: the cellular basis of the cochlear amplifier. J. Physiol. (Lond.) 388, 323–347 (1987).
Brownell, W. E., Bader, C. R., Bertrand, D. & de Ribaupierre, Y. Evoked mechanical responses of isolated cochlear outer hair cells. Science 227, 194–196 (1985).
Marcotti, W. & Kros, C. J. Developmental expression of the potassium current I K,n contributes to maturation of mouse outer hair cells. J. Physiol. (Lond.) 520, 653–660 (1999).
Li, Y. et al. Transcription factors expressed in mouse cochlear inner and outer hair cells. PLoS One 11, e0151291 (2016).
Zinn, E. et al. In silico reconstruction of the viral evolutionary lineage yields a potent gene therapy vector. Cell Reports 12, 1056–1068 (2015).
Landegger, L. D. et al. A synthetic AAV vector enables safe and efficient gene transfer to the mammalian inner ear. Nat. Biotechnol. 35, 280–284 (2017).
Roux, I. et al. Otoferlin, defective in a human deafness form, is essential for exocytosis at the auditory ribbon synapse. Cell 127, 277–289 (2006).
Simmons, D. D., Tong, B., Schrader, A. D. & Hornak, A. J. Oncomodulin identifies different hair cell types in the mammalian inner ear. J. Comp. Neurol. 518, 3785–3802 (2010).
Santos-Sacchi, J. Reversible inhibition of voltage-dependent outer hair cell motility and capacitance. J. Neurosci. 11, 3096–3110 (1991).
Zheng, J. et al. Prestin is the motor protein of cochlear outer hair cells. Nature 405, 149–155 (2000).
Caberlotto, E. et al. Usher type 1G protein sans is a critical component of the tip-link complex, a structure controlling actin polymerization in stereocilia. Proc. Natl Acad. Sci. USA 108, 5825–5830 (2011).
Johnson, K. R., Zheng, Q. Y. & Noben-Trauth, K. Strain background effects and genetic modifiers of hearing in mice. Brain Res. 1091, 79–88 (2006).
Hoelter, S. M. et al. “Sighted C3H” mice—a tool for analysing the influence of vision on mouse behaviour? Front. Biosci. 13, 5810–5823 (2008).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013).
Anders, S., Pyl, P. T. & Huber, W. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Anders, S. & Huber, W. Differential expression analysis for sequence count data. Genome Biol. 11, R106 (2010).
Ulitsky, I. et al. Expander: from expression microarrays to networks and functions. Nat. Protocols 5, 303–322 (2010).
Sharan, R., Maron-Katz, A. & Shamir, R. CLICK and EXPANDER: a system for clustering and visualizing gene expression data. Bioinformatics 19, 1787–1799 (2003).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Adzhubei, I. A. et al. A method and server for predicting damaging missense mutations. Nat. Methods 7, 248–249 (2010).
Choi, Y., Sims, G. E., Murphy, S., Miller, J. R. & Chan, A. P. Predicting the functional effect of amino acid substitutions and indels. PLoS One 7, e46688 (2012).
Kumar, P., Henikoff, S. & Ng, P. C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protocols 4, 1073–1081 (2009).
Källberg, M. et al. Template-based protein structure modeling using the RaptorX web server. Nat. Protocols 7, 1511–1522 (2012).
Hardisty-Hughes, R. E., Parker, A. & Brown, S. D. M. A hearing and vestibular phenotyping pipeline to identify mouse mutants with hearing impairment. Nat. Protocols 5, 177–190 (2010).
Mianné, J. et al. Correction of the auditory phenotype in C57BL/6N mice via CRISPR/Cas9-mediated homology directed repair. Genome Med. 8, 16 (2016).
Codner, G. F. et al. Application of long single-stranded DNA donors in genome editing: generation and validation of mouse mutants. BMC Biol. 16, 70 (2018).
Corns, L. F., Johnson, S. L., Kros, C. J. & Marcotti, W. Calcium entry into stereocilia drives adaptation of the mechanoelectrical transducer current of mammalian cochlear hair cells. Proc. Natl Acad. Sci. USA 111, 14918–14923 (2014).
