
Abstract
Increased storage of carbon in the oceans has been proposed as a mechanism to explain lower concentrations of atmospheric carbon dioxide during ice ages; however, unequivocal signatures of this storage have not been found1. In seawater, the dissolved gases oxygen and carbon dioxide are linked via the production and decay of organic material, with reconstructions of low oxygen concentrations in the past indicating an increase in biologically mediated carbon storage. Marine sediment proxy records have suggested that oxygen concentrations in the deep ocean were indeed lower during the last ice age, but that near-surface and intermediate waters of the Pacific Ocean—a large fraction of which are poorly oxygenated at present—were generally better oxygenated during the glacial1,2,3. This vertical opposition could suggest a minimal net basin-integrated change in carbon storage. Here we apply a dual-proxy approach, incorporating qualitative upper-water-column and quantitative bottom-water oxygen reconstructions4,5, to constrain changes in the vertical extent of low-oxygen waters in the eastern tropical Pacific since the last ice age. Our tandem proxy reconstructions provide evidence of a downward expansion of oxygen depletion in the eastern Pacific during the last glacial, with no indication of greater oxygenation in the upper reaches of the water column. We extrapolate our quantitative deep-water oxygen reconstructions to show that the respired carbon reservoir of the glacial Pacific was substantially increased, establishing it as an important component of the coupled mechanism that led to low levels of atmospheric carbon dioxide during the glacial.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

Data availability
Data generated during this study are available from https://doi.pangaea.de/10.1594/PANGAEA.891185.
Change history
21 March 2019
In this Letter, ‘δ18C’ should have been ‘δ13C’ in Fig. 3b, and the x axis should extend to 50 kyr rather than 40 kyr. This figure has been corrected online.
References
Sigman, D. M. & Boyle, E. A. Glacial/interglacial variations in atmospheric carbon dioxide. Nature 407, 859–869 (2000).
Galbraith, E. D. & Jaccard, S. L. Deglacial weakening of the oceanic soft tissue pump: global constraints from sedimentary nitrogen isotopes and oxygenation proxies. Quat. Sci. Rev. 109, 38–48 (2015).
Bradtmiller, L. I., Anderson, R. F., Sachs, J. P. & Fleisher, M. Q. A deeper respired carbon pool in the glacial equatorial Pacific Ocean. Earth Planet. Sci. Lett. 299, 417–425 (2010).
Hoogakker, B. A. A., Elderfield, H., Schmiedl, G., McCave, I. N. & Rickaby, R. E. M. Glacial–interglacial changes in bottom-water oxygen content on the Portuguese margin. Nat. Geosci. 8, 40–43 (2015).
Lu, Z. et al. Oxygen depletion recorded in upper waters of the glacial Southern Ocean. Nat. Commun. 7, 11146 (2016).
Bianchi, D., Dunne, J. P., Sarmiento, J. L. & Galbraith, E. D. Data-based estimates of suboxia, denitrification, and N2O production in the ocean and their sensitivities to dissolved O2. Global Biogeochem. Cycles 26, (2012).
Vaquer-Sunyer, R. & Duarte, C. M. Thresholds of hypoxia for marine biodiversity. Proc. Natl Acad. Sci. USA 105, 15452–15457 (2008).
Keeling, R. E., Körtzinger, A. & Gruber, N. Ocean deoxygenation in a warming world. Ann. Rev. Mar. Sci. 2, 199–229 (2010).
Lam, P. & Kuypers, M. M. M. Microbial nitrogen cycling processes in oxygen minimum zones. Ann. Rev. Mar. Sci. 3, 317–345 (2011).
Schmittner, A. & Somes, C. J. Complementary constraints from carbon (13C) and nitrogen (15N) isotopes on the glacial ocean’s soft-tissue biological pump. Paleoceanography 31, 669–693 (2016).
Schmidtko, S., Stramma, L. & Visbeck, M. Decline in global oceanic oxygen content during the past five decades. Nature 542, 335–339 (2017).
Stramma, L., Johnson, G. C., Sprintall, J. & Mohrholz, V. Expanding oxygen-minimum zones in the tropical oceans. Science 320, 655–658 (2008).
Bopp, L. et al. Multiple stressors of ocean ecosystems in the 21st century: projections with CMIP5 models. Biogeosciences 10, 6225–6245 (2013).
Long, M., Deutsch, C. & Ito, I. Finding forced trends in oceanic oxygen. Global Biogeochem. Cycles 30, 381–397 (2016).
Matsumoto, K. Biology-mediated temperature control on atmospheric pCO2 and ocean biogeochemistry. Geophys. Res. Lett. 34, L20605 (2007).
Pichevin, L. E. et al. Interhemispheric leakage of isotopically heavy nitrate in the eastern tropical Pacific during the last glacial period. Paleoceanography 25, PA1204 (2010).
Hendy, I. L. & Pedersen, T. F. Oxygen minimum zone expansion in the eastern tropical North Pacific during deglaciation. Geophys. Res. Lett. 33, L20602 (2006).
