
Abstract
Global surveillance has identified emerging SARS-CoV-2 variants of concern (VOC) associated with broadened host specificity, pathogenicity, and immune evasion to vaccine-induced immunity. Here we compared humoral and cellular responses against SARS-CoV-2 VOC in subjects immunized with the DNA vaccine, INO-4800. INO-4800 vaccination induced neutralizing antibodies against all variants tested, with reduced levels detected against B.1.351. IFNγ T cell responses were fully maintained against all variants tested.
Similar content being viewed by others

Main text
SARS-CoV-2, the causative agent of the COVID-19 pandemic, continues to cause unprecedented levels of mortality and socioeconomic burden. Concerningly, virus surveillance shows the global spread of novel SARS-CoV-2 variants, which are more infectious and display increased transmissibility and pathology1,2,3. Some of these variants of concern (VOC) contain mutations in the Spike protein receptor-binding domain (RBD), the region which interacts with the host ACE2 receptor, and to which many SARS-CoV-2 neutralizing antibodies target. The B.1.1.7 lineage, the first emerging VOC, contains the key N501Y and D614G mutations and the 69–70 deletion in the RBD and/or S1 regions and exhibits increased transmissibility, but does not appear to significantly evade neutralizing antibody responses generated by current vaccines approved for use4,5. The B.1.351 and P.1 lineages have additional mutations, including E484K in the RBD region6,7,8, as well as unique changes in position K417. Notably, sera isolated from convalescent individuals and vaccinees exposed to the original Wuhan Spike protein sequence have shown significantly lower levels of neutralizing activity against the B.1.351 and P.1 variants8,9,10,11,12.
INO-4800 is a SARS-CoV-2 Spike DNA-based vaccine that is delivered intradermally followed by electroporation (EP) using CELLECTRA® 2000 and is currently undergoing clinical development. In Phase 1 clinical trial, INO-4800 vaccination induced a balanced immune response characterized by both functional antibody and T cell responses in vaccinated subjects13. Both humoral and cellular immune responses have been shown to be important components of protection against betacoronaviruses14,15,16. In the present study, we have assessed the humoral and T cell responses against SARS-CoV-2 B.1.1.7, B.1.351, and P.1 variants elicited after INO-4800 vaccination (Supplementary Fig. 1A).
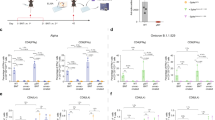
In INO-4800 vaccinated subjects, serum IgG antibody binding titers to SARS-CoV-2 full-length Spike proteins were evaluated by ELISA using proteins specific for B.1.1.7, B.1.351, and P.1 variants (Fig. 1a and Supplementary Fig. 1A). IgG binding titers were not negatively impacted between Wuhan and B.1.1.7 or B.1.351 variants. An average 1.9-fold reduction was observed for the P.1 variant in subjects tested at week 8 after receiving two doses of INO-4800 (Fig. 1a).

a Sera from Phase 1 INO-4800 vaccinees were assessed by ELISA for IgG binding to Wuhan, B.1.1.7, B.1.351, and P.1 variant Spike proteins (S1 and S2). Data points indicate endpoint titers for an individual study sample (n = 4, 0.5 mg vaccine dose; n = 5, 1.0 mg; n = 11, 2.0 mg) and were calculated as the titer that exhibited an OD 3 SD above baseline. b SARS-CoV-2 pseudovirus neutralization ID50 titers for sera samples from 13 Phase 1 INO-4800 vaccinees comparing Wuhan against B.1.1.7, B.1.351, and P.1. Each data point represents the mean of technical duplicates for each individual (n = 1, 0.5 mg vaccine dose; n = 4, 1.0 mg; n = 8, 2.0 mg). Dotted lines indicate the limit of detection of 16. ns not significant, *P < 0.05, ***P < 0.0001 (Wilcoxon signed-rank test).
