
Abstract
Heterosis utilization in a large proportion of crops depends on the use of cytoplasmic male sterility (CMS) tools, requiring the development of homozygous fertile lines and CMS lines1. Although doubled haploid (DH) technology has been developed for several crops to rapidly generate fertile lines2,3, CMS lines are generally created by multiple rounds of backcrossing, which is time consuming and expensive4. Here we describe a method for generating both homozygous fertile and CMS lines through in vivo paternal haploid induction (HI). We generated in-frame deletion and restored frameshift mutants of BoCENH3 in Brassica oleracea using the CRISPR/Cas9 system. The mutants induced paternal haploids by outcrossing. We subsequently generated HI lines with CMS cytoplasm, which enabled the generation of homozygous CMS lines in one step. The BoCENH3-based HI system provides a new DH technology to accelerate breeding in Brassica and other crops.
Similar content being viewed by others


Main
Heterosis utilization has been extensively applied in plant breeding and contributes greatly to crop yield5. One of the most important processes for heterosis utilization is to develop homozygous (true-breeding) lines, which requires 6–8 generations of selfing2,6. In addition, heterosis utilization in many crops including most Brassica crops, as well as millet, carrot and some rice, maize, soybean and sorghum crops, exploits cytoplasmic male sterility (CMS) as a pollination control tool for hybrid seed production, requiring another 6–8 rounds of backcrossing to generate homozygous CMS lines4. These selfing and backcrossing processes are time consuming and costly for most crops7.
Doubled haploid (DH) technology has broad application in generating true-breeding lines within 1–2 generations8. In vitro haploid induction (HI) methods such as anther culture and microspore embryogenesis are expensive and limited by genotype recalcitrance9. In vivo HI technology has commonly been used in maize breeding for several decades10. In recent years, due to the cloning of genes including MTL/NLD/ZmPLA1, DMP, ZmPOD65, CENH3 and ECS1/2 that are responsible for HI3,11,12,13, in vivo HI has been extended to several plant species including Arabidopsis, rice, wheat, tomato, rapeseed, cabbage and Medicago truncatula2,7,8,9,14,15,16.
The Arabidopsis CENTROMERIC HISTONE3 (CENH3) gene encodes a centromere-identifying protein histone H3 variant17. Modification of this gene has been shown to induce mainly paternal haploid progeny upon outcrossing to the wild type (WT)12,18. However, the establishment of a CENH3-based system is relatively difficult because it requires modifications such as the use of GFP–tailswap chimaeric proteins, the expression of non-native CENH3, in-frame deletions, single amino acid substitutions and heterozygous cenh3 null mutation (+/cenh3), while homozygous cenh3 null mutation is lethal8,19,20,21,22. At present, although CENH3 is conserved in both monocotyledonous and dicotyledonous plants, CENH3-based paternal HI is available only for Arabidopsis, maize and wheat8,21,22.
The paternal haploid inducer line has the advantage of transferring cytoplasm between different genotypes, as the cytoplasm is maternally inherited during crossing. The maize indeterminate gametophyte1 (ig1) mutation induces the formation of androgenetic haploids with an HI rate of 1–3% (refs. 23,24). This ig1 haploid inducer is routinely used by breeders to transfer CMS cytoplasms, such as C, S, SD, Vg, ME and CA, to inbred lines23. The CENH3-based haploid inducer also enables the transfer of cytoplasm among different ecotypes in Arabidopsis8,25, but it has not been employed to produce CMS lines.
We tested whether modification of the CENH3 homologue can be used for paternal HI and whether a CENH3-based paternal inducer line with a CMS cytoplasm enables the creation of CMS lines through the exchange of cytoplasmic and nuclear genomes in broccoli (Brassica oleracea var. italica), a globally important vegetable crop.
