
Abstract
The mammalian cochlea contains two types of mechanosensory hair cell that have different and critical functions in hearing. Inner hair cells (IHCs), which have an elaborate presynaptic apparatus, signal to cochlear neurons and communicate sound information to the brain. Outer hair cells (OHCs) mechanically amplify sound-induced vibrations, providing enhanced sensitivity to sound and sharp tuning. Cochlear hair cells are solely generated during development, and hair cell death—most often of OHCs—is the most common cause of deafness. OHCs and IHCs, together with supporting cells, originate in embryos from the prosensory region of the otocyst, but how hair cells differentiate into two different types is unknown1,2,3. Here we show that Insm1, which encodes a zinc finger protein that is transiently expressed in nascent OHCs, consolidates their fate by preventing trans-differentiation into IHCs. In the absence of INSM1, many hair cells that are born as OHCs switch fates to become mature IHCs. To identify the genetic mechanisms by which Insm1 operates, we compared the transcriptomes of immature IHCs and OHCs, and of OHCs with and without INSM1. In OHCs that lack INSM1, a set of genes is upregulated, most of which are normally preferentially expressed by IHCs. The homeotic cell transformation of OHCs without INSM1 into IHCs reveals a mechanism by which these neighbouring mechanosensory cells begin to differ: INSM1 represses a core set of early IHC-enriched genes in embryonic OHCs and makes them unresponsive to an IHC-inducing gradient, so that they proceed to mature as OHCs. Without INSM1, some of the OHCs in which these few IHC-enriched transcripts are upregulated trans-differentiate into IHCs, identifying candidate genes for IHC-specific differentiation.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
All data are available from the corresponding authors upon reasonable request. RNA-seq data are available for public view at the gEAR portal (https://umgear.org/).
Change history
05 December 2018
In Figs. 1e and 2g of this Letter, the labels ‘actin’ and ‘VGLUT3’, respectively, should have been in red instead of green font. This has been corrected online.
References
Puligilla, C. & Kelley, M. W. Building the world’s best hearing aid; regulation of cell fate in the cochlea. Curr. Opin. Genet. Dev. 19, 368–373 (2009).
Groves, A. K., Zhang, K. D. & Fekete, D. M. The genetics of hair cell development and regeneration. Annu. Rev. Neurosci. 36, 361–381 (2013).
Basch, M. L., Brown, R. M. II, Jen, H.-I. & Groves, A. K. Where hearing starts: the development of the mammalian cochlea. J. Anat. 228, 233–254 (2016).
Lorenzen, S. M., Duggan, A., Osipovich, A. B., Magnuson, M. A. & García-Añoveros, J. Insm1 promotes neurogenic proliferation in delaminated otic progenitors. Mech. Dev. 138, 233–245 (2015).
Gierl, M. S., Karoulias, N., Wende, H., Strehle, M. & Birchmeier, C. The zinc-finger factor Insm1 (IA-1) is essential for the development of pancreatic beta cells and intestinal endocrine cells. Genes Dev. 20, 2465–2478 (2006).
Rosenbaum, J. N., Duggan, A. & García-Añoveros, J. Insm1 promotes the transition of olfactory progenitors from apical and proliferative to basal, terminally dividing and neuronogenic. Neural Dev. 6, 6 (2011).
Yang, H., Xie, X., Deng, M., Chen, X. & Gan, L. Generation and characterization of Atoh1-Cre knock-in mouse line. Genesis 48, 407–413 (2010).
Ohyama, T. & Groves, A. K. Generation of Pax2-Cre mice by modification of a Pax2 bacterial artificial chromosome. Genesis 38, 195–199 (2004).
Ruben, R. J. Development of the inner ear of the mouse: a radioautographic study of terminal mitoses. Acta Otolaryngol. 220, 1–44 (1967).
Chen, P. & Segil, N. p27(Kip1) links cell proliferation to morphogenesis in the developing organ of Corti. Development 126, 1581–1590 (1999).
