
Abstract
Many plant pathogens induce water-soaked lesions in infected tissues. In the case of Pseudomonas syringae (Pst), water-soaking effectors stimulate abscisic acid (ABA) production and signaling, resulting in stomatal closure. This reduces transpiration, increases water accumulation, and induces an apoplastic microenvironment favorable for bacterial growth. Stomata are sensitive to environmental conditions, including light. Here, we show that a period of darkness is required for water-soaking, and that a constant light regime abrogates stomatal closure by Pst. We find that constant light induces resistance to Pst, and that this effect requires salicylic acid (SA). Constant light did not alter effector-induced accumulation of ABA, but induced greater SA production, promoting stomatal opening despite the presence of ABA. Furthermore, application of a SA analog was sufficient to prevent pathogen-induced stomatal closure and water-soaking. Our results suggest potential approaches for interfering with a common virulence strategy, as well as providing a physiological mechanism by which SA functions in defense against pathogens.
Similar content being viewed by others


Introduction
The outcome of a plant-pathogen interaction is determined by multiple factors. These include environmental conditions, as well as plant defense mechanisms, and the ability of pathogens to avoid or overcome the latter1,2. Plants encode a large variety of pattern recognition receptors that can detect microbe-associated molecular patterns and induce pattern-triggered immunity (PTI)2,3,4. Early PTI responses include acidification of the apoplast, production of reactive oxygen species (ROS) and transcriptional reprogramming2,3,4. Later responses include biosynthesis of defense-related phytohormones such as salicylic acid (SA), jasmonic acid (JA) and ethylene2,4. In turn, a key virulence mechanism used by pathogens is the translocation of effector proteins into host plant cells, many of which have been shown to inhibit PTI5. Plant pathogenic bacteria possess a needle-like structure, commonly referred to as the type-3 secretion system (T3SS), that delivers effector proteins which promote disease progression in their hosts6,7. However, inhibition of defense responses is not sufficient for optimal pathogen proliferation and additional effector activities are required for the establishment of favorable microenvironments in the apoplast8,9,10,11,12.
Plant stomata are often targeted by pathogenic effector proteins or toxins as these structures represent natural breaches for pathogen entry12,13. Indeed, an important aspect of early response to pathogens involves stomatal immunity, wherein PTI responses result in stomatal closure13,14. To counteract stomatal immunity, Pseudomonas syringae (Pst) produces a JA-mimicking phytotoxin called coronatine (COR). COR has been reported to induce stomatal reopening to allow the pathogen to gain access to the apoplastic space13,15,16. COR appears to contribute more significantly to Pst virulence at night in Arabidopsis thaliana (hereafter, Arabidopsis)15, suggesting either a time of day and/or an effect of light on bacterial interaction with stomata.
In contrast to early events in infection that promote stomatal opening, pathogens appear to induce stomatal closure in later stages9,10. A frequent feature of pathogen infection in plants is the creation of an aqueous environment in the plant apoplast, visually noticeable as water-soaked lesions, which are crucial for virulence11. Pst encodes two highly conserved effector proteins, HopM1 and AvrE1, that induce water-soaking lesions9,10,11. Notably, these water-soaking effectors stimulate host abscisic acid (ABA) biosynthesis and signaling pathways to induce stomatal closure, resulting in a loss of transpiration and accumulation of water in the apoplast9,10.
Stomatal status is strongly influenced by diurnal light cycles, and light also affects plant responses to pathogens17,18. Moreover, SA regulates phytochromes (PHY) proteins, which act as photoreceptors18, and PHYA and PHYB have been reported to be required for the induction of SA-responsive genes19,20. PHYTOCHROME-INTERACTING FACTORs (PIFs) are rapidly degraded during red-light perception by PHYs and are also involved in light-dependant modulation of immune signaling21. Furthermore, functional circadian cycles are required to respond to many abiotic and biotic stresses22, including responses to infection23. Indeed, plant susceptibility to pathogens varies, depending on the time of day of infection24,25. It has been reported that plants are less susceptible in the subjective morning because defense responses are regulated by the circadian clock26. Accumulation of SA has been reported to be circadian-gated, with levels increasing throughout the day and decreasing throughout the night27. Moreover, the circadian regulators CCA1 and LHY play key roles in defense against Pst, as they promote stomatal closure in early defense responses in a SA-independent manner25. However, as circadian cycles are tightly linked to light perception, it is important to determine if a given phenomenon is related to the presence of light or to circadian mechanisms per se.