Santos-Sacchi, J. Determination of cell capacitance using the exact empirical solution of partial δY/δC m and its phase angle. Biophys. J. 87, 714–727 (2004).
Santos-Sacchi, J. & Navarrete, E. Voltage-dependent changes in specific membrane capacitance caused by prestin, the outer hair cell lateral membrane motor. Pflugers Arch. 444, 99–106 (2002).
Isgrig, K. et al. Gene therapy restores balance and auditory functions in a mouse model of usher syndrome. Mol. Ther. 25, 780–791 (2017).
Finak, G. et al. MAST: a flexible statistical framework for assessing transcriptional changes and characterizing heterogeneity in single-cell RNA sequencing data. Genome Biol. 16, 278 (2015).
Satija, R., Farrell, J. A., Gennert, D., Schier, A. F. & Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 33, 495–502 (2015).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer, New York, 2009).
Gaujoux, R. & Seoighe, C. A flexible R package for nonnegative matrix factorization. BMC Bioinformatics 11, 367 (2010).
Geng, R. et al. Comprehensive expression of Wnt signaling pathway genes during development and maturation of the mouse cochlea. PLoS One 11, e0148339 (2016).
Acknowledgements
We thank L. Vizor, J. Sanderson and W. Chien for technical help and Z. Ahmed for comments on the manuscript. This work was supported by Action on Hearing Loss (G65 to M.R.B., R.H., W.M. and S.D.M.B.), Medical Research Council (MC_U142684175 to S.D.M.B.), Wellcome Trust (102892 to W.M.), NIDCD/NIH R01DC013817 and R01DC03544 (R.H.), DOD CDMRP MR130240 (R.H.), NIDCD/NIH T32DC00046 and F31DC016218 (M.S.M.), the Intramural Program at NIDCD DC000059 (M.W.K.), and NIDCD/NIH R01DC014658 (G.I.F.). S.L.J. is a Royal Society University Research Fellow. R.E. is a Faculty Fellow of the Edmond J. Safra Center for Bioinformatics at Tel Aviv University.
Reviewer information
Nature thanks M. Montcouquiol, B. Walters and the anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
L.C., M.S.M., M.C.K., Y.O., B.M., M.M., G.C., C.T.E., G.I.F., M.W.K., W.M., S.D.M.B., M.R.B. and R.H. designed and interpreted the experiments. L.C., M.S.M., M.C.K., S.L.J., Y.O., B.M., M.M., A.P., J.P., R.H., E.C.D., G.T., A.K.D., G.I.F. and W.M. performed the experiments. Y.S. and R.E. analysed the gene expression data. S.W. aided in the management of the cello colony. M.R.B. and R.H. conceived and coordinated the study. L.C., M.S.M., M.C.K., W.M., S.D.M.B., R.E., M.R.B. and R.H. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 RiboTag immunoprecipitation enriches for known OHC-expressed transcripts.
a, Representative prestincreERT2/+;ROSA26CAG-tdTomato cochlear whole-mount. The prestincreERT2-driven tdTomato expression is OHC-specific at P21 (n = 1). Scale bar, 20 µm. b, Schematic of the RiboTag immunoprecipitation protocol. Red OHCs represent Cre/HA-tagged ribosome expression. c, RiboTag RNA-seq log2 enrichment and depletion of transcripts for known inner ear cell type markers (EF = log2(IP/input)). d, Genes at least two-fold enriched in IHCs (n = 565 genes) or OHCs (n = 253 genes) in the published dataset8 are significantly depleted or enriched, respectively, by the RiboTag OHC immunoprecipitation at all time points examined (two-sided Wilcoxon’s test). Black line represents median EF, box demarcates first and third quartiles, whiskers demarcate first and third quartiles ± 1.5 × IQR values, dots represent single outliers. e, Clustering of genes differentially expressed across OHC postnatal development. Error bars denote s.d. Before clustering, expression levels were standardized to mean = 0 and s.d. = 1. f, Enriched Gene Ontology (GO) functional categories identified for the gene clusters in e (cluster 1 n = 160 genes, cluster 2 n = 63 genes). No significantly enriched GO categories were found for cluster 3 (n = 79 genes). Enrichment and statistical analyses were performed using the EXPANDER implemented tool TANGO.
