
Abstract
Eukaryotic cells traffic proteins and lipids between different compartments using protein-coated vesicles and tubules. The retromer complex is required to generate cargo-selective tubulovesicular carriers from endosomal membranes1,2,3. Conserved in eukaryotes, retromer controls the cellular localization and homeostasis of hundreds of transmembrane proteins, and its disruption is associated with major neurodegenerative disorders4,5,6,7. How retromer is assembled and how it is recruited to form coated tubules is not known. Here we describe the structure of the retromer complex (Vps26–Vps29–Vps35) assembled on membrane tubules with the bin/amphiphysin/rvs-domain-containing sorting nexin protein Vps5, using cryo-electron tomography and subtomogram averaging. This reveals a membrane-associated Vps5 array, from which arches of retromer extend away from the membrane surface. Vps35 forms the ‘legs’ of these arches, and Vps29 resides at the apex where it is free to interact with regulatory factors. The bases of the arches connect to each other and to Vps5 through Vps26, and the presence of the same arches on coated tubules within cells confirms their functional importance. Vps5 binds to Vps26 at a position analogous to the previously described cargo- and Snx3-binding site, which suggests the existence of distinct retromer-sorting nexin assemblies. The structure provides insight into the architecture of the coat and its mechanism of assembly, and suggests that retromer promotes tubule formation by directing the distribution of sorting nexin proteins on the membrane surface while providing a scaffold for regulatory-protein interactions.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

Data availability
Cryo-ET structures and representative tomograms have been deposited in the Electron Microscopy Data Bank (EMDB) under accession codes EMD-0154, EMD-0155, EMD-0156, EMD-0157, EMD-0158, EMD-0159, EMD-0160, EMD-0161, EMD-0162 and EMD-0163, and the molecular model in the Protein Data Bank (PDB) under accession code 6H7W. Coordinates and structure factors for Vps29 are in the PDB under accession code 5W8M, with raw data available at https://espace.library.uq.edu.au/view/UQ:682629. Gel source data can be found in Supplementary Fig. 1.
References
Burd, C. & Cullen, P. J. Retromer: a master conductor of endosome sorting. Cold Spring Harb. Perspect. Biol. 6, a016774 (2014).
Seaman, M. N. The retromer complex – endosomal protein recycling and beyond. J. Cell Sci. 125, 4693–4702 (2012).
Trousdale, C. & Kim, K. Retromer: structure, function, and roles in mammalian disease. Eur. J. Cell Biol. 94, 513–521 (2015).
McMillan, K. J., Korswagen, H. C. & Cullen, P. J. The emerging role of retromer in neuroprotection. Curr. Opin. Cell Biol. 47, 72–82 (2017).
Small, S. A. & Petsko, G. A. Retromer in Alzheimer disease, Parkinson disease and other neurological disorders. Nat. Rev. Neurosci. 16, 126–132 (2015).
Vilariño-Güell, C. et al. VPS35 mutations in Parkinson disease. Am. J. Hum. Genet. 89, 162–167 (2011).
Zimprich, A. et al. A mutation in VPS35, encoding a subunit of the retromer complex, causes late-onset Parkinson disease. Am. J. Hum. Genet. 89, 168–175 (2011).
Feng, S. et al. The sorting nexin 3 retromer pathway regulates the cell surface localization and activity of a Wnt-activated polycystin channel complex. J. Am. Soc. Nephrol. 28, 2973–2984 (2017).
Bean, B. D., Davey, M. & Conibear, E. Cargo selectivity of yeast sorting nexins. Traffic 18, 110–122 (2017).
Gallon, M. et al. A unique PDZ domain and arrestin-like fold interaction reveals mechanistic details of endocytic recycling by SNX27-retromer. Proc. Natl Acad. Sci. USA 111, E3604–E3613 (2014).
Zhang, P., Wu, Y., Belenkaya, T. Y. & Lin, X. SNX3 controls Wingless/Wnt secretion through regulating retromer-dependent recycling of Wntless. Cell Res. 21, 1677–1690 (2011).
Harterink, M. et al. A SNX3-dependent retromer pathway mediates retrograde transport of the Wnt sorting receptor Wntless and is required for Wnt secretion. Nat. Cell Biol. 13, 914–923 (2011).
Mim, C. et al. Structural basis of membrane bending by the N-BAR protein endophilin. Cell 149, 137–145 (2012).