In this study, we describe the involvement of light in modulating plant defense responses. Constant light treatment prevents bacterial infection by abrogating the ability of Pst to induce stomatal closure, thereby inhibiting the accumulation of water in the apoplast. This effect is due to a potentiation of SA signaling, which induces stomatal opening despite an increase in ABA biosynthesis and signaling that would normally induce stomatal closure. In addition to highlighting the interplay between abiotic and biotic stress, our results shed light on a physiological mechanism by which SA contributes to plant defense against pathogens.
Results
Constant light confers protection against Pst DC3000
To study the influence of light on the outcome of a bacterial infection in plants, we evaluated the virulence of Pst on Arabidopsis Col-0 wild-type (WT) plants under different light regimes. Plants were grown under 12 h light/dark cycles before being challenged with Pst and placed under either constant light (LL), 12 h light/dark (LD) or constant dark (DD) regimes for 3 days. We found that Pst-infected Arabidopsis plants kept in LL showed a drastic reduction in disease symptoms compared to those kept in LD or DD over the course of the infection (Fig. 1a). This observation was reflected in bacterial titers, as plants kept under LL accumulated 10-fold less Pst compared to other conditions (Fig. 1b). Interestingly, constant light had no additional protective effect against Pst DC3000 hrcC−, which lacks a T3SS, suggesting that this regime affects effector protein actions (Fig. 1b, c).

a Disease phenotypes in Arabidopsis leaves syringe-infiltrated with Pst DC3000 (1 × 105 CFU/ml). WT Plants were either kept under constant light (LL), 12 h light/dark (LD), or constant dark (DD) light cycles for 3 days after inoculation. Bacterial titers from WT leaves syringe-infiltrated with Pst DC3000 (b) or Pst hrcC − (c) at 3 days post-infection (dpi) under light regimes described in a (n = 3 biologically independent replicates). d Apoplastic oxidative burst triggered by 1 μM flg22 in WT Arabidopsis leaf punches. Leaf punches were kept in water under LL, LD or DD for 24 h prior to stimulation with flg22. Values are averages ± se (n = 27). e Total photon counts measured over 180 min from the experiments in d. Expression levels of the early PTI marker genes FRK1 (f) and WRKY29 (g) treated with 1 μM flg22 in plants that were previously placed in the indicated light settings and measured by qRT-PCR. Samples were kept under these light settings for 6 h after control or flg22 treatment after which they were harvested (n = 3 biologically independent replicates). Different letters indicate statistically significant differences, p < 0.05, ANOVA (b, c) or p < 0.0001, ANOVA followed by Tukey’s range test (e). All statistical tests are two-sided. In box plots, box edges delineate lower and upper quartiles, the center line represents the median, and whiskers extend to 1.5 times the interquartile range (IQR). In non-box-plot graphs, boxes and error bars represent mean ± S.E.M.
Circadian rhythms and light regulation have been shown to impact bacterial virulence24,25,28. Therefore, we evaluated whether plants subjected to different light regimes for a 24-h period displayed irregular modulation of immunity following treatment with the immunogenic peptide flg22. We first explored the production of apoplastic reactive oxygen species (aROS), a commonly used early marker of immune responses. We found that plants subjected to LL for 24 h prior to treatment with flg22 produced significantly more aROS than plants kept under LD or DD regimes (Fig. 1d, e). Notably, under LL, Arabidopsis plants displayed a moderate, but persistent second wave of aROS, as opposed to other light conditions (Fig. 1d). In contrast, plants that were subjected to DD treatment for a single day produced much less aROS in response to flg22, compared to plants kept under LD conditions (Fig. 1e). Hence, light appears to prime Arabidopsis plants to produce enhanced and more sustained aROS after immune elicitation, potentially contributing to disease resistance.
We evaluated the expression of two early PTI-responsive genes, FRK1 and WRKY29, to assess early PTI responsiveness. Plants were subjected to the three different light regimes for 24 h before being challenged with flg22 for 4 h. Although not significantly altered, these genes were less expressed in DD compared to LD or LL (Fig. 1f, g). Interestingly, constant light by itself resulted in increases in both FLS2 and RBOHD expression levels in the absence of flg22, although only the latter was statistically significant (Supplementary Fig. 1a, b). Contrary to this observation, FLS2 and RBOHD expression levels were not upregulated upon flg22 treatment under constant dark (Supplementary Fig. 1a, b). These results suggests that light perception and/or signaling is critical for the establishment of a proper early PTI response.