Using the Arabidopsis CENH3 as query, we searched the broccoli HDEM genome with the Basic Local Alignment Search Tool for protein (BLASTP) and found only one CENH3 homologue BolC8t52879H (BoCENH3). Phylogenetic analysis revealed that BoCENH3 is highly similar to Arabidopsis CENH3 with 66% sequence identity, and they are assigned to the same subclade (Supplementary Fig. 1 and Supplementary Table 1). We cloned BoCENH3 from the broccoli inbred line CX33 and detected its expression in various broccoli tissues by reverse transcription quantitative PCR(RT‒qPCR), which indicated that BoCENH3 is highly expressed in pistils and young buds, especially in pistils at 24–72 h after pollination (Supplementary Fig. 2), consistent with what has been found for authentic CENH3 genes in Arabidopsis, barley and wheat19,21.
We employed a CRISPR/Cas9 system to create hypomorphic alleles of BoCENH3. A CRISPR/Cas9 construct was generated with two specific guide RNA sequences (sgRNA1 and sgRNA2) targeting the 7th exon of BoCENH3, located within the sequence encoding the putative α2-helix region26 (Fig. 1a,b). The construct was subsequently introduced into the inbred line CX33, after which 21 independent transgenic lines were obtained. Among these transgenic lines, 11 lines harboured mutations in either of the targeted regions (3 for both sgRNA1 and sgRNA2, and 8 for only sgRNA2), corresponding to editing efficiencies of 14.29% for sgRNA1 and 52.38% for sgRNA2 (Supplementary Table 2). Eight lines were heterozygous or chimaeric mutants with deletions/insertions that led to frameshift and premature termination. Although not homozygous mutants, they displayed wrinkled leaves, a typical feature of defects in cell division (Supplementary Fig. 3). No homozygous mutants were produced from the T0 genome editing lines or selfing of T0 lines, consistent with the previous reports that CENH3 is essential for plant development and reproduction12,26. Two representative heterozygous (BoCENH3/bocenh3) mutants were crossed by pollen donors, the broccoli lines B54 and 22TZ, and 2,512 progenies from four crosses were assessed (Supplementary Table 2). Although the heterozygous CENH3 mutant (+/cenh3) with a null allele triggered haploid production in maize22, the broccoli heterozygous (BoCENH3/bocenh3) mutants did not induce haploids at the scale tested here (zero haploid in a screen of ~600 plants, for an average HI frequency of <0.16). The HI rate in lines homozygous for WT CENH3 is probably considerably lower than this upper limit, as in a test for spontaneous paternal haploid in the related species A. thaliana, paternal haploids were not observed in a population of over 800,000 F1 progenies (<1 × 10−6)27. The remaining two lines, #3 and #8, harboured in-frame deletions in targeted region 2 and a restored frameshift (RFS) between the two targeted regions, respectively (Fig. 1a,b and Supplementary Table 2). From the T1 generation, we generated non-transgenic homozygous mutants BoCENH3Δ3 (BoCENH3Δ3/BoCENH3Δ3) and BoCENH3RFS33 (BoCENH3RFS33/BoCENH3RFS33), which exhibited a normal appearance compared with that of WT plants. When selfing or crossing as a female parent, the BoCENH3Δ3 and BoCENH3RFS33 mutants showed significantly reduced seed setting rates (P < 0.01; unpaired, two-tailed t-test) (Fig. 1c,d).