Extended Data Fig. 2 Auditory phenotyping, SNP mapping and whole-genome sequencing of mouse pedigree MPC173, subsequently named cello.
a, Specific expression of Helios can be seen in the nuclei of wild-type P8 OHCs (white arrow), and is maintained in wild-type OHCs at 1 month (white arrows). Scale bars, 50 µm (P8) and 10 μm (1 month). n = 3 biologically independent samples for each time point. b, Auditory brainstem response phenotyping of pedigree MPC173 at 9 months of age identified 17 biologically independent animals with increased hearing thresholds (red triangles) compared to their normal hearing colony mates (black triangles). n = 15 biologically independent animals. c, The mutation mapped to an 8.4-Mb region on chromosome 1 between single nucleotide polymorphism (SNP) rs31869113 and rs13475914 (chr1: 63280183–71629721), containing 66 genes. d, Detection of a non-synonymous mutation in cello. DNA sequencing identified a nucleotide transversion (c.1551C>A) in the Ikzf2 gene at codon 517, thus altering the wild-type sequence CAC, encoding histidine, to the mutant (M) sequence CAA, encoding glutamine. Electropherograms derived from a cello mutant mouse (Ikzf2cello/cello) and a wild-type colony mate (Ikzf2+/+) control showing the sequence surrounding Ikzf2 nucleotide 1551 (indicated by an arrow). e, Helios is expressed in the OHC nuclei of both Ikzf2+/+ and Ikzf2cello/cello mice at P8. n = 3 biologically independent samples per genotype. Loss of labelling when the anti-Helios antibody is ‘pre-blocked’ confirms specificity. n = 1 biologically independent sample. Scale bars, 20 µm. DC, Deiters’ cells.
Extended Data Fig. 3 The Ikzf2cello mutation disrupts homodimerization of Helios.
a, Cos-7 cells transfected with Ikzf2+- or Ikzf2cello-Myc. Nuclear localization is unaffected by the Ikzf2cello mutation. n = 2 biologically independent experiments. Scale bars, 10 µm. b, Co-immunoprecipitation (IP) of Myc-tagged (62 kDa) and GFP-tagged (88 kDa) Ikzf2+ and Ikzf2cello constructs. Transfected cell lysates were immunoprecipitated using an anti-Myc antibody and analysed by western blotting with both anti-Myc and anti-GFP antibodies. Results show that wild-type Ikzf2+ Helios can dimerize, but that dimerization is impaired by the cello mutation. LC, cell lysate loading control. c, Reciprocal immunoprecipitation reactions using an anti-GFP antibody confirm dimerization of wild-type Ikzf2+ Helios and reduced dimerization of mutant Ikzf2cello Helios. d, Quantification of co-immunoprecipitation western blots. Band intensities were determined and used to calculate the relative ratio of the co-immunoprecipitation to immunoprecipitation signal. n = 4 biologically independent experiments. Data are mean ± s.e.m. Anti-Myc IP: ***P < 0.0001 (Ikzf2+-Myc + Ikzf2+-GFP vs Ikzf2+-Myc + Ikzf2cello-GFP, vs Ikzf2cello-Myc + Ikzf2+-GFP and vs Ikzf2cello-Myc + Ikzf2cello-GFP). *P = 0.0476 (Ikzf2cello-Myc + Ikzf2+-GFP vs Ikzf2cello-Myc + Ikzf2cello-GFP). P = 0.1488 (Ikzf2+-Myc + Ikzf2cello-GFP vs Ikzf2cello-Myc + Ikzf2+-GFP). P = 0.9020 (Ikzf2+-Myc + Ikzf2cello-GFP vs Ikzf2cello-Myc + Ikzf2cello-GFP). Anti-GFP IP: ***P < 0.0001 (Ikzf2+-Myc + Ikzf2+-GFP vs Ikzf2cello-Myc + Ikzf2+-GFP, vs Ikzf2+-Myc + Ikzf2cello-GFP and vs Ikzf2cello-Myc + Ikzf2cello-GFP). *P = 0.0202 (Ikzf2cello-Myc + Ikzf2+-GFP vs Ikzf2+-Myc + Ikzf2cello-GFP) *P = 0.0346 (Ikzf2cello-Myc + Ikzf2+-GFP vs Ikzf2cello-Myc + Ikzf2cello-GFP). P = 0.9894 (Ikzf2+-Myc + Ikzf2cello-GFP vs + Ikzf2cello-Myc + Ikzf2cello-GFP) (one-way ANOVA with Tukey post hoc test). See Supplementary Fig. 1 for source images.