Pst DC3000 requires darkness to induce water-soaking lesions
The virulence of Pst is greatly enhanced by its ability to induce water-soaking lesions by closing stomata under high humidity conditions9,10. As light modulates stomatal aperture29, we questioned whether a pathogen’s ability to induce stomatal closure could be affected by diurnal light oscillations. We observed that, under LL conditions, Pst could no longer induce water-soaking lesions in Arabidopsis leaves, in contrast to LD or DD (Fig. 2a). The latter was not due to differences in levels of bacteria, as inoculations were carried with a high concentration of bacteria (1 × 108 CFU/ml) (Fig. 2b). Consistent with the lack of water soaking, we found that stomata of inoculated Arabidopsis plants kept under LL conditions were still fully open 24 h post inoculation (Fig. 2c). In contrast, stomata of inoculated plants kept under LD conditions were closed, as expected, as were those of plants kept under DD conditions (Fig. 2c).

a Water-soaking phenotypes in Arabidopsis leaves syringe-infiltrated with Pst DC3000 (1 × 108 CFU/ml) at 24 hours post-infection (hpi) under LL, LD or DD conditions. b Bacterial titers from plants described in a (n = 3 biologically independent replicates). c Stomatal aperture measurements in plants described in a by epifluorescence microscopy (n = 194 stomata for LL Control, n = 190 stomata for LL DC3000, n = 229 stomata for LD Control, n = 196 stomata for LD DC3000, n = 185 stomata for DD Control, n = 262 stomata for DD DC3000). d Water-soaking phenotypes in Arabidopsis leaves that were syringe-infiltrated with Pst DC3000 (1 × 108 CFU/ml) at 24 hpi under LL or LD conditions (left panel) and in plants treated in the same way followed by transferred to DD for 6 h (right panel). e Stomatal aperture measurements from leaves treated as in d (n = 220 stomata for Mock and n = 156 stomata for DC3000 [upper left panel], n = 182 stomata for Mock and n = 227 stomata for DC3000 [upper right panel], n = 252 stomata for Mock and n = 207 for DC3000 [bottom left panel], n = 229 stomata for Mock and n = 195 stomata for DC3000 [bottom right panel]). Different letters indicate statistically significant differences, p < 0.05, ANOVA (b) or Kruskal–Wallis test (c). Asterisks indicate statistically significant differences compared to control, *p < 0.05, ****p < 2.2 × 10−16, Student’s t test (e, upper right panel) or Wilcoxon–Mann–Whitney test (e, upper left, lower right and lower left panels). All statistical tests are two-sided. In box plots, box edges delineate lower and upper quartiles, the center line represents the median, and whiskers extend to 1.5 times the interquartile range (IQR).
To consolidate our findings, we designed an experiment where Arabidopsis plants were infected with Pst and either kept under LL or LD conditions for the first 16 h of the infection. At this timepoint, LD, but not LL treated plants underwent water soaking (Fig. 2d). Plants were then placed in darkness to determine if water soaking could be reappearing. Indeed, we found that the water-soaking phenotype could be fully restored after keeping LL treated plants in darkness for 6 h (Fig. 2d). Stomatal behavior analysis further supported the observation that constant light prevents Pst from inducing stomatal closure, but that this phenomenon is swiftly reversed once under dark conditions (Fig. 2e).
Pst manipulates its host ABA machinery via secretion of its conserved effectors HopM1 and AvrE1 to induce stomatal closure leading to water-soaking9,10. We designed experiments to test whether a deficiency in apoplast hydration was the main reason behind reduced Pst aggressivity under LL. Since the Arabidopsis aba2-1 mutant was shown to prevent Pst-induced water-soaked lesions9,10, we compared the bacterial levels of these plants infected under LL and LD conditions. We found that, while bacterial population levels differed between LL and LD conditions in WT plants, they were unaltered in aba2-1 plants (Supplementary Fig. 2). Moreover, under LL, Pst grew to levels similar to Pst hopM1−/avrE1−(h−/a−), which cannot induce water-soaking (Supplementary Fig. 3). Furthermore, the pathogen’s ability to manipulate ABA appeared unaffected as transcript levels of the ABA biosynthesis and signaling marker genes, NCED3 and RD29A, respectively, in WT infected plants were similar under all light conditions (Supplementary Fig. 4). These results suggest that the lack of apoplastic fluid could at least partly explain the growth deficiency of Pst under LL. Thus, it appears that pathogens might benefit from natural light cycles to establish a favorable microenvironment inside host leaves.