We performed a SARS-CoV-2 pseudovirus neutralization assay using sera collected from thirteen subjects two weeks after administration of a third dose of 0.5, 1, or 2 mg of INO-4800 (Supplementary Table 1). Neutralizing activity was detected against Wuhan and the emerging variants in all serum samples tested (Fig. 1b). The mean ID50 titers for the Wuhan, B.1.1.7, B.1.351, and P.1. were 643, 295, 105, and 664, respectively (Supplementary Tables 1 and 2). Compared to Wuhan, there was a 2.1 and 6.9-fold reduction for B.1.1.7 and B.1.351, respectively, while there was no difference between Wuhan and the P.1 variant. These results are consistent with other recent studies, which have demonstrated a significant reduction in neutralizing activity in vaccinated individuals towards the B.1.351 (≥6-fold reduction), while the B.1.1.7 lineage has demonstrated a reduced activity of two-fold or less4,7,8,9,17. Strikingly, while the P.1 strain presents with similar RBD mutations as B.1.351, we did not observe any reduction in neutralizing activity compared to the Wuhan strain in INO-4800 vaccinated individuals18,19. The P.1 lineage has similar changes in the RBD to the B.1.351 lineage as they contain the N501Y mutation found in the B.1.1.7 lineage and identical E484K mutations. However, they have similar but different mutations at position K417, with a change to T for the P.1 and to N for the B.1.351 lineage. While both are changes to polar uncharged side chains, the B.1.351 N mutation carries an additional amino group. It is possible that this position and this change allow for the impact of neutralizing responses observed. In addition, since P.1 and B.1.351 have almost identical RBD mutations, changes outside the RBD may play a role in neutralization, especially in the N-terminal domain (NTD), where a large number of mutations have accumulated in the P.1 and B.1.351 variants. However, B.1.351 variant contains unique deletions in the NTD region that is a known epitope for potent neutralizing antibodies19. The P.1 variant, while having similar RBD changes relative to B.1.351, it does not have said deletions in the NTD.
Recent reports have supported an important role for T cell immunity in protecting against COVID-19 in the absence of antibodies to SARS-CoV-220,21. Through optimized DNA construct design, combined with intradermal enhanced delivery, Inovio’s DNA platform technology has been demonstrated to drive balanced humoral and cellular immune responses to a wide range of infectious disease and tumor antigen targets22,23,24. We, therefore, compared cellular immune responses to Wuhan and SARS-CoV-2 Spike variants elicited by INO-4800 vaccination. Peripheral blood mononuclear cells (PBMCs) isolated from ten subjects at week 8 after receiving their second dose of INO-4800 were stimulated with Wuhan, B.1.1.7, B.1.351, or P.1 Spike peptides and cellular responses were measured by IFNγ ELISpot assay. We observed strikingly similar cellular responses to Wuhan (median = 82.2 IFNγ spot-forming units [SFUs]/106 PBMCs, IQR = 58.9−205.3), B.1.1.7 (median = 79.4, IQR = 38.9−179.7), B.1.351 (median = 80.0, IQR = 40.0−208.6) and P.1 (median = 78.3, IQR = 53.1−177.8) Spike peptides (Fig. 2). This is consistent with published results showing that, compared with neutralizing antibody responses, cellular immunity is relatively unimpaired by current VOCs 25. Here, we show that T cell responses are consistently maintained between Wuhan and all SARS-CoV-2 variants tested, including B.1.351 and P.1. Cells stimulated with peptides against these variants generated IFNγ responses as well as cytokines associated with CD8+ cytotoxic T cell responses (data not shown).

PBMCs from 10 Phase 1 subjects were collected 8 weeks after receiving the second dose of INO-4800 (n = 5, 1.0 mg; n = 5, 2.0 mg). PBMCs were treated with peptide pools spanning the entire Spike proteins of the Wuhan, B.1.1.7, B.1.351, or P.1 variants, and cellular responses were measured by IFNγ ELISpot assay. Mean ± s.e.m. IFNγ SFUs/million PBMCs of experimental triplicates are shown. ns not significant (Wilcoxon signed-rank test).
There is a growing concern regarding the protective efficacy of vaccines recently approved for emergency use and those vaccines currently in development against the VOC. Recent studies have shown a dramatic reduction in serum neutralization levels against B.1.351, a 2 to 4-fold reduction against P.14,7,18, and a minimal impact against B.1.1.717. In addition, vaccine trials have shown a considerable reduction in protective efficacy against B.1.35111,12,26. Interestingly, a small reduction in protective efficacy is observed in Latin American countries including Brazil, suggesting that P.1 may have emerged around the time of clinical trials were in effect26. Here we report the neutralizing antibody and T cell activity measured in INO-4800 vaccinated subjects against emerging SARS-CoV-2 variants first detected in the United Kingdom, South Africa, and Brazil. The neutralization levels against B.1.351 and B.1.1.7 for the INO-4800 SARS-CoV-2 Spike DNA vaccine are consistent with previous reports of subjects receiving vaccines encoding for the ancestral Spike protein7,27. We also showed that INO-4800 generates robust neutralizing antibodies against P.1 at levels comparable to the Wuhan strain. Despite similarities in the RBD of P.1 and B.1.351, studies have reported a reduced impact in neutralization against P.1, compared to B.1.35118,19. In sera from individuals who received the SARS-CoV-2 mRNA-1273 or the BNT162b2 vaccines, there was a 2.8 and 2.2-fold reduction in neutralization against P.1, compared to 8.6 and 6.5-fold for B.1.351, respectively18. The lower resistance to neutralization conferred by the P.1 variant suggests that changes outside the RBD, especially in the NTD, where P.1 and B.1.351 acquired and accumulated different mutations, may play a role in neutralization. INO-4800 induces cross-reactive T cell responses against B.1.1.7, B.1.351, and P.1 variants that are comparable to the Wuhan strain. Taken together, these data demonstrate maintenance of one or both cellular and humoral arms of the immune response against emerging SARS-CoV-2 variants for the INO-4800 vaccine, which will likely be critical factors to impact the ongoing COVID-19 pandemic.