Pylypenko, O., Lundmark, R., Rasmuson, E., Carlsson, S. R. & Rak, A. The PX-BAR membrane-remodeling unit of sorting nexin 9. EMBO J. 26, 4788–4800 (2007).
Seaman, M. N., McCaffery, J. M. & Emr, S. D. A membrane coat complex essential for endosome-to-Golgi retrograde transport in yeast. J. Cell Biol. 142, 665–681 (1998).
Nothwehr, S. F., Bruinsma, P. & Strawn, L. A. Distinct domains within Vps35p mediate the retrieval of two different cargo proteins from the yeast prevacuolar/endosomal compartment. Mol. Biol. Cell 10, 875–890 (1999).
Lucas, M. et al. Structural mechanism for cargo recognition by the retromer complex. Cell 167, 1623–1635.e14 (2016).
Hierro, A. et al. Functional architecture of the retromer cargo-recognition complex. Nature 449, 1063–1067 (2007).
Norwood, S. J. et al. Assembly and solution structure of the core retromer protein complex. Traffic 12, 56–71 (2011).
Frost, A. et al. Structural basis of membrane invagination by F-BAR domains. Cell 132, 807–817 (2008).
Kvainickas, A. et al. Cargo-selective SNX-BAR proteins mediate retromer trimer independent retrograde transport. J. Cell Biol. 216, 3677–3693 (2017).
Simonetti, B., Danson, C. M., Heesom, K. J. & Cullen, P. J. Sequence-dependent cargo recognition by SNX-BARs mediates retromer-independent transport of CI-MPR. J. Cell Biol. 216, 3695–3712 (2017).
Jia, D. et al. Structural and mechanistic insights into regulation of the retromer coat by TBC1d5. Nat. Commun. 7, 13305 (2016).
Hesketh, G. G. et al. VARP is recruited on to endosomes by direct interaction with retromer, where together they function in export to the cell surface. Dev. Cell 29, 591–606 (2014).
Dodonova, S. O. et al. 9Å structure of the COPI coat reveals that the Arf1 GTPase occupies two contrasting molecular environments. eLife 6, e26691 (2017).
Bykov, Y. S. et al. The structure of the COPI coat determined within the cell. eLife 6, e32493 (2017).
Adam, J., Basnet, N. & Mizuno, N. Structural insights into the cooperative remodeling of membranes by amphiphysin/BIN1. Sci. Rep. 5, 15452 (2015).
Daum, B. et al. Supramolecular organization of the human N-BAR domain in shaping the sarcolemma membrane. J. Struct. Biol. 194, 375–382 (2016).
Dodonova, S. O. et al. A structure of the COPI coat and the role of coat proteins in membrane vesicle assembly. Science 349, 195–198 (2015).
Battye, T. G., Kontogiannis, L., Johnson, O., Powell, H. R. & Leslie, A. G. iMOSFLM: a new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallogr. D 67, 271–281 (2011).
Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D 62, 72–82 (2006).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D 66, 213–221 (2010).
Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. J. Struct. Biol. 152, 36–51 (2005).
Hagen, W. J. H., Wan, W. & Briggs, J. A. G. Implementation of a cryo-electron tomography tilt-scheme optimized for high resolution subtomogram averaging. J. Struct. Biol. 197, 191–198 (2017).
Grant, T. & Grigorieff, N. Measuring the optimal exposure for single particle cryo-EM using a 2.6 Å reconstruction of rotavirus VP6. eLife 4, e06980 (2015).
Kremer, J. R., Mastronarde, D. N. & McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. J. Struct. Biol. 116, 71–76 (1996).
Xiong, Q., Morphew, M. K., Schwartz, C. L., Hoenger, A. H. & Mastronarde, D. N. CTF determination and correction for low dose tomographic tilt series. J. Struct. Biol. 168, 378–387 (2009).
Rohou, A. & Grigorieff, N. CTFFIND4: Fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Förster, F., Medalia, O., Zauberman, N., Baumeister, W. & Fass, D. Retrovirus envelope protein complex structure in situ studied by cryo-electron tomography. Proc. Natl Acad. Sci. USA 102, 4729–4734 (2005).
Nickell, S. et al. TOM software toolbox: acquisition and analysis for electron tomography. J. Struct. Biol. 149, 227–234 (2005).