Kelley, M. W., Talreja, D. R. & Corwin, J. T. Replacement of hair cells after laser microbeam irradiation in cultured organs of corti from embryonic and neonatal mice. J. Neurosci. 15, 3013–3026 (1995).
Cox, B. C. et al. Spontaneous hair cell regeneration in the neonatal mouse cochlea in vivo. Development 141, 816–829 (2014).
Bramhall, N. F., Shi, F., Arnold, K., Hochedlinger, K. & Edge, A. S. B. Lgr5-positive supporting cells generate new hair cells in the postnatal cochlea. Stem Cell Reports 2, 311–322 (2014).
Hu, L. et al. Diphtheria toxin-induced cell death triggers Wnt-dependent hair cell regeneration in neonatal mice. J. Neurosci. 36, 9479–9489 (2016).
White, P. M., Doetzlhofer, A., Lee, Y. S., Groves, A. K. & Segil, N. Mammalian cochlear supporting cells can divide and trans-differentiate into hair cells. Nature 441, 984–987 (2006).
Mburu, P. et al. Whirlin complexes with p55 at the stereocilia tip during hair cell development. Proc. Natl Acad. Sci. USA 103, 10973–10978 (2006).
Flores, E. N. et al. A non-canonical pathway from cochlea to brain signals tissue-damaging noise. Curr. Biol. 25, 606–612 (2015).
Breslin, M. B., Zhu, M., Notkins, A. L. & Lan, M. S. Neuroendocrine differentiation factor, IA-1, is a transcriptional repressor and contains a specific DNA-binding domain: identification of consensus IA-1 binding sequence. Nucleic Acids Res. 30, 1038–1045 (2002).
Jia, S., Wildner, H. & Birchmeier, C. Insm1 controls the differentiation of pulmonary neuroendocrine cells by repressing Hes1. Dev. Biol. 408, 90–98 (2015).
Lan, M. S. & Breslin, M. B. Structure, expression, and biological function of INSM1 transcription factor in neuroendocrine differentiation. FASEB J. 23, 2024–2033 (2009).
Liu, W.-D., Wang, H.-W., Muguira, M., Breslin, M. B. & Lan, M. S. INSM1 functions as a transcriptional repressor of the neuroD/β2 gene through the recruitment of cyclin D1 and histone deacetylases. Biochem. J. 397, 169–177 (2006).
Wang, H.-W. et al. Identification of an INSM1-binding site in the insulin promoter: negative regulation of the insulin gene transcription. J. Endocrinol. 198, 29–39 (2008).
Osipovich, A. B. et al. Insm1 promotes endocrine cell differentiation by modulating the expression of a network of genes that includes Neurog3 and Ripply3. Development 141, 2939–2949 (2014).
Jia, S. et al. Insm1 cooperates with Neurod1 and Foxa2 to maintain mature pancreatic β-cell function. EMBO J. 34, 1417–1433 (2015).
Cai, T. et al. Characterization of the transcriptome of nascent hair cells and identification of direct targets of the Atoh1 transcription factor. J. Neurosci. 35, 5870–5883 (2015).
Scheffer, D. I., Shen, J., Corey, D. P. & Chen, Z.-Y. Gene expression by mouse inner ear hair cells during development. J. Neurosci. 35, 6366–6380 (2015).
Yang, T. et al. Expression and localization of CaBP Ca2+ binding proteins in the mouse cochlea. PLoS One 11, e0147495 (2016).
Jacques, B. E., Montcouquiol, M. E., Layman, E. M., Lewandoski, M. & Kelley, M. W. Fgf8 induces pillar cell fate and regulates cellular patterning in the mammalian cochlea. Development 134, 3021–3029 (2007).
Pirvola, U. et al. FGFR1 is required for the development of the auditory sensory epithelium. Neuron 35, 671–680 (2002).
Liu, H. et al. Characterization of transcriptomes of cochlear inner and outer hair cells. J. Neurosci. 34, 11085–11095 (2014).
Liu, P., Jenkins, N. A. & Copeland, N. G. A highly efficient recombineering-based method for generating conditional knockout mutations. Genome Res. 13, 476–484 (2003).