SA is required to prevent water-soaking under continuous light
SA is a key immune signaling molecule and has been reported to antagonize multiple actions driven by ABA signaling30,31,32. However, how SA and ABA signaling interact with each other with respect to stomatal control during late phases of pathogen infection is unknown. We investigated whether plants subjected to a constant light cycle for 24 h would be altered in SA signaling or accumulation. First, we analyzed expression levels of SA responsive-genes, ICS1 (ISOCHORISMATE SYNTHASE 1) and PR1 (PATHOGENESIS-RELATED PROTEIN 1), which are often used to assess SA biosynthesis and responsiveness, respectively. We observed that the expression levels of these two genes were increased under LL conditions relative to LD or DD regimes, although the increase of ICS1 relative expression was not significant (Fig. 3a, b). Consistent with this, we observed, using ultra performance liquid chromatography-mass spectrometry (UPLC-MS), that upon treatment with flg22, plants kept under LL conditions showed an increase in SA levels compared to LD and DD treated plants (Fig. 3c). Likewise, upon infiltration of plants with Pst, plants grown in LL conditions showed a greater induction of ICS1 and accumulation of SA than plants grown in LD or DD (Supplementary Fig. 5). As such, constant light appears not only to modulate early PTI responses, but also SA-related responses.

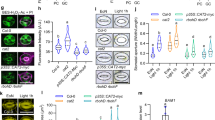
a, b Expression levels, measured by qRT-PCR, of SA biosynthesis (ICS1) and signaling (PR1) marker genes in WT Arabidopsis plants treated with 1 μM flg22 after having previously been placed in the identified light settings. Samples were harvested at 24 hpi (n = 3 biologically independent replicates). c Quantification of SA in WT Arabidopsis plants mock inoculated (control) or inoculated with 1 μM flg22 under the same experimental settings as in a measured by UPLC-MS at 24 hpi (n = 3 biologically independent replicates). d Water-soaking phenotypes in Arabidopsis WT (Col-0), sid2 and npr1 mutant plants syringe-infiltrated with Pst DC3000 (1 × 108 CFU/ml) at 24 hpi under LL, LD or DD. e Stomatal aperture measurements from infected leaves displayed in d (n = 159 stomata for all conditions). f Bacterial titers from Arabidopsis WT (Col-0), sid2 and npr1 mutant plants infected with Pst DC3000 (1 × 108 CFU/ml) at 24 hpi (n = 3 biologically independent replicates). g Bacterial titer of Arabidopsis WT (Col-0), sid2 and npr1 mutant plants infected with Pst DC3000 (1 × 105 CFU/ml) in which infiltrated leaves were allowed to return to a pre-infiltrated state (−H2O) or in plants that were immediately domed following infiltration (+H2O). Bacterial titers were measured at 3 dpi (n = 3 biologically independent samples). Asterisks indicate statistically significant differences compared to control, ns non-significant, *p < 0.05, **p < 0.001, ****p < 2.2 × 10−16, ANOVA followed by Tukey’s range test compared to LD (c), Student’s t test (e, f) or Wilcoxon–Mann–Whitney test (e, g). All statistical tests are two-sided. In box plots, box edges delineate lower and upper quartiles, the center line represents the median, and whiskers extend to 1.5 times the interquartile range (IQR). In non-box-plot graphs, boxes and error bars represent mean ± S.E.M.
In agreement with our findings, a recent report revealed that SA-related responses are primed by photoperiodic stress33. Considering that SA can antagonize ABA responses, we investigated whether SA is necessary for light-mediated prevention of water-soaked lesions. We inoculated Arabidopsis mutants, npr1-1 and sid2-2, which are compromised in SA signaling and biosynthesis, respectively, with Pst under different light conditions. Interestingly, we observed water-soaking in the inoculated leaves of npr1-1 and sid2-2 mutant plants at 24 h post-infection (hpi) under both LD and LL conditions, whereas wild-type plants showed water-soaking only under LD conditions (Fig. 3d). Consistent with these results, we observed that Pst inoculation induced stomatal closure at 24 hpi in all genotypes under LD conditions (Fig. 3e). Furthermore, apoplastic hydration was elevated in LL in SA-related mutants, but not in Col-0 under LL (Supplementary Fig. 6). However, constant light did not hamper the ability of the pathogen to induce stomatal closure in the npr1-1 mutant, or (to a lesser degree) in the sid2-2 mutant, as it does in WT plants (Fig. 3e). These altered responses were not due to differences in bacterial densities, as bacterial titers did not differ in mutant versus wild-type plants at this time point (Fig. 3f).