Galbraith, E. D., Kienast, M. & The NICOPP working group members. The acceleration of ocean denitrification during deglacial warming. Nat. Geosci. 6, 579–584 (2013).
Moffitt, S. E. et al. Paleoceanographic insights on recent oxygen minimum zone expansion: lessons for modern oceanography. PLoS ONE 10, e0115246 (2015).
Lu, Z., Jenkyns, H. C. & Rickaby, R. E. M. Iodine to calcium ratios in marine carbonates as a paleo-redox proxy during oceanic anoxic events. Geology 38, 1107–1110 (2010).
McCorkle, D. C. & Emerson, S. R. The relationship between pore water carbon isotopic composition and bottom water oxygen concentration. Geochim. Cosmochim. Acta 52, 1169–1178 (1988).
Karstensen, J., Stramma, L. & Visbeck, M. Oxygen minimum zones in the eastern tropical Atlantic and Pacific oceans. Prog. Oceanogr. 77, 331–350 (2008).
van Geen, A. et al. On the preservation of laminated sediments along the western margin of North America. Paleoceanography 18, 1098 (2003).
Nameroff, T. J., Calvert, E. & Murray, J. W. Glacial–interglacial variability in the eastern tropical North Pacific oxygen minimum zone recorded by redox-sensitive trace metals. Paleoceanography 19, PA1010 (2004).
Costa, K. M. et al. Productivity patterns in the Equatorial Pacific over the last 30,000 years. Global Biogeochem. Cycles 31, 850–865 (2017).
Kienast, M. et al. Eastern Pacific cooling and Atlantic overturning circulation during the last deglaciation. Nature 443, 846–849 (2006).
de la Fuente, M., Skinner, L., Calvo, E., Pelejero, C. & Cacho, I. Increased reservoir ages and poorly ventilated deep waters inferred in the glacial Eastern Equatorial Pacific. Nat. Commun. 6, 7420 (2015).
Garcia, H. et al. World Ocean Atlas 2013, Volume 3: Dissolved Oxygen, Apparent Oxygen Utilization, and Oxygen Saturation (ed. Levitus, S.) (NOAA Atlas NESDIS 75, 2013).
Stern, J. V. & Lisiecki, L. E. Termination 1 timing in radiocarbon-dated regional benthic δ18O stacks. Paleoceanography 29, 1127–1142 (2014).
Benway, H. M., Mix, A. C., Haley, B. A. & Klinkhammer, G. P. Eastern Pacific warm pool paleosalinity and climate variability: 0–30 kyr. Paleoceanography 21, PA3008 (2006).
Umling, N. E. & Thunell, R. C. Synchronous deglacial thermocline and deep-water ventilation in the eastern equatorial Pacific. Nat. Commun. 8, 14203 (2017).
Blaauw, M. & Christen, J. A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474 (2011).
Codispotti, L., Yoshinari, T. & Devol, A.H. in Respiration in Aquatic Ecosystems (eds del Giorgio, P. & Williams, P.) Ch. 12 (Oxford Univ. Press, Oxford, 2005).
Mix, A. C. et al. Proc. ODP, Init. Rep. https://doi.org/10.2973/odp.proc.ir.202.113.2003 (2003).
Eide, M., Olsen, A., Ninnemann, U. S. & Eldevik, T. A global estimate of the full oceanic 13C Suess effect since the preindustrial. Global Biogeochem. Cycles 31, 492–514 (2017).
Chance, R. et al. Seasonal and interannual variation of dissolved iodine speciation at a coastal Antarctic site. Mar. Chem. 118, 171–181 (2010).
Spokes, L. J. & Liss, P. L. Photochemically induced redox reactions in seawater, II. Nitrogen and iodide. Mar. Chem. 54, 1–10 (1996).
Chance, R., Baker, A. R., Carpenter, L. & Jickells, T. D. The distribution of iodide at the sea surface. Environ. Sci. Process Impacts 16, 1841–1859 (2014).
Fairbanks, R. G., Sverdlove, M., Free, R., Wiebe, P. H. & Bé, A. W. H. Vertical distribution and isotopic fractionation of living planktonic foraminifera in the Panama Basin. Nature 298, 841–844 (1982).
Ravelo, A. C. & Fairbanks, R. G. Oxygen isotopic composition of multiple species of planktonic foraminifera: recorders of modern photic zone temperature gradient. Paleoceanography 7, 815–831 (1992).
Farmer, E. C., Kaplan, A., de Menocal, P. B. & Lynch-Stieglitz, J. Corroborating ecological depth preferences of planktonic foraminifera in the tropical Atlantic with the stable isotope ratios of core top specimens. Paleoceanography 22, (2007).
Barker, S., Greaves, M. & Elderfield, H. A study of cleaning procedures used for foraminiferal Mg/Ca paleothermometry. Geochem. Geophys. Geosyst. 4, 8407 (2003).