Methods
Clinical trial subject samples
Serum and PBMC samples were acquired from participants of the phase I INO-4800 clinical trial (NCT04336410) described previously13. The trial has since been expanded to include participants of 51−64 and 64+ years of age as separate groups in addition to the original 18–50 age group. A 0.5 mg dose group was also added. The institutional review board of each clinical site approved the trial. All trial participants provided written informed consent. Sera from 20 subjects out of the 120 total study participants were selected for analysis on variant Spike protein binding ELISAs and variant pseudovirus neutralization assays. The samples analyzed by pseudovirus neutralization assay were collected from subjects two weeks after a third dose of INO-4800, and the samples used for other ELISA and ELISpot were collected after two doses.
Antigen binding ELISA
Binding ELISAs were performed as described previously13, except different variants of SARS-CoV-2 S1+S2 proteins were used for plate coating. The S1+S2 wild-type Spike protein (Acro Biosystems #SPN-C52H8) contained amino acids 16-1213 of the full Spike protein (Accession #QHD43416.1) with R683A and R685A mutations to eliminate the furin cleavage site. The B.1.1.7, B.1.351, and P.1 S1+S2 variant proteins (Acro Biosystems #SPN-C52Hc,#SPN-C52H6, and #SPN-C52Hg, respectively) additionally contained the following proline substitutions for trimeric protein stabilization: F817P, A892P, A899P, A942P, K986P, and V987P. The B.1.1.7 protein contained the following variant-specific amino acid substitutions: HV69-70del, Y144del, N501Y, A570D, D614G, P681H, T716I, S982A, and D1118H; the B.1.351 protein contained the following substitutions: L18F, D80A, D215G, R246I, K417N, E484K, N501Y, D614G, and A701V; and the P.1 protein contained the following: L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, D614G, H655Y, T1027I, and V1176F. Assay plates were coated using 100 µL of 2 µg/mL of protein.
SARS-CoV-2 pseudovirus production
SARS-CoV-2 pseudovirus stocks encoding for the Wuhan, B.1.1.7, B.1.351, or P.1 Spike protein was produced using HEK 293T cells transfected with Lipofectamine 3000 (ThermoFisher) using IgE-SARS-CoV-2 S plasmid variants (Genscript) co-transfected with pNL4-3.Luc.R-E- plasmid (NIH AIDS reagent) at a 1:8 ratio. Seventy-two hours post transfection, supernatants were collected, steri-filtered (Millipore Sigma), and aliquoted for storage at −80 °C.
SARS-CoV-2 pseudoviral neutralization assay
CHO cells stably expressing ACE2 (ACE2-CHOs) were used as target cells plated at 10,000 cells/well. SARS-CoV-2 pseudovirus was tittered to yield greater than 30 times the cells only control relative luminescence units (RLU) after 72 h of infection. Sera from 13 INO-4800 vaccinated subjects were heat-inactivated and serially diluted two-fold starting at 1:16 dilution. Sera were incubated with SARS-CoV-2 pseudovirus for 90 min at room temperature. After incubation, sera-pseudovirus mixture was added to ACE2-CHOs and allowed to incubate in a standard incubator (37% humidity, 5% CO2) for 72 h. After 72 h, cells were lysed using Bright-Glo™ Luciferase Assay (Promega) and RLU was measured using an automated luminometer. Neutralization titers (ID50) were calculated using GraphPad Prism 8 and defined as the reciprocal serum dilution at which RLU were reduced by 50% compared to RLU in virus control wells after subtraction of background RLU in cell control wells.