The impact of SA on plant responses to infection may be due to multiple effects. Given the effect of SA on water-soaking and stomatal control (Fig. 3d, e), we tested if the effect of SA might be related to the establishment of an aqueous environment, which is beneficial to bacteria. To do so, we employed an inoculation method wherein plants are immediately placed in a >95% humidity environment after inoculation without allowing liquid from the inoculum to evaporate. Under these conditions, leaves remain saturated with water, mimicking at least in part the phenomenon of water-soaking. Artificial induction of water soaking had no effect on bacterial growth of either wild-type or mutant plants under LD conditions (Fig. 3g). Keeping plants under LL conditions resulted in a dramatic decrease in bacterial growth in WT plants, while this growth defect was partially rescued in sid2-2 and npr1-1 mutants (Fig. 3g). Importantly, artificial water-soaking resulted in a dramatic increase of bacterial growth under LL conditions, with both WT and mutant plants showing bacterial levels similar to LD grown plants (Fig. 3g). These results indicate that the induction of resistance induced by constant light requires SA signaling and that the effects of SA are, at least in part, related to its effects on inhibiting stomatal closure. This, in turn, inhibits the pathogen’s ability to create an ideal aqueous environment in the apoplast.
BTH suppresses Pst-induced water-soaking lesions
Benzothiadiazole (BTH) is an analog of SA and, like SA, triggers systemic acquired resistance in plants and confers protection against Pst34,35. Since SA biosynthesis and signaling are essential for preventing water-soaking under LL (Fig. 3d), we explored the potential of using BTH to prevent this phenomenon. Plants that were pre-treated with BTH 24 h prior to a Pst challenge did not display water-soaking lesions, in contrast to control-treated plants (Fig. 4a). Most importantly, BTH prevented Pst-induced water-soaking lesions under a LD regime, in which water-soaking lesions are normally visible. Consistent with this result, Pst did not induce stomatal closure in LD-grown plants pre-treated with BTH (Fig. 4b). These effects were independent of the number of bacteria, as bacterial titers in leaves did not differ between treatments at the time of stomatal evaluation (Fig. 4c).

a Water-soaking phenotypes in WT Arabidopsis infiltrated with DMSO (0.1%) or BTH (50 μM) prior to syringe-infiltration with Pst DC3000 (1 × 108 CFU/ml) 24 h later. Photos were taken 24 h post-infection. b Stomatal aperture in WT Arabidopsis treated as in a and in which MgCl2 (10 mM; −DC3000) or Pst (1 × 108 CFU/ml; +DC3000) was syringe-filtrated. Plants were left under light/dark conditions and stomatal apertures assessed at 24 h post infiltration (hpi) (n = 202 stomata for all conditions). c Bacterial titers from plants treated in a measured 24 hpi (n = 3 biologically independent replicates). d Bacterial titers from Arabidopsis leaves that were infiltrated with DMSO (0.1%) or BTH (50 μM) 24 h prior to being challenged with Pst DC3000 (1 × 105 CFU/ml). Plants were placed under the indicated light conditions after infection. Bacterial titers were assessed at 3 days post-infection and plants were kept under the indicated light conditions throughout the infection process (n = 3 biologically independent replicates). Asterisks indicate statistically significant differences compared to control, ns non-significant, *p = 0.001, ***p < 0.0001, ****p < 2.2 × 10−16, Student’s t test (b for Control, c) or Wilcoxon–Mann–Whitney test (b for BTH, d). All statistical tests are two-sided. In box plots, box edges delineate lower and upper quartiles, the center line represents the median, and whiskers extend to 1.5 times the interquartile range (IQR).
We next tested the ability of BTH to mitigate a Pst infection under either LL or LD in locally pre-treated leaves. Interestingly, levels of bacteria in control or BTH pre-treated leaves were similar under constant light, suggesting that BTH does not induce more resistance the effect of a 24-h period of light alone (Fig. 4d). However, a clear difference was observed between control and BTH pre-treated plants under LD conditions (~100-fold less bacteria). At the same time, LD-grown plants pre-treated with BTH showed a protection similar to albeit statistically significantly different from that accorded by growing under LL (5.69 ± 0.15 CFU/cm2 (LL); 6.25 ± 0.36 CFU/cm2 (LD)) (Fig. 4d). These results suggest that BTH provides protection similar to that provided by a constant light treatment, and further indicates that the effects of constant light in this context are due to the actions of SA.