Extended Data Fig. 4 Auditory function and HC bundle survival in cello mice.
a, Representative click ABR waveforms for Ikzf2+/+, Ikzf2cello/+ and Ikzf2cello/cello littermates at P16. n = 4 biologically independent animals per genotype. b, c, Averaged ABR thresholds for cello mice at 1-month of age (b) and 9 months of age (c). Age-matched Ikzf2+/+ and Ikzf2cello/+ controls display thresholds within the expected range (15–30 dB SPL) at all time points tested. n = 5 biologically independent animals per genotype. Data are mean thresholds ± s.e.m. 1-month Ikzf2cello/cello vs Ikzf2+/+: ****P < 0.0001 (8 kHz, 16 kHz, 32 kHz, click). 1-month Ikzf2cello/cello vs 1 Ikzf2cello/+: ****P < 0.0001 (8 kHz, 16 kHz, 32 kHz, click). 9-month Ikzf2cello/cello vs Ikzf2+/+: ****P < 0.0001 (8 kHz, 16 kHz, 32 kHz, click). 9-month Ikzf2cello/cello vs Ikzf2cello/+: ****P < 0.0001 (8 kHz, 16 kHz, 32 kHz, click) (one-way ANOVA with Tukey post hoc test). d, OHC and IHC bundle counts for cello mice from P16 to 18 months of age. Grey, Ikzf2+/+; black, Ikzf2cello/+; red, Ikzf2cello/cello. Data are mean ± s.e.m. n.s., non-significant. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (one-way ANOVA with Tukey post hoc test). Number of biologically independent samples for OHC and IHC bundle counts are shown. See also Supplementary Table 5 and 6.
Extended Data Fig. 5 Scanning electron microscopy of cello mice and auditory function of Ikzf2cello/del890 compound heterozygotes.
a, Scanning electron micrographs of the organ of Corti of cello mice from P16 to 18 months of age. Representative images from the mid-region of the cochlear spiral are shown. Scale bars, 10 µm. n = 3 (P16 Ikzf2cello/+, P16 Ikzf2cello/cello, 1-m Ikzf2cello/cello, 9-m Ikzf2+/+, 18-m Ikzf2+/+, 18-m Ikzf2cello/+, 18-m Ikzf2cello/cello), n = 4 (P16 Ikzf2+/+, 1-m Ikzf2+/+, 3-m Ikzf2+/+, 3-m Ikzf2cello/cello, 6-m Ikzf2+/+, 6-m Ikzf2cello/cello, 9-m Ikzf2cello/+, 9-m Ikzf2cello/cello) and n = 5 (1-m Ikzf2cello/+, 3-m Ikzf2cello/+, 6-m Ikzf2cello/+) biologically independent samples. b–d, Scanning electron micrographs of OHC stereocilia bundles of cello mice at P16, showing that wild-type Ikzf2+/+ (b), Ikzf2cello+ (c) and mutant Ikzf2cello/cello (d) mice display overall expected bundle patterning. Images are from the mid-region of the cochlear spiral. Scale bars, 1 µm. n = 3 biologically independent samples for each genotype. e, The genomic and domain structure of Ikzf2del890. Black, 5′ untranslated region; light grey, N-terminal DNA-binding domain; dark grey, C-terminal dimerization domain. The Ikzf2cello mutation lies in ZnF6. The del890 mutation deletes exon 4 and the surrounding intronic sequence. f, Averaged ABR thresholds for Ikzf2cello/del890 compound heterozygotes at 1 month of age, showing increased thresholds (≥40 dB SPL) at all frequencies tested compared to Ikzf2+/+, Ikzf2cello/+ and Ikzf2del890/+ control colony mates. Data are mean ± s.e.m. n = 4 (Ikzf2+/+, Ikzf2+/del890), n = 2 (Ikzf2cello/+) and n = 5 (Ikzf2cello/del890) biologically independent samples. Ikzf2cello/del890 vs Ikzf2+/+: *P = 0.011 (8 kHz), **P = 0.002 (16 kHz), ****P < 0.0001 (32 kHz), ***P = 0.0001 (click); Ikzf2cello/del890 vs Ikzf2cello/+: P = 0.078 (8 kHz), *P = 0.034 (16 kHz), **P = 0.001 (32 kHz), **P = 0.001 (click); Ikzf2cello/del890 vs Ikzf2+/del890: *P = 0.025 (8 kHz), **P = 0.009 (16 kHz), ***P = 0.0002 (32 kHz), ***P = 0.0002 (click) (one-way ANOVA with Tukey post hoc test).