Kanki, H., Suzuki, H. & Itohara, S. High-efficiency CAG-FLPe deleter mice in C57BL/6J background. Exp. Anim. 55, 137–141 (2006).
Neely, S. T. & Liu, Z. EMAV: Otoacoustic emission averager. Omaha, NE: Technical Memo No 17. Boy’s Town National Research Hospital (1994).
Neely, S. T. & Stevenson, R. SysRes. Omaha, NE: Tech Memo No. 1. BoysTown National Research Hospital NE (1992).
Pearce, M., Richter, C. P. & Cheatham, M. A. A reconsideration of sound calibration in the mouse. J. Neurosci. Methods 106, 57–67 (2001).
Cheatham, M. A. et al. Loss of the tectorial membrane protein CEACAM16 enhances spontaneous, stimulus-frequency, and transiently evoked otoacoustic emissions. J. Neurosci. 34, 10325–10338 (2014).
Montgomery, S. C. & Cox, B. C. Whole mount dissection and immunofluorescence of the adult mouse cochlea. J. Vis. Exp. 107, e53561 (2016).
Delmaghani, S. et al. Hypervulnerability to sound exposure through impaired adaptive proliferation of peroxisomes. Cell 163, 894–906 (2015).
Nagy, A., Gertsenstein, M., Vintersten, K. & Behringer, R. Staining frozen mouse embryo sections for β-galactosidase (lacZ) activity. CSH Protoc. 2007, pdb.prot4726 (2007).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Anders, S., Pyl, P. T. & Huber, W. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Yang, H. et al. Gfi1-Cre knock-in mouse line: A tool for inner ear hair cell-specific gene deletion. Genesis 48, 400–406 (2010).
Acknowledgements
NU core facilities used were NUSeq, TTML, CAM and FC (partially supported by CA060553 to RHLCCC). We thank A. Groves for protocols and advice and D. He for original databases of microarray results. Supported by NIH grants DC015903, DC000089 and DC012483.
Reviewer information
Nature thanks S. Heller and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
J.G.-A. and A.D. conceived the project. T.W., S.M.L., J.A.C., C.Z.F., A.K.H., F.M., J.C.C., M.A.C., A.D. and J.G.-A. performed experiments. T.W., S.M.L., J.A.C., C.Z.F., M.J.S., M.A.C., A.D. and J.G.-A. analysed data. J.G.-A. and T.W. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Generation of a conditional KO allele of Insm1.
a, We generated a targeting construct in which the sole exon of Insm1 (green rectangle, with the coding sequence in dark green and the UTRs in light green) has a loxP site (purple triangle) inserted in a poorly conserved area of its 5′UTR and another loxP site downstream of the Insm1 gene. The construct also incorporates a neomycin resistance cassette (NEO, blue) surrounded by Frt sites (red triangles) and a thymidine kinase cassette (HSV-TK; orange), which are used to select for recombination events after gene targeting. b, We screened 439 clones and identified 5 recombinants (1 non-recombinant wild type, B5, and 3 recombinants, B6, E10 and B3, are shown) with PCR using primers indicated in a (arrows). The expected size for the wild-type allele using primers WT to R2 is 6,163 bp. The expected size for recombinant clones using CKO-reverse is 6,145 bp. c, Selected embryonic stem (ES) cell clones were additionally screened for homologous recombination upstream of the first loxP site by Southern blotting after digestion with SpeI and using the 5′ probe indicated in a6. Southern blotting was performed twice. The expected band sizes for wild-type and conditional KO alleles are 18,162 bp and 14,333 bp, respectively. From one of these clones (B3) we generated first chimeric mice and then mice with floxed alleles of Insm1 (obtained by crossing the chimaeras with mice expressing the FlpE recombinase (B6-Tg(CAG-FLPe)36, which deleted the NEO cassette flanked by FRT sites). Homozygous Insm1F/F mice are viable, demonstrating that the loxP insertions do not interfere with the vital functions of Insm1 and hence may be used for its conditional ablation. Co-expression with Cre recombinase generates an Insm1 KO allele lacking its entire coding sequence.