Altabet, M. A. et al. The nitrogen isotope biogeochemistry of sinking particles from the margin of the Eastern North Pacific. Deep Sea Res. Part I 46, 655–679 (1999).
Deutsch, C., Sigman, D. M., Thunell, R. C., Meckler, A. N. & Haug, G. H. Isotopic constraints on glacial/interglacial changes in the oceanic nitrogen budget. Global. Biogechem. Cycles 4, 1–22 (2004).
Farrell, J. W., Pedersen, T. F., Calvert, S. E. & Nielsen, B. Glacial–interglacial changes in nutrient utilization in the equatorial Pacific Ocean. Nature 377, 514–517 (1995).
Devol, A. H. in Nitrogen in the Marine Environment 2nd edn (eds Capone, D. G. et al.) 263–301 (Academic, Burlington, 2008).
Adkins, J. F., McIntyre, K. & Schrag, D. P. The salinity, temperature, and δ18O of the glacial deep ocean. Science 298, 1769–1773 (2002).
Debelius, B., Gómez-Parra, A. & Forja, J. M. Oxygen solubility in evaporated seawater as a function of temperature and salinity. Hydrobiologia 632, 157–165 (2009).
Robinson, R. S., Martinez, P., Pena, L. D. & Cacho, I. Nitrogen isotope evidence for deglacial changes in nutrient supply in the eastern equatorial Pacific. Paleoceanography 24, PA4213 (2009).
Rafter, P. A. & Charles, C. D. Pleistocene equatorial Pacific dynamics inferred from the zonal asymmetry in sedimentary nitrogen isotopes. Paleoceanography 27, PA3102 (2012).
Boyer, T. P. et al. World Ocean Database 2013 (ed. Levitus, S.) (NOAA Atlas NESDIS 75, 2013).
Acknowledgements
This study benefited from discussions with R. Ganeshram. This work is supported by UK Natural Environment Research Council (NERC) grant NE/I020563/1 (to B.A.A.H.), National Science Foundation (NSF) grants OCE-1232620 and OCE-1736542 (to Z.L.) and Swiss National fund PP00P2_144811 (to O.C.). This research used samples and/or data provided by the Ocean Drilling Program (ODP). ODP is sponsored by the US National Science Foundation and participating countries (Natural Environment Research Council in the UK) under the management of Joint Oceanographic Institutions (JOI), Inc. M. Hall, J. Rolfe and C. Day are acknowledged for help with stable isotope analyses.
Author information
Authors and Affiliations
Contributions
B.A.A.H. and Z.L. conceived and coordinated the work. B.A.A.H., Z.L., N.U., L.J. and X.Z. carried out data analyses; O.C. carried out data synthesis. B.A.A.H., Z.L. and E.G. constructed the figures and wrote the paper, with contributions from the other co-authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
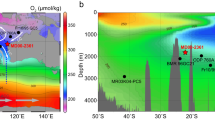
Extended Data Fig. 3 Overview and LGM evolution of carbon isotopes and oxygen concentrations in the eastern tropical Pacific.
a, Dissolved oxygen concentrations (modern: North Atlantic north of 50° N, dark blue; South Atlantic south of 50° S, light blue; southeast Pacific south of 50° S, black; southwest Pacific south of 50° S, grey; northeast Pacific north of 50° N, dark purple; northwest Pacific north of 50° N, light purple; and reconstructed for the past 40 kyr: ODP site 1242, dark green; TR163-25, light green) plotted against carbon isotopes of DIC of seawater (‰) (data from refs 28,51 using https://www.nodc.noaa.gov/OC5/SELECT/dbsearch/dbsearch.html. Square boxes represent modern values at the two sites; diamonds represent LGM values (average 18–22 kyr bp). b, Latitudinal profile of the difference in Pacific carbon isotopes between the LGM (18–22 kyr, from epifaunal benthic foraminifera) and recent (DIC) seawater carbon isotopes (extrapolated from ref. 34). Inset, histogram of LGM-DIC δ13C (waters deeper than 1.3 km) has a normal distribution (0.1‰ bin width).
Rights and permissions
About this article

Cite this article
Hoogakker, B.A.A., Lu, Z., Umling, N. et al. Glacial expansion of oxygen-depleted seawater in the eastern tropical Pacific. Nature 562, 410–413 (2018). https://doi.org/10.1038/s41586-018-0589-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0589-x
Keywords
This article is cited by
-
Southern Ocean glacial conditions and their influence on deglacial events
Nature Reviews Earth & Environment (2023)
-
A global synthesis of high-resolution stable isotope data from benthic foraminifera of the last deglaciation
Scientific Data (2023)
-
Indian Ocean glacial deoxygenation and respired carbon accumulation during mid-late Quaternary ice ages
Nature Communications (2023)
-
Planktonic foraminifera organic carbon isotopes as archives of upper ocean carbon cycling
Nature Communications (2022)
-
Deglacial restructuring of the Eastern equatorial Pacific oxygen minimum zone
Communications Earth & Environment (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