Constant light allows plants to recover from bacterial infection
Given the role of light in preventing disease symptoms and pathogen growth, we evaluated whether constant light could mitigate the progression of the pathogen in plants that were already infected. To test this, plants were inoculated and left under a LD cycle for a day to allow the pathogen to establish an infection. Plants were then transferred to LL conditions for 2 days, then again to a LD cycle for two additional days. Control plants were similarly inoculated but kept under a LD cycle for the duration of the experiment (Fig. 5a). No disease symptoms were observed on plant leaves that had been subjected to the 2-day LL treatment (Fig. 5b). Bacterial titer assays were also performed and a decrease in bacterial load was observed when plants were kept under LL conditions compared to plants kept under LD conditions for the entire experiment (Fig. 5c). These results suggest that constant light treatment allows plants to recover from a Pst infection.

a Schematic diagram representing treatments and phenotype assessment of light-mediated disease prevention. This figure was created with BioRender.com. b Disease symptoms in Arabidopsis plants infected with 1 × 105 CFU/ml under different light settings, as, indicated. c Bacterial titers from plants shown in b and harvested at the indicated timepoints (n = 3 biologically independent replicates). Bacterial titers were significantly different between plants subjected to a LL treatment versus control (Student’s t test, p = 0.0024) d Stomatal aperture measurements of leaves evaluated as in c (n = 160 stomata for 1 dpi LD control and DC3000, 3 dpi LL control and LD DC3000; n = 161 stomata for 3 dpi LL DC3000 and LD control; n = 163 stomata for all data at 5 dpi). Letters indicate statistically significant differences compared to control with Student’s t test (1 dpi) or Wilcoxon–Mann–Whitney test (3 and 5 dpi). Data points and error bars represent mean ± S.E.M. In box plots, box edges delineate lower and upper quartiles, the center line represents the median, and whiskers extend to 1.5 times the interquartile range (IQR).
We also assessed stomatal aperture over the course of a 5-day infection period in the same experiment. As expected, infiltration with Pst induced stomatal closure in LD conditions within 24 h compared to control plants (Fig. 5d). At 3 dpi, plants kept under LD conditions maintained this phenotype to an even greater degree, presumably due to increased bacterial numbers (Fig. 5c, d). In contrast, moving plants to LL conditions resulted in stomatal opening similar to uninoculated plants (Fig. 5d). Consistent with a report showing that phytocytokines induce reopening of stomates at late stages of infection36, we observed that previously closed stomata reopened at 5 dpi even when plants were returned to a LD regime (Fig. 5d). This is consistent with observations that water-soaking lesions are present in the early phases of Pst infection, and disappear in later phases11. Nonetheless, our results show that it is possible to revert pathogen-induced stomatal closure to prevent disease progression by placing infected plants under a constant light regime for 2 days.
PIFs may modulate disease resistance through SA signaling
We investigated the implication of light photoreception and signaling in susceptibility to bacterial infection under LL or LD regimes. For this, we used the Arabidopsis blue-light insensitive (blus1) mutant as well as a quadruple mutant for phytochrome-interacting factors (pifq), which is responsible for integrating red and far/red light signaling. The Arabidopsis blus1 mutant behaved similarly to wild-type plants in terms of water-soaking under LD and LL (Fig. 6a). However, pifq mutants displayed a reestablishment of water-soaked lesions 24 h after inoculation with Pst (Fig. 6a). These observations were consistent with measurements of stomatal aperture, wherein only pifq mutants infected with Pst had closed stomata under constant light (Fig. 6b). Interestingly, all three genotypes showed similar increases of induction of expression of ICS1 upon infection with Pst, with somewhat higher levels in LL versus LD (Fig. 6c). However, upon inoculation with Pst, WT and blus1 plants showed much higher levels of induction of PR1 expression in LL versus LD conditions, whereas pifq plants did not (Fig. 6d). At the same time, upon inoculation with Pst, pifq mutant plants kept under LL conditions showed no statistically significant difference (Wilcoxon Rank Sum test) in PR1 induction to WT plants grown in a LD regime (Fig. 6d). Thus, the levels of PR1 induction and stomatal closure in LL-grown pifq plants resemble those of LD-grown WT plants, further underlining the link between SA signaling, stomatal closure, and water-soaking. Light photoreception mutants displayed no significant differences in susceptibility toward a Pst infection compared to WT plants under different light regimes (Supplementary Fig. 7). This can be explained by the fact that water-soaking symptom appearance in pifq mutant requires a high load of bacteria upon infiltration, while a low load is used for bacterial growth assays. These results thus suggest that PIFs might not be absolutely required for, but may be involved in, reinforcing the SA module during constant light after immune elicitation.