Extended Data Fig. 6 The MET and adult-like potassium currents are normal in Ikzf2cello mice.
a, b, MET currents were recorded from OHCs of P9 Ikzf2cello/cello and Ikzf2cello/+ (control) littermates. During voltage steps, hair bundles were displaced by applying a 50-Hz sinusoidal force stimuli (the driver voltage to the fluid jet is shown above the traces)39. At hyperpolarized membrane potentials (−121 mV), saturating excitatory bundle stimulation (that is, towards the taller stereocilia) elicited a large inward MET current from both Ikzf2cello/+ and Ikzf2cello/cello OHCs, whereas inhibitory bundle stimulation (that is, away from the taller stereocilia) closed the MET channels and reduced the resting current. Because the MET current reverses near 0 mV, it became outward when excitatory bundle stimulation was applied during voltage steps positive to its reversal potential. At positive membrane potentials (+99 mV), excitatory bundle stimulation now elicited similar outward MET currents with larger resting amplitudes. Arrows indicate closure of the MET channels (that is, disappearance of the resting current) during inhibitory bundle displacements, arrowheads indicate the larger resting MET current at +99 mV compared to −121 mV. c, Peak-to-peak current–voltage curves obtained from Ikzf2cello/+ (n = 10 biologically independent samples) and Ikzf2cello/cello (n = 8 biologically independent samples) OHCs at P9. The maximal MET current and the resting open probability of the MET channel were found to be similar between the two genotypes. Data are mean ± s.e.m. d, e, Total K+ currents recorded from P18 Ikzf2cello/+ control (d) and Ikzf2cello/cello mutant (e) OHCs. The size of the K+ current, which is mainly due to the negatively activated IK,n (in addition to a small delayed rectifier IK15), was smaller in Ikzf2cello/cello OHCs. f, Average peak current–voltage relationship for the total K+ current recorded from the OHCs of Ikzf2cello/+ (n = 9 OHCs from 6 biologically independent animals) and Ikzf2cello/cello (n = 7 OHCs from 5 biologically independent animals) mice at P16–P18. Data are mean ± s.e.m. g, h, After normalization to the significantly reduced surface area of Ikzf2cello/cello OHCs (for this set of experiments: Ikzf2cello/+: 14.2 ± 0.4 pF; Ikzf2cello/cello: 11.2 ± 0.5 pF; P < 0.0005), both the total IK (g) and isolated IK,n (h) were not significantly different between the two genotypes at P16–P18 (two-sided Welch’s t-test). Data are mean ± s.e.m. i, NanoString validations of genes downregulated in P8 Ikzf2cello/cello cochleae at P16, normalized to wild-type reads. Data are mean ± s.d. (n = 4 biologically independent samples per genotype). *P = 0.038 (Ppp17r1 in Ikzf2cello/cello vs Ikzf2+/+), *P = 0.037 (Ppp17r1 in Ikzf2cello/cello vs Ikzf2cello/+) (two-sided Welch’s t-test).