Castaño-Díez, D., Kudryashev, M., Arheit, M. & Stahlberg, H. Dynamo: a flexible, user-friendly development tool for subtomogram averaging of cryo-EM data in high-performance computing environments. J. Struct. Biol. 178, 139–151 (2012).
Schur, F. K. et al. An atomic model of HIV-1 capsid-SP1 reveals structures regulating assembly and maturation. Science 353, 506–508 (2016).
Trabuco, L. G., Villa, E., Mitra, K., Frank, J. & Schulten, K. Flexible fitting of atomic structures into electron microscopy maps using molecular dynamics. Structure 16, 673–683 (2008).
Yang, J. et al. The I-TASSER Suite: protein structure and function prediction. Nat. Methods 12, 7–8 (2015).
Biasini, M. et al. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 42, W252–W258 (2014).
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Goddard, T. D. et al. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 27, 14–25 (2018).
Schaffer, M. et al. Cryo-focused ion beam sample preparation for imaging vitreous cells by cryo-electron tomography. Bio Protoc. 5, e1575 (2015).
Schaffer, M. et al. Optimized cryo-focused ion beam sample preparation aimed at in situ structural studies of membrane proteins. J. Struct. Biol. 197, 73–82 (2017).
Albert, S. et al. Proteasomes tether to two distinct sites at the nuclear pore complex. Proc. Natl Acad. Sci. USA 114, 13726–13731 (2017).
Wan, W. et al. Structure and assembly of the Ebola virus nucleocapsid. Nature 551, 394–397 (2017).
Bärlocher, K. et al. Structural insights into Legionella RidL-Vps29 retromer subunit interaction reveal displacement of the regulator TBC1D5. Nat. Commun. 8, 1543 (2017).
Romano-Moreno, M. et al. Molecular mechanism for the subversion of the retromer coat by the Legionella effector RidL. Proc. Natl Acad. Sci. USA 114, E11151–E11160 (2017).
Harrison, M. S. et al. A mechanism for retromer endosomal coat complex assembly with cargo. Proc. Natl Acad. Sci. USA 111, 267–272 (2014).
Liu, T. T., Gomez, T. S., Sackey, B. K., Billadeau, D. D. & Burd, C. G. Rab GTPase regulation of retromer-mediated cargo export during endosome maturation. Mol. Biol. Cell 23, 2505–2515 (2012).
Purushothaman, L. K., Arlt, H., Kuhlee, A., Raunser, S. & Ungermann, C. Retromer-driven membrane tubulation separates endosomal recycling from Rab7/Ypt7-dependent fusion. Mol. Biol. Cell 28, 783–791 (2017).
Arlt, H., Reggiori, F. & Ungermann, C. Retromer and the dynamin Vps1 cooperate in the retrieval of transmembrane proteins from vacuoles. J. Cell Sci. 128, 645–655 (2015).
Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 16, 10881–10890 (1988).
Robert, X. & Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 42, W320–W324 (2014).
Acknowledgements
We acknowledge support from the University of Queensland Remote Operation Crystallisation Facility, the Centre for Microscopy and Microanalysis (CMM), and the Australian Synchrotron. We thank S. Norwood for assistance and discussion, J. Plitzko and W. Baumeister for enabling in situ cryo-ET, and W. Hagen for supporting data collection. Work in B.M.C.’s laboratory was supported by the Australian Research Council (DP160101743) and National Health and Medical Research Council (NHMRC) (APP1042082; APP1058734). M.S. and B.J.E. were supported by the Max Planck Society. R.D.T. and B.M.C. are supported by NHMRC Senior Research Fellowships (APP1041929; APP1136021). N.L. and D.J.O. are supported by Wellcome Trust grant 207455/Z/17/Z. J.A.G.B., Y.B. and O.K. were supported by the European Molecular Biology Laboratory (EMBL) and the Medical Research Council (MC_UP_1201/16).