a, Schematic diagram of BoCENH3 gene structure and CRISPR/Cas9-based genome editing. Black blocks, gene coding region; red lines, sgRNA-targeted regions (T1, T2); underlining indicates the target sequence; protospacer adjacent motif sequences are highlighted in blue; insertions/deletions are highlighted in red. b, Amino acid alignment of BoCENH3, BoCENH3Δ3 and BoCENH3RFS33. Putative CATD and α-helix are indicated by black lines. Amino acid changes in BoCENH3Δ3 and BoCENH3RFS33 are outlined in black boxes. c, Representative siliques from selfed CX33 (WT) and BoCENH3 mutants. d, Per silique seed setting performance of selfed WT, BoCENH3Δ3 and BoCENH3RFS33. Means ± s.d. (n = 30) (unpaired, two-tailed t-test). e, Haploid genotyping with DNA marker. M, DNA size marker; I–II, PCR bands of the B54 inbred line and the BoCENH3Δ3 mutant; III–IV, F1 hybrids from BoCENH3Δ3 × B54; V–VII, three haploids from B54. f, Flow cytometry analysis of diploid B54 and haploid B54 generated by BoCENH3Δ3 inducer. g, Chromosome numbers in diploid B54 (n = 18) and haploid B54 (n = 9) plants. h, Phenotypes of diploid, haploid and aneuploid B54 seedlings. i–l, Phenotypes of diploid and haploid B54 adult plants (i), buds (j), flowers (k) and dissected flower tissues (l). m, Haploid induction rates of BoCENH3Δ3 and BoCENH3RFS33. Scale bars, 1 cm (c), 10 µm (g), 5 cm (h), 10 cm (i), 0.5 cm (j,k) and 1 cm (l).
To test whether the BoCENH3Δ3 and BoCENH3RFS33 mutants could induce haploids, these mutants were crossed by 3 pollen donors: B54, B188 and 22TZ. These pollen donors were broccoli inbred lines with distinct genetic backgrounds and phenotypes (Supplementary Fig. 4). We developed 9 molecular markers (one for each chromosome) showing insertion–deletion (indel) polymorphisms between the mutants and the male parents to screen all the progenies. Potential haploids that exhibited genotypes identical to those of the male parents were identified (Fig. 1e and Supplementary Fig. 5). Flow cytometry analyses and chromosome spreads confirmed that all the plants were true haploids (Fig. 1f,g). These haploids were morphologically similar to the corresponding male parents but had smaller plant sizes, thinner leaves and male sterility (Fig. 1h–l). The HI rate was 0.52% on average for BoCENH3Δ3 and 1.14% for BoCENH3RFS33, substantially higher than that observed in selfed WT plants (0 in 647 progenies, HI rate < 0.15%), WT × B54 crosses (0 in 574 progenies, HI rate < 0.17%) and WT × 22TZ crosses (0 in 709 progenies, HI rate < 0.14%) (Fig. 1m). Nevertheless, this HI rate is relatively lower than the previously reported 1–44% in Arabidopsis (varied in different CENH3 mutants)26, ~7% in wheat (in-frame modification of the N-terminal domain)21 and 5% in maize (heterozygous mutant with a cenh3 null allele)22. The generation of additional types of BoCENH3 mutants may be helpful in increasing the HI rate in broccoli. In addition, we identified aneuploids from the outcrossing progenies (Fig. 1h,m and Supplementary Fig. 6). Given that some CENH3 mutants could induce both paternal maternal and maternal haploids26, we tested whether the BoCENH3 mutants could trigger maternal haploids in reciprocal crosses. Although the seed setting rate was also slightly reduced when these two mutants were crossed as males, no haploids were identified at the scale (2,699 individuals) tested here (Supplementary Table 4).