Extended Data Fig. 7 Transduction of cochlear hair cells using Anc80L65 and hair cell enrichment by flow cytometry.
a, Schematic representation of inner ear viral gene delivery via the posterior semicircular canal of CD-1 mice for hair cell marker immunolabelling. b, Immunolabelling for GFP in the Anc80-eGFP injected, and MYC in the Anc80-Ikzf2 injected ears, showing mainly hair cell transduction, although some MYC staining could also be observed in supporting cells (blue arrow). n = 3 biologically independent samples per condition. Nuclear MYC staining suggests proper trafficking of the MYC-tagged Helios protein in transduced cells. White arrows indicate OHCs, white arrowheads indicate IHCs. Scale bars, 10 µm. c, d, Flow cytometry of dissociated cochlear GFP-positive and tdTomato-positive cells from P8 Myo15cre/+;ROSA26CAG-tdTomato mice injected with either Anc80-eGFP (c, 2 mice) or Anc80-Ikzf2 (d, 4 mice). Cells were first gated by forward and side scatter to exclude doublets. For the Anc80-eGFP-transduced cochlear sample, transduced cells were identified based on GFP expression, and hair cells were further identified by tdTomato expression. tdTomato single-positive, GFP single-positive and tdTomato and GFP double-positive cells were collected. For the Anc80-Ikzf2-transduced cochlear sample, hair cells were gated based on tdTomato single-positive expression and collected.
Extended Data Fig. 8 Transcriptional conversion of Anc80-Ikzf2-transduced IHCs.
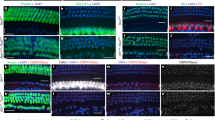
a, Heat map for the top 30 differently expressed genes between all hair cells profiled. Scaled expression values shown as z-scores, with yellow indicating higher and purple indicating lower expression than the mean. b, OHC enriched genes that are induced in Anc80-Ikzf2(+) IHCs. Anc80-Ikzf2(−) IHC (n = 34) vs Anc80-Ikzf2(+) IHC (n = 40) FDR: Pde6d = 2.03 × 10−12, Ldhb = 3.74 × 10−11. Dots represent the expression values of individual cells, with width of violins summarizing overall relative distribution of expression. c, IHC enriched genes that are highly expressed in control IHCs vs control OHCs, but are significantly reduced in Anc80-Ikzf2(+) IHCs. Anc80-Ikzf2(−) IHC (n = 34) vs Anc80-Ikzf2(+) IHC (n = 40) FDR: Fgf8 = 3.30 × 10−14, Atp2a3 = 2.46 × 10−13, Rprm = 2.27 × 10−13 (Kruskal–Wallis test followed by post hoc pairwise Wilcoxon ranked sum test adjusted for multiple comparisons). d, IHC-enriched genes that show only moderately reduced expression in Anc80-Ikzf2(+) IHCs. Anc80-Ikzf2(−) IHC (n = 34) vs Anc80-Ikzf2(+) IHC (n = 40) FDR: Shtn1 = 8.59 × 10−5, Tbx2 = 3.88 × 10−8, Cabp2 = 1.40 × 10−10 (Kruskal–Wallis test followed by post hoc pairwise Wilcoxon ranked sum adjusted for multiple comparisons). e, f, Top 20 genes negatively (e) or positively (f) correlated with Ikzf2 expression in control hair cells, shown alongside corresponding correlations of gene expression within all Anc80-Ikzf2-transduced hair cells, Anc80-Ikzf2-transduced IHCs, or Anc80-Ikzf2 transduced-OHCs. See also Extended Data Fig. 9. g, Genes that are negatively correlated with Ikzf2 (n = 20, Pearson correlation < −0.6) are not enriched in OHCs at P8 compared to all other genes detected in the RiboTag OHC dataset (background genes, n = 13,124). Genes that are positively correlated with Ikzf2 (n = 41, Pearson correlation > 0.6) are significantly enriched in OHCs at P8 compared to background genes (n = 13,103) (P = 0.025, two-sided Wilcoxon’s test). Black line represents median enrichment factor (log2 fold change), box demarcates first and third quartiles, whiskers demarcate first and third quartile ± 1.5 × IQR values, dots represent single outliers. h, One of the most differentially expressed genes observed in our scRNA-seq experiment was Fcrlb, a gene which encodes an Fc receptor like protein, and the expression of which has not been previously described in the ear. Fcrlb is significantly downregulated in Anc80-Ikzf2(+) hair cells. Anc80-Ikzf2(−) IHC (n = 34) vs Anc80-Ikzf2(+) IHC (n = 40) FDR = 4.89 × 10−6. Anc80-Ikzf2(−) OHC (n = 132) vs Anc80-Ikzf2(+) OHC (n = 148) FDR = 6.88 × 10−8 (Kruskal–Wallis test followed by post hoc pairwise Wilcoxon ranked sum test adjusted for multiple comparisons). See also Supplementary Tables 8–11.