Extended Data Fig. 2 Conditional ablation of Insm1 in cochleae.
In situ hybridization for Insm1 transcripts on cryosections of embryonic E16.5 and neonatal (P0 and P1) cochleae. a–c, In control cochleae (top), Insm1 is expressed in all OHCs (72/72 OHCs from 3 animals) and spiral ganglion (SG; white arrows) at E16.5. However, no Insm1 was detected in the organs of Corti from apical turns, in which recognizable hair cells have not yet appeared (a, c; asterisks). By postnatal age P0–P1, Insm1 mRNA is present in 90% of OHCs (94/105 OHCs from 2 animals), and it is undetectable in spiral ganglion (b, d). a, b, Bottom, in TgPax2Cre/+;Insm1F/F mice, Insm1 mRNA is undetectable in spiral ganglion and in all OHCs from E16.5 (0/69 OHCs from 2 mice; a) and P0 (0/42 OHCs from 1 mouse; b) cochleae. c, d, Bottom, in Atoh1Cre/+;Insm1Flox/Flox cochleae, Insm1 mRNA is present in spiral ganglion and 43% of OHCs (18/42 OHCs from 1 animal) at E16.5 (c), reduced to 7% (4/54) at E17.5 (not shown), and entirely absent all OHCs (0/60 OHCs from 1 animal) and spiral ganglion by postnatal day P1 (d). For quantification at E16.5, we did not include organs of Corti from apical turns, which do not yet have recognizable hair cells. Filled arrowheads indicate organs of Corti with Insm1 expression, and empty arrowheads indicate organs of Corti without Insm1 expression. Scale bars, 200 µm.
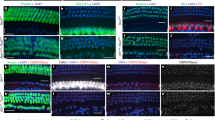
Extended Data Fig. 3 Conditional ablation of Insm1 in hair cells and spiral ganglia neurons using TgPax2Cre causes hearing impairment and the appearance of IHC-like cells in place of OHCs.
a, b, Hearing thresholds determined by ABRs (a) and DPOAEs (b) of TgPax2Cre/+;Insm1F/F mice at age P35–P46 (black traces; n = 4, 4 females) and control littermates (red traces; n = 5, 2 males and 3 females). The fact that shifts in ABR threshold are larger than shifts in DPOAE threshold may indicate an additional contribution to hearing impairment of the spiral ganglion neurons lacking INSM1 in TgPax2Cre/+;Insm1F/F cochlea. c, Despite the prevalence of OHCs with IHC characteristics (oc-IHCs) in TgPax2Cre/+;Insm1F/F cochleae (46.0 ± 5.64% (mean ± s.d.), n = 3 mice; Fig. 1m, n), these mice have the same density of IHCs (9.87 ± 2.41 cells per 100 µm along the organ of Corti; n = 3) as littermate controls (TgPax2Cre/+;Insm1F/+ and Insm1F/F; 10.54 ± 1.67 cells per 100 µm; n = 3) suggesting that oc-IHCs are not IHCs displaced from the inner to the outer compartment. d, There is no OHC loss in TgPax2Cre/+;Insm1F/F mice at ~P14–P16. Densities of oc-HCs do not differ between TgPax2Cre/+;Insm1F/F mice (OHCs and oc-IHCs) and their littermate controls (OHCs only) (29.88 ± 7.45 cells per 100 µm along the organ of Corti of TgPax2Cre/+;Insm1F/F mice, n = 3; 34.3 ± 6.92 cells per 100 µm in TgPax2Cre/+;Insm1F/+ and Insm1F/F littermate controls, n = 3). e, The number of oc-HCs per IHC does not differ between TgPax2Cre/+;Insm1F/F mice and their littermate controls (3.03 ± 0.25 OHCs plus oc-IHCs per IHC in TgPax2Cre/+;Insm1F/F mice, n = 3; 3.24 ± 0.21 OHCs per IHC in TgPax2Cre/+;Insm1F/+ and Insm1F/F littermate controls, n = 3). One-tailed Student’s t-tests were used in c–e, n is number of mice. Statistical significance is defined as P < 0.05. f, Immunofluorescence for the IHC-enriched calmodulin (green) and hair cell marker myosin VIIa (white) on whole-mount organs of Corti from mid-cochlear positions at ages ~P14–P16 confirmed that many TgPax2Cre/+;Insm1F/F oc-HCs had IHC characteristics, in addition to having the flask shape and large nuclei of IHCs (blue, DAPI, marked with asterisks), as well as lacking prestin and expressing VGLUT3 (Fig. 1m). Scale bars, 10 µm. Biological replicates were used for all experiments and similar immunohistochemistry results obtained from three or more mice per genotype.