SARS-CoV-2 Spike ELISpot assay
Peripheral mononuclear cells (PBMCs) were stimulated in vitro with 15-mer peptides (overlapping by 11 amino acids) spanning the full-length Spike protein sequence of the indicated variants. Variant peptide pools (JPT PepmixTM) included the following changes to match published deletions/mutation in each variant: B.1.1.7 variant (delta69-70, delta144, N501Y, A570D, D614G, P681H, T716I, S982A, and D1118H), B.1.351 variant (L18F, D80A, D215G, delta242-244, R246I, K417N, E484K, N501Y, D614G, A701V); P.1 variant L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, D614G, H655Y, T1027I, and V1176F). Cells were incubated overnight with peptide pools at a concentration of 1 μg per ml per peptide in a precoated ELISpot plate, (MabTech, Human IFNγ ELISpot Plus). Cells were then washed off, and the plates were developed via a biotinylated anti-IFN-γ detection antibody followed by a streptavidin-enzyme conjugate resulting in visible spots. After plates were developed, spots were scanned and quantified using the CTL S6 Micro Analyzer (CTL) with ImmunoCapture and ImmunoSpot software. Values are shown as the background-subtracted average of measured triplicates. The ELISpot assay qualification determined that 12 spot forming units were the lower limit of detection. Thus, anything above this cutoff signal is an antigen-specific cellular response.
Statistical methods
GraphPad Prism 8.1.2 (GraphPad Software, San Diego, USA) was used for graphical and statistical analysis of data sets. P values of <0.05 were considered statistically significant. A nonparametric two-tailed student t-test Wilcoxon signed-rank test was used to assess statistical significance in Figs. 1 and 2.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
References
Chen, R. E. et al. Resistance of SARS-CoV-2 variants to neutralization by monoclonal and serum-derived polyclonal antibodies. Nat. Med. https://doi.org/10.1038/s41591-021-01294-w (2021).
Davies, N. G. et al. Increased mortality in community-tested cases of SARS-CoV-2 lineage B.1.1.7. Nature https://doi.org/10.1038/s41586-021-03426-1 (2021).
Davies, N. G. et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science https://doi.org/10.1126/science.abg3055 (2021).
Wu, K. et al. Serum Neutralizing Activity Elicited by mRNA-1273 Vaccine. N. Engl. J. Med. https://doi.org/10.1056/NEJMc2102179 (2021).
Xie, X. et al. Neutralization of SARS-CoV-2 spike 69/70 deletion, E484K and N501Y variants by BNT162b2 vaccine-elicited sera. Nat. Med. https://doi.org/10.1038/s41591-021-01270-4 (2021).
Wibmer, C. K. et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. https://doi.org/10.1038/s41591-021-01285-x (2021).
Wang, Z. et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature https://doi.org/10.1038/s41586-021-03324-6 (2021).
Garcia-Beltran, W. F. et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell https://doi.org/10.1016/j.cell.2021.03.013 (2021).
Wang, P. et al. Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature https://doi.org/10.1038/s41586-021-03398-2 (2021).
Edara, V. V. et al. Reduced binding and neutralization of infection- and vaccine-induced antibodies to the B.1.351 (South African) SARS-CoV-2 variant. Preprint at bioRxiv https://doi.org/10.1101/2021.02.20.432046 (2021).
Madhi, S. A. et al. Efficacy of the ChAdOx1 nCoV-19 Covid-19 vaccine against the B.1.351 variant. N. Engl. J. Med. https://doi.org/10.1056/NEJMoa2102214 (2021).
Mahase, E. Covid-19: Novavax vaccine efficacy is 86% against UK variant and 60% against South African variant. J. BMJ 372, n296 (2021).
Tebas, P. et al. Safety and immunogenicity of INO-4800 DNA vaccine against SARS-CoV-2: a preliminary report of an open-label, Phase 1 clinical trial. EClinicalMedicine 31, 100689 (2021).
Channappanavar, R., Zhao, J. & Perlman, S. T cell-mediated immune response to respiratory coronaviruses. Immunologic Res. 59, 118–128 (2014).
Sariol, A. & Perlman, S. Lessons for COVID-19 immunity from other coronavirus infections. Immunity 53, 248–263 (2020).
McMahan, K. et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 590, 630–634 (2021).
Planas, D. et al. Sensitivity of infectious SARS-CoV-2 B.1.1.7 and B.1.351 variants to neutralizing antibodies. Nat. Med. https://doi.org/10.1038/s41591-021-01318-5 (2021).
Wang, P. et al. Increased resistance of SARS-CoV-2 variant P.1 to antibody neutralization. Cell Host Microbe. https://doi.org/10.1016/j.chom.2021.04.007 (2021).