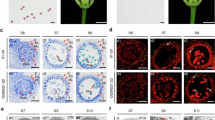
Almost all commercial B. oleracea hybrids are produced on the basis of the Ogura CMS system, which requires the creation of CMS lines28. Currently, CMS lines in Brassica crops are exclusively developed through the traditional backcrossing procedure28,29. Although swapping of CMS via a paternal HI line (ig1) is already used in maize23, this strategy has not been extended to other species. We tested whether the CMS cytoplasm can be exchanged to inbred lines via BoCENH3-based HI. To create HI lines in the Ogura CMS cytoplasm, the CMS line CMS219 was crossed with BoCENH3RFS33 pollen and then backcrossed to BoCENH3RFS33. From the BC1 generation, we identified homozygous BoCENH3RFS33 individuals with Ogura CMS cytoplasm (harbouring the orf138 gene)28. The HI-CMS line was crossed with a B54 pollen donor. We identified 5 haploids from 492 progenies (1.02%) using molecular markers, flow cytometry and plant phenotype analyses (Fig. 2a and Supplementary Fig. 7). Using the orf138 specific gene marker and nuclear genomic markers, we confirmed that these haploids had a B54 genetic background and Ogura CMS cytoplasm (Fig. 2b,c). These CMS B54 plants were treated with 200 mg l−1 colchicine solution for chromosome doubling. We successfully obtained two diploid CMS-B54 lines, which were morphologically similar to its maintainer B54 and showed typical Ogura male sterile traits (Fig. 2a,d and Supplementary Fig. 8). When crossed to its maintainer line, CMS-B54 exhibited normal seed setting performance (Fig. 2e and Supplementary Fig. 9). To further confirm the genomic background of the haploids and diploids, the parental lines, two diploid CMS B54 plants and six B54 haploids were subjected to whole-genome resequencing. More than 318,000 single-nucleotide polymorphisms (SNPs) were identified from the samples. Most SNPs (>99.5%) from B54 CMS lines and B54 haploids showed genotypes identical to the original B54 line and no credible maternally derived SNPs were found (Supplementary Table 5), indicating that BoCENH3RFS33 could induce clean paternal haploids.

a, Phenotypes of WT B54 and doubled haploid CMS B54 seedlings. White arrows indicate haploid leaves before diploidization with colchicine and black arrows indicate diploid leaves after treatment with colchicine. b, PCR confirmation of the cytoplasm type using an orf138 specific marker. M, DNA size marker; I, Ogura CMS line; II, inbred line B54; III–IV, B54 haploids with Ogura CMS cytoplasm. Three independent experiments were performed with similar results. c, Genetic background of the HI-CMS line, B54, F1 hybrid and haploid revealed by DNA markers on 9 chromosomes. d,e, Phenotype (d) and seed setting performance (e) of the CMS-B54 line. f, A proposed model of an effective CMS-based heterosis utilization system. Scale bars, 5 cm (a), 10 cm (whole plant in d), 5 cm (single flower in d) and 5 cm (e).
In summary, we successfully established a CENH3-based paternal HI system in broccoli, providing a novel and cost-effective DH technology without genotype recalcitrance. Importantly, by creating paternal haploids with maternal cytoplasm, homozygous CMS lines can be created in one step, which is a breakthrough for crops using CMS systems for hybrid seed production (a proposed system is shown in Fig. 2f). The success of this HI system could accelerate the improvement of B. oleracea cultivars and pave the way for extending CENH3-based HI to other crops.
Methods
Plant materials
The broccoli inbred line CX33 was employed for the purpose of genetic transformation and BoCENH3 knockout. The inbred lines B54, B188 and the 22TZ were used as test lines for HI of BoCENH3 inducers. The broccoli CMS line CMS219 was used as a donor of Ogura cytoplasm. All plants and the derived transgenic plants were grown under local conditions in Beijing, China. All selfing and cross-pollinations were performed in a greenhouse.
RNA isolation and RT‒qPCR analyses
Fresh broccoli tissues, including leaves, stems, roots, floral buds, petals, sepals, mature pollens, pistils and siliques were sampled and submerged in liquid nitrogen. An RNA extraction kit (Tiangen Biotech, 4992239) was used for total RNA isolation from these broccoli tissues. A complementary (c)DNA Synthesis kit (Tiangen, 4992910) was used for synthesizing cDNA following standard protocol. For qPCR assays, we prepared 25 μl PCR mixture using SYBR Green master mix (Takara, RR82WR) and performed PCR experiments on an RT–qPCR detection platform (CFX96 Touch, Bio-Rad). As an internal control, RT‒qPCR for the Actin gene of B. oleracea was also performed alongside the experimental group. Each assay was biologically repeated at least three times. The data analysis was performed using the 2−ΔΔCT method30.