Extended Data Fig. 9 scRNA-seq allows for high-resolution discrimination of cell types and their transcriptional changes due to overexpression of Ikzf2.
a, Custom annotation strategy with theoretical reads mapping to unambiguous regions of the various custom viral loci, as well as those regions that get discarded because of endogenous sequence similarity (that is, ambiguous reads). b, Violin plots of the overall scRNA-seq detection metrics, including number of unique molecules detected in each of the major cell type cluster identified (low Anc80-Ikzf2 expressing IHCs: viral Ikzf2 (vIk)− IHCs n = 34; low Anc80-Ikzf2 expressing OHCs: vIk− OHCs n = 132; high Anc80-Ikzf2 expressing IHCs: vIk+ IHCs n = 40; high Anc80-Ikzf2 expressing OHCs: vIk+ OHCs n = 140; and non-HCs: n = 219). c, FeaturePlots with red showing higher expression across all profiled cells, including cells identified as non-hair cells. Expression from loci captured with custom annotation shown to support cluster identification. A final labelled t-SNE plot shows all cells profiled clustered by predicted cell type. Misc, cells from all miscellaneous clusters with fewer than 5 cells; NSC, non-sensory epithelial cell; SC, organ of Corti supporting cell. Other clusters are defined by the highest differentially expressed marker gene. d, Pearson correlation scatter plots for selected genes within all profiled hair cells, hair cells from the Anc80-eGFP sample, or IHCs from the Anc80-Ikzf2 sample. e, A Pearson correlation heat map of all hair cells detected showing overall transcriptional similarities between the non-transduced IHCs and OHCs, along with the Anc80-Ikzf2-transduced IHCs and OHCs.
Extended Data Fig. 10 Ikzf2 overexpression induces prestin expression and electromotility in IHCs but does not affect hair bundle morphology.