a Water-soaking phenotypes of Arabidopsis WT (Col-0), blus1 and pifq mutant plants syringe-infiltrated with Pst 3000 (1 × 108 CFU/ml) under the indicated light settings. Photos were taken at 24 h post infiltration (hpi). b Stomatal aperture measurements from infected leaves displayed in a (n = 192 stomata for all LD DC3000 data, n = 196 stomata for all control data, n = 199 stomata for all LL DC3000 data). Expression levels, assessed by qRT-PCR, of ICS1 (c) and PR1 (d) marker genes in Arabidopsis WT (Col-0), blus1 and pifq mutant plants infected with Pst DC3000 (1 × 108 CFU/ml) or with control solution (MgCl2 10 mM) in plants that were placed in the identified light settings. Samples were harvested at 24 hpi (n = 3 biologically independent replicates). Asterisks indicate statistically significant differences compared to control, ns non-significant, *p < 0.05, ****p < 2.2 × 10−16, Wilcoxon–Mann–Whitney test (b) or ****p < 0.0001, Student’s t test (d). All statistical tests are two-sided. In box plots, box edges delineate lower and upper quartiles, the center line represents the median, and whiskers extend to 1.5 times the interquartile range (IQR). In non-box-plot graphs, boxes and error bars represent mean ± S.E.M.
Discussion
Induction of a water-soaked apoplast is crucial for disease development in plants11,37,38,39. At the same time, it is known that this can be affected by light. For example, in an early study of fire blight, it was observed that “as the sun came out, many of these areas (water-soaked lesions) cleared up completely”40. In this study, we provide evidence that diurnal light cycles are intimately linked with the ability of a bacterial pathogen to establish an aqueous apoplast. We show that a period of darkness is required to induce water-soaking and that constant light prevent induction of stomatal closure mediated by water-soaking effectors. Light appears to counteract the stomatal-closing effects of ABA through potentiation of SA responses. Furthermore, BTH, a SA analog, was sufficient to prevent bacterial induction of water-soaking lesions. Whether SA signaling directly inhibits some aspect of ABA action in this case remains to be determined. Considering how common the induction of water-soaked lesions is as an initial step in microbial pathogenesis37, finding non-invasive and easily applicable solutions to prevent them is of great practical interest.
In studies of the circadian clock machinery plants are often placed in constant light to study oscillations in gene regulation and other phenomena41. As such, much of our understanding of circadian-gated regulation of phytohormone networks rely heavily on data obtained under constant light regimes27,42,43. However, light can affect molecular processes independently of circadian mechanisms. Indeed, Arabidopsis plants with mutations in key circadian cycle-regulating genes are protected against infection when grown under LL conditions compare to LD. Importantly, the degree of protection is similar to that seen with WT plants (Supplementary Fig. 8), suggesting that the effects of constant light are not dependant on the circadian cycle.
Our data suggest that, under constant light regimes, plant immune responses are augmented or primed. Indeed, apoplastic ROS bursts following flg22 perception are more intense under LL conditions and less intense under DD (Fig. 1d). These results are consistent with the elevated levels of expression of FLS2, RBOHD, ICS1 and PR1 under LL conditions prior to flg22 treatment (Supplementary Figs. 1 and 3a, b). Furthermore, the expression levels of early PTI-responsive genes, FRK1 and WRKY29, were significantly increased under constant light conditions prior to, and after, treatment with flg22 (Fig. 1f, g). Nonetheless, consistent with increased aROS production, plants grown under LL conditions appear to respond with greater intensity to flg22 in terms of SA accumulation (Fig. 3c). How light affects the responsiveness to immune elicitors remains to be investigated. However, our results showing that it affects aROS production and SA-related responses, but not early PTI responses, suggest that constant light affects specific modules in plant immunity. At the same time, our results are consistent with a recent study showing that, under photoperiodic stress, the Arabidopsis transcriptional signature resembles that of a response to pathogens33. Notably, SA signatures increased in plants kept under constant light and were reduced in dark-grown plants.
It has been reported that pathogen aggressiveness is increased in infections occurring in the shade44. However, whether this involves host stomatal responses and is related to pathogen-induced water-soaking lesions has not been investigated. Diurnal light changes affect stomatal aperture, where stomata are open during the day and partially close during the night45. As such, a synergistic interaction may occur in which the mechanism(s) behind dark-induced stomatal closure, while not well defined, may act as facilitators for effector-mediated induction of stomatal closure. It is important to note that, in infected plants, light-induced inhibition of stomatal closure and water-soaking can be rapidly reversed. That is, non-water-soaked infected plants with LL and SA-dependent open stomata undergo water-soaking after a 4-h dark treatment (Fig. 2d, e). This suggests that dark conditions may be a prerequisite for the induction of water soaking. Current understanding of the mechanism behind darkness-induced stomatal closure suggest that ABA signaling and metabolism are important for the response speed, but not essential for closure, suggesting ABA-dependent and independent mechanisms46. Thus, we speculate that the balance between the differential effects of ABA and SA on stomatal aperture may be affected by additional light-dependent mechanisms (Fig. 7). Whether this is related to previously described antagonistic effects of SA on ABA signaling30,31,32 remains to be determined. Likewise, this could include regulation of ABA levels, which increase substantially within a 4-h period of darkness47.