Phylogenetic analyses
The protein sequence of BoCENH3 was downloaded from the broccoli draft genome HDEM (https://www.genoscope.cns.fr/projet_BKL/cgi-bin/gbrowse/boleracea/)31. To search for proteins homologous to this sequence, we utilized the BLASTP tool in Ensembl Plants (http://plants.ensembl.org/index.html) with BoCENH3 protein sequence as a query. A total of 12 BoCENH3 homologue proteins from 10 species were used for phylogenetic analyses. The protein sequences were subjected to alignment through ClustalW in MEGA software (v.7) and then a neighbour-joining phylogenetic tree was constructed using MEGA (Poisson model, 1,000 bootstrap replicates). Sequences of the homologous proteins used in this study are shown in Supplementary Table 1.
Genetic transformation and CRISPR/Cas9-mediated gene editing in broccoli
The sequence of BoCENH3 from broccoli CX33 was used for sgRNA design. We searched for the conserved motifs of α-helix and centromere-targeting domain (CATD) in BoCENH3 by comparing it to the known Arabidopsis CENH3 (ref. 26). Two sgRNA sequences targeting the putative α2-helix region were chosen and inserted into a modified vector downstream of the Arabidopsis U6 promoter. We further cloned and inserted this cassette to a vector with a CaMV 35S promoter-derived Bar selection marker and a CaMV 35S promoter-derived Cas9 gene as previously described32. The protocol for Agrobacterium-mediated broccoli transformation using the material CX33 has been described previously32,33. Positive transgenic plants were screened by Basta resistance and Bar gene specific marker.
The genomic fragment encompassing the sgRNA-targeted regions was amplified from positive transgenic plants and subjected to Sanger sequencing. To determine the mutations in T0 plants, PCR amplicons from these lines with expected mutations in BoCENH3 were inserted into a cloning vector, introduced into Escherichia coli strain EH5α and then sequenced (at least 20 independent clones for each line)32. Homozygous in-frame deletion and restored frameshift mutants of BoCENH3 in the T1 or BC1 generations were confirmed by PCR and Sanger sequencing.
Whole-genome resequencing and genotyping
Genomic DNA was extracted from leaves of B54, BoCENH3RFS33 haploid inducer, two B54 CMS lines and six B54 haploids. DNA libraries were constructed and then subjected to whole-genome sequencing using the DNBSEQ-T7 platform. Low-quality reads were filtered using Trimmomatic software and the retained clean reads were processed with the BWA software by aligning the reads to the broccoli HDEM draft genome. Uniquely mapped reads were employed in the process of whole-genome SNP calling via GATK4 following the pipeline for GATK best practices. High-quality SNPs were selected and filtered following the reported parameters34. Only sites where genotyping was available for all accessions were retained.
Haploid screening
The progenies of the BoCENH3 mutants × broccoli lines were analysed by genetic background, flow cytometry, chromosome number and plant phenotype analyses. For genetic background analyses, markers were developed on the basis of indel variations between the BoCENH3 HI inducer and broccoli test lines, and used to genotype all the progenies. For potential haploids identified by molecular markers, flow cytometry and chromosome number analyses were performed following previously described protocols35.
To determine the cytoplasmic background of the haploids or DH individuals, PCR was performed using an orf138 (the causal gene of Ogura CMS) specific marker. Information on all primers used in the study can be found in Supplementary Table 6.
Chromosome doubling
To generate DH lines, the identified CMS B54 haploid seedlings at the stage of 3–4 true leaves were treated with 200 mg l−1 colchicine solution by root dip treatment for 20 h. The ploidy levels of plants were detected after treatment.