Reviewer information
Nature thanks K. Grünewald and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
O.K., N.L., R.D.T., D.J.O., J.A.G.B. and B.M.C. designed the project. O.K. performed cryo-ET, tomogram reconstruction, subtomogram averaging and model building with assistance from J.A.G.B. O.K. and N.L. performed electron microscopy sample preparation and screening. N.L. performed biochemical characterization and protein preparation, crystal structure determination and assisted with tomogram reconstruction. N.A. assisted N.L. with preliminary electron microscopy sample screening. Y.S.B. performed image analysis for in situ cryo-ET. M.S. and B.D.E. performed in situ cryo-ET. O.K., N.L., J.A.G.B. and B.M.C. prepared the manuscript with input from all authors in the final version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 The retromer–Vps5 complex in solution and binding to membranes.
a, Retromer forms a stable complex in solution. Fractions containing retromer (Vps35, Vps26 and Vps29) after gel-filtration on a Superdex 200 column analysed by Coomassie-stained SDS–PAGE. b, Gel-filtration profile of Vps5 and multiangle laser light scattering analysis of molecular weight. Mean molecular weight and standard deviation from three independent gel-filtration experiments are shown. The expected molecular weight of Vps5 monomer is 67 kDa, so the observed molecular weight of 129 kDa indicates formation of a homodimer. c, A Coomassie-stained gel from SDS–PAGE of Vps5 fractions from b, indicating the formation of a homodimer. d, Vps5 binds to retromer in solution. SDS–PAGE gels of GST–Vps5 and of retromer are shown under ‘input’. Retromer was incubated with GST-tagged Vps5 or GST baits, and the resultant complex was isolated on glutathione sepharose beads (‘pull-down’). At the bottom is the intact PAGE gel from which the input and pull-down lanes were extracted. e, Membrane recruitment of retromer is dependent on Vps5. GST–Vps5 alone, GST–Vps5 with retromer complex and retromer complex alone were incubated with liposomes and pelleted to isolate the liposome-bound protein fraction. The supernatant (S) and pelleted fraction (P) were compared with Coomassie-stained SDS–PAGE gels. POPC/POPE liposomes were used as a negative control. Vps5 is efficiently pelleted by Folch brain extract liposomes, and the introduction of PI3P does not increase the amount of pelleted protein. The retromer complex shows no membrane association on its own, but is recruited to Folch membranes and membranes containing both Folch extract and PI3P when it interacts with Vps5. f, Retromer promotes tubule formation by Vps5. Characteristic cryo-electron microscopy images at medium (left) and high (right) magnification of Folch liposomes incubated either with Vps5 alone (top) or in the presence of the retromer complex (bottom). Data shown in all panels are representative of at least three independent experiments.
Extended Data Fig. 2 Overview of the subtomogram-averaging procedure.
Stages in the subtomogram averaging procedure are shown from top to bottom. Key steps are illustrated by average volumes (grey) overlaid with the corresponding alignment mask (gold). Alignment masks are shown at a 0.5 value threshold. Volumes in a–c are low-pass-filtered to 50 Å. a, The final iteration of the reference-free subtomogram-averaging procedure independently conducted in bin4 tomograms that were acquired at −2.5 mm (left) and −5.5 mm (right) defoci, filtered to 50 Å resolution. b, The average of the references shown in a. c, The volume from b was rotated to place either the apex of the arch (left) or the base of the arch (right) in the box centre, two-fold symmetrized, and filtered to 50 Å resolution. These two volumes were used as starting references for further alignments. d, The references after alignment at bin8. e, The references after alignment at bin2. After subtomogram-averaging convergence in bin2, focused alignment was conducted on individual structural features. f, The final maps for alignments focused on the regions within the gold alignment masks (boxes 1–3, arch structure; boxes 4 and 5, base structure).
Extended Data Fig. 3 Cryo-ET map and reconstruction resolution.
a, Mask-corrected FSC curves for each of the final focused maps shown in Extended Data Fig. 2f. The overall resolution at the 0.143 criterion is marked. b, Sharpened maps coloured by local resolution according to the indicated colour map determined by FSC within a moving local mask. Arrowheads indicate an unassigned density, which may correspond to a helical element in loop 305–387 of Vps35.
Extended Data Fig. 4 Crystal structure of C. thermophilum Vps29, and comparison of cryo-ET structures with previous crystal structures.