Extended Data Fig. 4 IHC-like cells in the outer compartment (oc-IHCs) result from OHC misdifferentiation in the absence of INSM1, not from IHC displacement or from trans-differentiation of supporting cells.
a, Atoh1Cre/+;Insm1F/F mice have the same density of IHCs (11.2 ± 1.2 (mean ± s.d.) cells per 100 µm; n = 12) as littermate controls (Atoh1Cre/+;Insm1+/F and Insm1F/F; 11.5 ± 1.3 cells per 100 µm; n = 13). b, There is no loss of OHCs in Atoh1Cre/+;Insm1F/F mice up to P34. Densities of oc-HCs do not differ between Atoh1Cre/+;Insm1F/F mice (OHCs + oc-IHCs) and their littermate controls (OHCs only) (34.6 ± 3.8 cells per 100 µm in Atoh1Cre/+;Insm1F/F mice, n = 9; 37.3 ± 4.5 cells per 100 µm in Atoh1Cre/+;Insm1+/F and Insm1F/F littermate controls, n = 10). c, The number of oc-HCs per IHC do not differ significantly between Atoh1Cre/+;Insm1F/F mice and their littermate controls (3.1 ± 0.3 OHCs and oc-IHCs per IHC in Atoh1Cre/+;Insm1F/F mice, n = 9; 3.3 ± 0.2 OHCs per IHC in Atoh1Cre/+;Insm1+/F and Insm1F/F littermate controls, n = 10). The criteria for identification of oc-IHCs in Atoh1Cre/+;Insm1F/F mice were the presence in the outer compartment of hair cells expressing IHC markers (VGLUT3, high levels of calmodulin and/or nuclear CtBP2), lacking OHC markers (oncomodulin and/or prestin) and with a shape (determined by myosin VIIa immunoreactivity) like that of IHCs. Mice used for hair cell counts were P0–P34. One-tailed Student’s t-tests were used in a–c; n is number of mice. Statistical significance is defined as P < 0.05. d, SOX2 immunoreactivity, which labels the nuclei of cochlear supporting cells and, under certain conditions, of hair cells trans-differentiated from them postnatally, was not present in cells of the OHC region in Atoh1Cre/+;Insm1F/F pups (0/95 OHCs at P0, 0/42 OHCs at P2, and 0/39 OHCs at P5). e, To track postnatal cell proliferation in the organ of Corti, neonatal mice were injected twice daily with the thymidine analogue 5-ethynyl-2′-deoxyuridine (EdU) from P0 to P5 or P8. The lack of EdU in any hair cell from in Atoh1Cre/+;Insm1F/F mice (0/77 oc-HCs at P5 and 0/40 oc-HCs at P8) confirmed that these cells, including oc-IHCs, were not produced from postnatally dividing supporting cells. Unless otherwise noted, images are from mid cochlear positions. Hair cells were identified by myosin VIIa immunoreactivity, phalloidin, DAPI and Hoechst. Scale bars, 10 µm. Biological replicates were used for all experiments and similar immunohistochemistry results obtained from three or more mice per genotype.
Extended Data Fig. 5 Conditional ablation of Insm1 in hair cells using Gfi1-Cre causes no hearing impairment, and results in only very few oc-IHCs.