Phenotyping and statistical analyses
Images of plant tissues were acquired using a digital camera (EOS M6, Canon). Subsequent processing of these images was conducted using Adobe Photoshop CS6 (v.13.0) and Adobe Illustrator 2022 (v.26.2). Statistical analyses were conducted using GraphPad Prism (v.9) and Microsoft Excel (2019).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The whole-genome resequencing data used in this study are accessible in NCBI through the accession code PRJNA1050660. The broccoli HDEM reference genome is publicly available (https://www.genoscope.cns.fr/projet_BKL/cgi-bin/gbrowse/boleracea/). CENH3 homologue proteins can be obtained from the database Ensembl Plants (http://plants.ensembl.org/index.html). Source data are provided with this paper.
References
Chen, L. & Liu, Y. G. Male sterility and fertility restoration in crops. Annu. Rev. Plant Biol. 65, 579–606 (2014).
Liu, C. et al. Extension of the in vivo haploid induction system from diploid maize to hexaploid wheat. Plant Biotechnol. J. 18, 316 (2020).
Kelliher, T. et al. MATRILINEAL, a sperm-specific phospholipase, triggers maize haploid induction. Nature 542, 105–109 (2017).
Gautam, R. et al. Male sterility in plants: an overview of advancements from natural CMS to genetically manipulated systems for hybrid seed production. Theor. Appl. Genet. 136, 195 (2023).
Wan, X. et al. Breeding with dominant genic male-sterility genes to boost crop grain yield in the post-heterosis utilization era. Mol. Plant 14, 531–534 (2021).
Zhong, Y. et al. Establishment of a dmp based maternal haploid induction system for polyploid Brassica napus and Nicotiana tabacum. J. Integr. Plant Biol. 64, 1281–1294 (2022).
Li, Y. et al. An in planta haploid induction system in Brassica napus. J. Integr. Plant Biol. 64, 1140–1144 (2022).
Ravi, M. et al. A haploid genetics toolbox for Arabidopsis thaliana. Nat. Commun. 5, 5334 (2014).
Zhao, X. et al. In vivo maternal haploid induction based on genome editing of DMP in Brassica oleracea. Plant Biotechnol. J. 20, 2242 (2022).
Prigge, V. et al. New insights into the genetics of in vivo induction of maternal haploids, the backbone of doubled haploid technology in maize. Genetics 190, 781–793 (2012).
Zhong, Y. et al. Mutation of ZmDMP enhances haploid induction in maize. Nat. Plants 5, 575–580 (2019).
Ravi, M. & Chan, S. W. L. Haploid plants produced by centromere-mediated genome elimination. Nature 464, 615–618 (2010).
Zhang, X. et al. A female in vivo haploid-induction system via mutagenesis of egg cell-specific peptidases. Mol. Plant 16, 471–480 (2023).
Yao, L. et al. OsMATL mutation induces haploid seed formation in indica rice. Nat. Plants 4, 530–533 (2018).
Zhong, Y. et al. In vivo maternal haploid induction in tomato. Plant Biotechnol. J. 20, 250–252 (2021).
Wang, N. et al. In planta haploid induction by genome editing of DMP in the model legume Medicago truncatula. Plant Biotechnol. J. 20, 22 (2022).
Talbert, P. B. et al. Centromeric localization and adaptive evolution of an Arabidopsis histone H3 variant. Plant Cell 14, 1053–1066 (2002).
Sanei, M. et al. Loss of centromeric histone H3 (CENH3) from centromeres precedes uniparental chromosome elimination in interspecific barley hybrids. Proc. Natl Acad. Sci. USA 108, E498–E505 (2011).
Karimi-Ashtiyani, R. et al. Point mutation impairs centromeric CENH3 loading and induces haploid plants. Proc. Natl Acad. Sci. USA 112, 11211–11216 (2015).
Wang, Z. et al. A simple and highly efficient strategy to induce both paternal and maternal haploids through temperature manipulation. Nat. Plants 9, 699–705 (2023).
Lv, J. et al. Generation of paternal haploids in wheat by genome editing of the centromeric histone CENH3. Nat. Biotechnol. 38, 1397–1401 (2020).