Dejnirattisai, W. et al. Antibody evasion by the P.1 strain of SARS-CoV-2. Cell https://doi.org/10.1016/j.cell.2021.03.055 (2021).
Sekine, T. et al. Robust T cell immunity in convalescent individuals with asymptomatic or mild COVID-19. Cell 183, 158–168.e114 (2020).
Gallais, F. et al. Intrafamilial exposure to SARS-CoV-2 associated with cellular immune response without Seroconversion, France. Emerg. Infect. Dis. 27, 113–121 (2021).
Smith, T. R. F. et al. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat. Commun. 11, 2601 (2020).
Edupuganti, S. et al. Intramuscular and intradermal electroporation of HIV-1 PENNVAX-GP(®) DNA vaccine and IL-12 Is safe, tolerable, acceptable in healthy adults. Vaccines https://doi.org/10.3390/vaccines8040741 (2020).
Tebas, P. et al. Intradermal SynCon® Ebola GP DNA vaccine is temperature stable and safely demonstrates cellular and humoral immunogenicity advantages in healthy volunteers. J. Infect. Dis. 220, 400–410 (2019).
Tarke, A. et al. Impact of SARS-CoV-2 variants on the total CD4+ and CD8+ T cell reactivity in infected or vaccinated individuals. Cell Rep. https://doi.org/10.1016/j.xcrm.2021.100355 (2021).
Johnson, J. Johnson & Johnson Announces Single-Shot Janssen COVID-19 Vaccine Candidate Met Primary Endpoints in Interim Analysis of its Phase 3 ENSEMBLE Trial. https://www.jnj.com/johnson-and-johnson-announces-single-shot-janssen-covid-19-vaccine-candidate-met-primary-endpoints-in-interim-analysis-of-its-phase-3-ensemble-trial (2021).
Stephenson, K. E. et al. Immunogenicity of the Ad26.COV2.S vaccine for COVID-19. Jama https://doi.org/10.1001/jama.2021.3645 (2021).
Acknowledgements
We acknowledge the members of the Inovio Pharmaceuticals R&D department for significant technical assistance. This work is funded by Coalition for Epidemic Preparedness Innovations (CEPI).
Author information
Authors and Affiliations
Contributions
Conceptualization, V.M.A., S.J.R., J.D.B., K.E.B., T.R.F.S., J.J.K., and L.M.H.; Methodology, V.M.A., A.C.Q., I.M., J.A., C.R., D.E., K.S., and T.M.; Investigation, V.M.A., A.C.Q., I.M., J.A., C.R., D.E., K.S., T.M., and P.P.; Resources, M.P., E.R., P.T., P.P., and D.B.W.; Writing-original draft preparation, V.M.A., J.T., A.C.Q., I.M., J.A., C.R., D.E., K.S., and T.M.; Writing-review and editing, V.M.A., J.T., A.C.Q., I.M., J.A., C.R., D.E., K.S., and T.M; Project administration, S.J.R., J.D.B., K.E.B., T.R.F.S., J.J.K., and L.M.H.
Corresponding author
Ethics declarations
Competing interests
T.R.F.S., J.J.K., D.E., S.J.R., A.CQ., I.M., J.A., C.R., J.T., T.M., K.S., P.P., T.H., V.M.A., J.D.B., L.M.H., and K.E.B. are employees of Inovio Pharmaceuticals and as such receive salary and benefits, including ownership of stock and stock options, from the company. D.B.W. has received grant funding, participates in industry collaborations, has received speaking honoraria, and has received fees for consulting, including serving on scientific review committees and board services. Remuneration received by D.B.W. includes direct payments or stock or stock options, and in the interest of disclosure, he notes potential conflicts associated with this work with Inovio and possibly others. In addition, he has a patent DNA vaccine delivery pending to Inovio. All other authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Andrade, V.M., Christensen-Quick, A., Agnes, J. et al. INO-4800 DNA vaccine induces neutralizing antibodies and T cell activity against global SARS-CoV-2 variants. npj Vaccines 6, 121 (2021). https://doi.org/10.1038/s41541-021-00384-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41541-021-00384-7
This article is cited by
-
Genetic fusion of CCL11 to antigens enhances antigenicity in nucleic acid vaccines and eradicates tumor mass through optimizing T-cell response
Molecular Cancer (2024)
-
Nanoparticle-based DNA vaccine protects against SARS-CoV-2 variants in female preclinical models
Nature Communications (2024)
-
Safety and immune response kinetics of GRAd-COV2 vaccine: phase 1 clinical trial results
npj Vaccines (2022)
-
Advances in the design and development of SARS-CoV-2 vaccines
Military Medical Research (2021)