We generated a conditional knockout of Insm1 in hair cells using Gfi1-Cre, in which the expression of cre recombinase coincided with that of Insm1. a, b, Hearing thresholds determined by ABRs (a) and DPOAEs (b) of Gfi1Cre/+;Insm1F/F mice at age P30–P35 (black traces; n = 6, 4 males and 2 females) and control littermates (red traces; n = 6; 1 Insm1F/F, 1 Insm1F/+ and 4 Gfi1Cre/+;Insm1F/+; 4 males and 2 females). There is no significant difference in ABR and DPOAE thresholds at any frequency tested between Gfi1Cre/+;Insm1F/F mice and their control littermates. c, Immunohistochemistry in whole-mount organs of Corti from mid-cochlear positions of P34 mice tested for hearing in a, b revealed that Gfi1Cre/+;Insm1F/F mice had normal IHCs expressing high levels of calmodulin. However, very few oc-HCs also expressed calmodulin at high levels, oncomodulin at low levels, and had a round, flask shape similar to that of IHCs (0.78%; 5/526 OHCs from 2 mice). Because in these Gfi1Cre/+;Insm1F/F mice the onset of Cre recombinase expression coincides with that of Insm1 (E15.5–E17.5)43, their nascent OHCs will express Insm1 for at least several hours. This result indicates that brief expression of Insm1 is sufficient to promote proper OHC differentiation. Scale bars, 10 µm. Biological replicates were used for all experiments and similar immunohistochemistry results obtained from three or more mice per genotype.
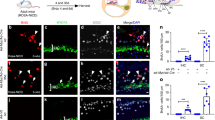
Extended Data Fig. 6 FACS purification of and RNA extraction from OHCs from an E18.5 Insm1GFP.Cre/+ embryo.
a–c, Forward and side light scattering were used to exclude dead cells and debris (a) and aggregates (≥2 cells) (b, c). d, Live cells were gated in green and red (PerCP-Cy5, to assess autofluorescence) channels to define the GFP+ (green dots) and GFP– (red dots) sorting windows. e, Myosin VIIa immunoreactivity and DAPI stain of cells collected through cytospinning after FACS confirm that most of the 547 sorted GFP+ cells are hair cells. This verification was done on all hair cell pools sorted (three pools per genotype). Inset is a representative merged image of one sorted OHC at high magnification. In this pool, no autofluorescent cells were collected. f, RT–qPCR after cell sorting (mean ± s.e.m.) reveals that, compared with GFP– cells, GFP+ cells express the hair cell marker gene Myo7a and not the supporting cell marker genes S100 and Hes5. g, To ensure the quality of the extracted RNA, the RIN score was determined using a BioAnalyzer. g, Similar RIN scores were obtained from all pools of OHCs examined (including the three per genotype used for RNA-seq in Fig. 4a).
Supplementary information
Supplementary Information
This file contains a guide to the Supplementary Tables as well as the full Table legends.
Supplementary Tables
This file contains Supplementary Tables 1-8 – see Supplementary Information document for full guide.
Rights and permissions
About this article

Cite this article
Wiwatpanit, T., Lorenzen, S.M., Cantú, J.A. et al. Trans-differentiation of outer hair cells into inner hair cells in the absence of INSM1. Nature 563, 691–695 (2018). https://doi.org/10.1038/s41586-018-0570-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0570-8
Keywords
This article is cited by
-
Fgf8P2A-3×GFP/+: A New Genetic Mouse Model for Specifically Labeling and Sorting Cochlear Inner Hair Cells
Neuroscience Bulletin (2023)
-
TBX2 specifies and maintains inner hair and supporting cell fate in the Organ of Corti
Nature Communications (2022)
-
Tbx2 is a master regulator of inner versus outer hair cell differentiation
Nature (2022)
-
Advancing discovery in hearing research via biologist-friendly access to multi-omic data
Human Genetics (2022)
-
Characterization of the development of the mouse cochlear epithelium at the single cell level
Nature Communications (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