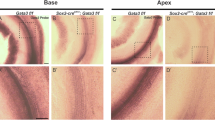
a, The OHC electromotility protein prestin is expressed in the OHCs of Ikzf2cello/cello mutants (n = 6 biologically independent samples). In addition, the pattern of prestin expression is not affected by Anc80-eGFP transduction, but is induced in Anc80-Ikzf2-transduced IHCs (n = 3 biologically independent samples per condition). Scale bars, 10 µm. b, Expression of prestin can be seen in Anc80-Ikzf2-transduced IHCs as early as P8 and up to 8 weeks of age, and overlaps with MYC staining (n = 6 biologically independent samples at P8, n = 3 biologically independent samples at 6–8 weeks). Scale bars, 20 µm. c, Scanning electron micrographs of IHC and OHC stereocilia bundles of Anc80-Ikzf2- and Anc80-eGFP-injected mice at P23 showing expected bundle patterning. Images are from the mid-basal region of the cochlear spiral. Scale bars, 1 µm. Number of biologically independent samples (P16–P23): Anc80-Ikzf2-injected cochlea n = 8, Anc80-Ikzf2 contralateral cochlea n = 6, Anc80-eGFP-injected cochlea n = 3. d, Representative traces of the voltage-dependent (nonlinear) component of the membrane capacitance (an electrical signature of electromotility) in the IHCs of Anc80-Ikzf2-injected mouse (red) and its non-injected littermate (grey). Mice were injected with Anc80-Ikzf2 at P2 and recorded at P16. e, Normalized maximal nonlinear capacitance in all recorded IHCs of mice injected with Anc80-Ikzf2 at P2 (red) at different ages after injection and their non-injected littermates (black). Each symbol represents one biologically independent cell, and the total number of cells is indicated in parentheses. Because Anc80-Ikzf2 transduction is not 100% efficient in the apical turn of the cochlea at the time points tested, some IHCs of Anc80-Ikzf2-injected mice do not show prominent nonlinear capacitance, whereas the other IHCs do. In the IHCs with maximal nonlinear capacitance of more than 0.25 pF (due to presumable Ikzf2 expression), the parameters of the Boltzmann fit were as following (mean ± s.e.m.): Qmax = 0.10 ± 0.02 pC; Vpk = −31 ± 1 mV; z = 0.91 ± 0.02; Clin = 11.7 ± 1.2 pF; ΔCsa = 0.14 ± 0.07 pF (n = 12). For information on the fitting procedure, see Methods.
Supplementary information
Supplementary Information
This file contains Supplementary Figures 1-2, Supplementary Tables 5, 6, 12 and 13 and full legends for Supplementary Tables 1-4 and 7-11
Supplementary Table 1
Average normalized RNA-seq reads for cochlear input and RiboTag OHC immunoprecipitated (IP) samples for genes detected at all five postnatal time points; P8, P14, P28, 6 weeks (wk), and 10wk
Supplementary Table 2
RiboTag RNA-seq log2 fold change enrichment and depletion of transcripts in the OHC immunoprecipitated samples compared to input at all 5 postnatal time points (EF = enrichment factor)
Supplementary Table 3
Lists of genes belonging to each of the 3 clusters of differentially expressed OHC genes, with their corresponding gene ontology (GO) terms
Supplementary Table 4
List of genes found to be OHC enriched by the RiboTag OHC dataset at 10 weeks, as well as the IHC/OHC dataset published by Liu et al
Supplementary Table 7
normalized reads and differential expression results comparing gene expression in whole cochlea of wild-type and cello mutant mice at P8
Supplementary Table 8
genes significantly differentially expressed between control Anc80-eGFP transduced IHCs and OHCs as measured by scRNA-seq
Supplementary Table 9
Results of a differential gene expression analysis between Anc80-Ikzf2(-) IHCs and OHCs compared to Anc80-Ikzf2(+) IHCs and OHCs as measured by scRNA-seq
Supplementary Table 10
results of a non-parametric analysis of variance between the four classified groups of HCs identified by scRNA-seq: Anc80-Ikzf2(-) IHCs and OHCs, Anc80-Ikzf2(+) IHCs and OHCs
Supplementary Table 11
Results of a Pearson correlation analysis of Ikzf2 and Anc80-Ikzf2 expression with all other genes in control and virally transduced HC conditions
Rights and permissions
About this article

Cite this article
Chessum, L., Matern, M.S., Kelly, M.C. et al. Helios is a key transcriptional regulator of outer hair cell maturation. Nature 563, 696–700 (2018). https://doi.org/10.1038/s41586-018-0728-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0728-4
Keywords
This article is cited by
-
Stem Cell-Based Hair Cell Regeneration and Therapy in the Inner Ear
Neuroscience Bulletin (2024)
-
Advance and Application of Single-cell Transcriptomics in Auditory Research
Neuroscience Bulletin (2023)
-
Fgf8P2A-3×GFP/+: A New Genetic Mouse Model for Specifically Labeling and Sorting Cochlear Inner Hair Cells
Neuroscience Bulletin (2023)
-
TBX2 specifies and maintains inner hair and supporting cell fate in the Organ of Corti
Nature Communications (2022)
-
Tbx2 is a master regulator of inner versus outer hair cell differentiation
Nature (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