Wang, N. et al. Haploid induction by a maize cenh3 null mutant. Sci. Adv. 7, eabe2299 (2021).
Kindiger, B. & Hamann, S. Generation of haploids in maize: a modification of the indeterminate gametophyte (ig) system. Crop Sci. 33, 342–344 (1993).
Evans, M. M. S. The indeterminate gametophyte1 gene of maize encodes a LOB domain protein required for embryo sac and leaf development. Plant Cell 19, 46–62 (2007).
Flood, P. J. et al. Reciprocal cybrids reveal how organellar genomes affect plant phenotypes. Nat. Plants 6, 13–21 (2020).
Kuppu, S. et al. A variety of changes, including CRISPR/Cas9‐mediated deletions, in CENH3 lead to haploid induction on outcrossing. Plant Biotechnol. J. 18, 2068–2080 (2020).
Portemer, V. et al. Large genetic screens for gynogenesis and androgenesis haploid inducers in Arabidopsis thaliana failed to identify mutants. Front. Plant Sci. 6, 147 (2015).
Yu, H. et al. Creation of fertility-restored materials for Ogura CMS in Brassica oleracea by introducing Rfo gene from Brassica napus via an allotriploid strategy. Theor. Appl. Genet. 133, 2825–2837 (2020).
Shu, J. et al. Evaluation and selection of sources of cytoplasmic male sterility in broccoli. Euphytica 215, 125 (2019).
Livak, K. & Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001).
Genete, M. et al. Chromosome-scale assemblies of plant genomes using nanopore long reads and optical maps. Nat. Plants 4, 879–887 (2018).
Liu, Y. et al. Functional characterization of BoGL5 by an efficient CRISPR/Cas9 genome editing system in broccoli. Sci. Hortic. 319, 112136 (2023).
Han, F. et al. A natural mutation in the promoter of Ms-cd1 causes dominant male sterility in Brassica oleracea. Nat. Commun. 14, 6212 (2023).
Zhong, Y. et al. A DMP-triggered in vivo maternal haploid induction system in the dicotyledonous Arabidopsis. Nat. Plants 6, 466–472 (2020).
Zhu, M. et al. Introgression of clubroot resistant gene into Brassica oleracea L. from Brassica rapa based on homoeologous exchange. Hortic. Res. 9, uhac195 (2022).
Acknowledgements
This work was supported by the National Key Research and Development Program of China (Grant No. 2022YFF1003000 to F.H.), the National Natural Science Foundation of China (Grant No. 32302544 to F.H.; 32002042 to X.Z.; 32172580 to Z.L.), the China Agriculture Research System (Grant No. CARS-23-05A to Z.L.), and the Agricultural Science and Technology Innovation Program (Grant No. ASTIP to Z.L.).
Author information
Authors and Affiliations
Contributions
F.H. designed and conceived the overall research. F.H., Yuxiang Liu and X.Z. performed the experiments, analysed the data and wrote the manuscript. Yumei Liu, H.Z. and Z.L. contributed to the revision of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Plants thanks Anne Britt, Jing Li and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Figs. 1–9. Unprocessed gels for Supplementary Fig. 5.
Supplementary Tables
Supplementary Tables 1–6.
Supplementary Data
Statistical source data for Supplementary Figs. 2 and 9.
Source data
Source Data Fig. 1
Statistical source data for Fig. 1d.
Source Data Fig. 1
Unprocessed gels for Fig. 1e.
Source Data Fig. 2
Unprocessed gels for Fig. 2b.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Han, F., Zhang, X., Liu, Y. et al. One-step creation of CMS lines using a BoCENH3-based haploid induction system in Brassica crop. Nat. Plants 10, 581–586 (2024). https://doi.org/10.1038/s41477-024-01643-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41477-024-01643-w
This article is cited by
-
Haploids fast-track hybrid plant breeding
Nature Plants (2024)