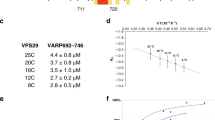
a, Crystal structure of C. thermophilum Vps29 (red) overlaid with the crystal structure of human VPS29 (blue)18. Crystallographic structure determination statistics are given in Extended Data Table 2. b, The fitted Vps26 dimer model with monomers coloured in dark green and light green. The homodimeric interface is formed by β-sheet extension of two N-terminal β-sandwich domains. The positions of the docked Vps26 models suggest the formation of an extended hydrophobic core between subunits. Close-up images of fitted Vps26 subunits highlight the extended hydrophobic core. c, Surface representation of the Vps26–Vps35–Vps29 trimer mapped with binding regions for retromer effectors. Neighbouring Vps5 and retromer proteins in the assembled array are shown as ribbons. Retromer components are coloured as in Fig. 1. The bottom boxes show higher-magnification views of the indicated regions of the overviews. Binding sites observed in structural data are coloured according to the colour of the corresponding label; dashed lines indicate binding regions identified in biochemical assays. The binding interfaces of human SNX27, SNX3, SNX3–DMT1-II, VARP–TBC1D5 were modelled using coordinates with PDB accession numbers 4P2A, 5F0L and 5GTU, respectively; the DMT1-II cargo peptide is shown as a ribbon. The SNX3–DMT1-II binding site overlaps with that of Vps5. The binding site of the SNX27 PDZ domain on VPS2610 is accessible, although owing to a lack of structural information regarding full-length SNX27 it is unclear whether this binding is consistent with simultaneous membrane binding by the SNX27 PX domains. The regulatory factors VARP and TBC1D5 share a binding interface on VPS29 that is exposed towards the outer extremity of the coat. This site in human VPS29 is also hijacked by the RidL protein from the pathogen Legionella pneumophila54,55. However, as for SNX27, full-length structures of VARP, TBC1D5 and RidL are not available so we cannot be sure how they will be arranged in the fully assembled array. Rab7 has been speculated to contribute to membrane recruitment of retromer by binding to VPS35 in the region indicated by the dashed line56,57. The deletion of this helical region (helix 6 in Saccharomyces cerevisiae) resulted in loss of interaction with Ypt757 (the Rab7 homologue in yeast). It has recently been shown that retromer binding to the PX-BAR complex displaces Rab7 during the formation of tubules58,59. The VPS10 ‘binding site’ (dashed line) indicates a region where point mutations affect VPS10 recycling16; however, no biochemical interactions between Vps10 and retromer have been shown, and our efforts at detecting a physical interaction between Vps35 and the cytosolic domains of Vps10 have not shown any direct binding. d, SNX3 and DMT1-II (transparent yellow and dark green surfaces respectively), as bound to VPS35–VPS26 from PDB 5F0L17, overlaid with our retromer–Vps5 complex structure (ribbons, coloured as above) demonstrating a steric clash between Vps5 BAR and SNX3 PX domains. Note that the C-terminal helix of Vps5 BAR (arrowheads) clashes with the DMT1-II cargo peptide density. The left box shows the same view as that above in c; the right panel shows the model rotated by 90° around the vertical axis to provide the view along the long axis of the BAR domain.
Extended Data Fig. 5 Global rigid-body docking of Vps5, Vps26–Vps35(N) and Vps35(C)–Vps29, and adaptation of the retromer coat to different membrane curvatures.
a–c, Fitting of structures to electron density maps was performed from 10,000 random initial placements of atomic models using the ‘fit’ command in Chimera. a–c, The cross-correlation between the model and the electron microscopy map is plotted against the fraction of the structural model within the electron-microscopy density threshold for: the global fit of the Vps5 dimer into the membrane-associated BAR domain density under the arch (a) (map f3 in Extended Data Fig. 2); the global fit of Vps26–Vps35(N) into the base of the arch (b) (map f5 in Extended Data Fig. 2); the global fit of Vps29/Vps35(C) into the apex of the arch (c) (map f1 in Extended Data Fig. 2). Arrows indicate the high-scoring rigid-body fits that were used as starting points for flexible fitting. d, e, For a subset of about 50% of the data we calculated tube centroids by spline fitting, and determined local membrane-curvature as the inverse of the distance from the subtomogram to the tube centroid. d, Slices through averages of 20% of the subtomograms from the dataset with lowest (left) and highest (right) membrane curvature, focused on the arch (top) or Vps26 dimer (bottom). See also the animation in Supplementary Video 5. e, The distribution of membrane curvatures of retromer tubules in vivo and in vitro. The lumenal diameters of each tube were measured manually, from which the mean and standard deviation were calculated.
Extended Data Fig. 6 Comparison of yeast PX-BAR proteins Vps5 and Vps17.
a, Overlay of the Vps5 heterodimer model after flexible fitting into the cryo-ET structure (blue) with the human SNX9 PX-BAR domains (beige; the bound PI3P head group is also shown in magenta in stick representation). The PX and BAR domains in Vps5 adopt a very similar architecture to the SNX9 protein, but there are variations in the angle between the BAR domains, and in the orientations of the lateral PX domains. The second and third α-helices of the Vps5 BAR domain are also longer than those of SNX9. b, Sequence alignment of the C. thermophilum PX-BAR proteins CtVps5 and CtVps17 and the human PX-BAR proteins SNX1 and SNX5). The secondary structure of CtVps5 is indicated above the sequences. The sequence alignment and its representation were prepared in MultAlign60 and ESPript 3.061. c, Overlay of ribbon models of CtVps5 (blue) and CtVps17 (grey). The structure of CtVps17 was modelled using CtVps5 as a template (SWISS-MODEL).
Extended Data Fig. 7 Arrangement of the retromer coat on membrane tubules by cryo-ET.
a, b, 3D plots that visualize the relative positions of neighbouring Vps26 dimers (a) or neighbouring Vps35–Vps29 arches (b) (see Supplementary Information for details). The isosurface for visualization is set at 8σ. c, d, Flattened cylindrical projections through the boxed regions in volumes a, b, respectively. The blue (low) to red (high) gradient colouring is proportional to pixel values. The white circle shows the position of the central Vps26 dimer or arch. e, A close-up view of the boxed region in c with arrows indicating the position and the identity of neighbours corresponding to each of six nearest-neighbour relative arrangements between Vps26 dimers. f, Bar plot of the occurrence of arrangements from e for 15,795 analysed Vps26 dimers. The arrangements numbered 3 and 6, in which Vps26 dimers are very closely packed, are less frequent than other arrangements. Models of these relative arrangements are shown in Fig. 3b, c. g, Density maps are shown for the local retromer structure for each of the six different relative Vps26 arrangements. Numbering corresponds to arrangements shown in Fig. 3b. Maps are radially coloured in grey, blue, green and gold for the membrane, Vps5, Vps26 and Vps35–Vps29 layers, respectively. h, Overlay of density maps for arrangements 2 and 3, and 2 and 4, showing that in some arrangements, the Vps35 arch can tilt relative to the tubule to accommodate nearby arches. The average of arrangement 2 is coloured as above, whereas the averages of arrangement 3 and 4 are coloured transparent grey.
Supplementary information
Supplementary Figures
This file contains source images for gels.
Video 1
CryoET image of retromer-Vps5 coated tubules (example 1). Sequential image slices through an example tomogram from the total dataset of 71 tomograms of retromer-Vps5-coated membrane tubules. Scale bar – 100 nm. We note that the tips and residual donor liposomes are also coated. These have similar membrane curvatures at least in one direction to that of the sides of the tubules.
Video 2
CryoET image of retromer-Vps5 coated tubules (example 2). Sequential image slices through a second example tomogram from the total dataset of 71 tomograms of retromer-Vps5-coated membrane tubules. Scale bar – 100 nm.
Video 3
CryoET image of retromer-Vps5 coated tubules (example 3). Sequential image slices through a third example tomogram from the total dataset of 71 tomograms of retromer-Vps5-coated membrane tubules. Scale bar – 100 nm.
Video 4
Structure of the membrane-assembled retromer coat by cryo-electron tomography. A video summary of the structural data presented in this study.
Video 5
Retromer coat adapting to different membrane curvature. An animation of the data shown in Extended Data Fig. 5a.
Rights and permissions
About this article

Cite this article
Kovtun, O., Leneva, N., Bykov, Y.S. et al. Structure of the membrane-assembled retromer coat determined by cryo-electron tomography. Nature 561, 561–564 (2018). https://doi.org/10.1038/s41586-018-0526-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0526-z
Keywords
This article is cited by
-
Plasma FIB milling for the determination of structures in situ
Nature Communications (2023)
-
On the causal role of retromer-dependent endosomal recycling in Alzheimer’s disease
Nature Cell Biology (2023)
-
Architecture of the ESCPE-1 membrane coat
Nature Structural & Molecular Biology (2023)
-
Structural organization of the retriever–CCC endosomal recycling complex
Nature Structural & Molecular Biology (2023)
-
Cryo-electron tomography on focused ion beam lamellae transforms structural cell biology
Nature Methods (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}