In continuous light (left), HopM1 and AvrE1-mediated stimulation of ABA biosynthesis and signaling occurs. However, elevated SA accumulation in continuous light conditions ensures that stomata remain open despite the pro-closure influence of ABA, thus preventing water-soaking. When plants experience a period of darkness (right), the pro-closure influence of ABA predominates, possibly due to reduced SA accumulation, leading to water-soaking. This figure was created with BioRender.com.
Water-soaking effectors of Pst modulate ABA-related responses9,10. The induction of expression of ABA biosynthesis and signaling pathways by Pst was unaltered between all light regimes tested (Supplementary Fig. 4). Indeed, SA and ABA mutants do not show large differences in the accumulation of ABA and SA, respectively upon bacterial infection, suggesting that the reported SA antagonism of ABA-related responses may be downstream of ABA-responsive targets30,31,32. Our observations are reminiscent of a previous report showing that SA accumulation antagonizes ABA responses, including stomatal closure, even if ABA over accumulates and certain ABA-responsive genes are activated at the transcriptional level32. Whether SA-mediated antagonism of ABA responses is solely responsible for the pathogen’s inability to induce an aqueous apoplast remains to be investigated.
ABA is a major susceptibility factor to most known ABA producing or ABA inducing plant pathogens48. In addition to Pst, exposure to photoperiodic stress leads to enhanced disease resistance to the hemi-biotrophic fungus M. oryzae and to the necrotrophic fungus B. cinerea49,50. Interestingly, these two fungal pathogens produce ABA and induce water-soaking lesions early on during the infection process. Whether water-soaking contributes to fungal pathogenicity is not clear, but ABA was found to be a key virulence component for M. oryzae and B. cinerea51,52. It is thus interesting to speculate that the reduced virulence of these fungal pathogens in plants exposed to photoperiodic stress is caused by their inability to create an aqueous apoplast as well. Therefore, exposing plants to constant light for a short amount of time could be a very impactful strategy to reduce infections from pathogens from different kingdoms of life.
We are still in the dark about the specific wavelengths of light that affect the defense responses described herein. However, it has been reported that red light treatment improves resistance against Pst and that this treatment increased SA signaling as well as transcription of genes involved in redox homeostasis, such as RBOH and GSTs53. Our results provides further evidence for the role of red light in mediating resistance54, as a quadruple mutant of PIF genes, which are involved in red light signaling, displays water-soaking in constant light, whereas this is not observed in the Arabidopsis blue light signaling mutant blus1 (Fig. 6a). Arabidopsis pifq mutant displayed less SA potentiation upon pathogen inoculation, which could be the reason behind the induction of water-soaking in this mutant. Further research will be required to dissect how red-light circuits are integrated into immune responses.
In summary, our study better defines a physiological mechanism for disease resistance and adds to the growing trend demonstrating that plant defenses involve starving pathogens for water and nutrients11,55. We provide a proof of concept showing that altering light conditions for relatively short periods of time can limit pathogen damage (Fig. 5) and suggest that this could have broad applications. While controlling photoperiods to prevent disease outbreaks is not as applicable for field production, this simple method could be used to controlled environment food production, which is growing quickly and is linked to urban food sovereignty. At the same time, our current and previous work9,10,39 indicate that any treatment that leads to increased stomatal opening and transpiration leads to decreases in pathogen virulence. As such, strategies to prevent pathogen-induction of water-soaking could lead to broad-spectrum disease resistance.
Methods
Plant material
Arabidopsis plants were grown in PromixTM soil (PremierTech) in growth chambers either with 12 h light/dark, 24 h light or 24 h dark photoperiod, with relative humidity of ~60% at 21 °C. Light intensity was measured at 180 µmoles/m2/s in all growth chambers when lights were on. Four- to 5-week-old Arabidopsis plants were used for all experiments described.
Bacterial disease assays
Pseudomonas syringae pv. tomato DC3000 WT and mutant strains were cultured overnight at 28 °C in Luria-Bertani (LB) media containing 50 mg/l of rifampicin. On the day of the infection, fresh LB media was inoculated with 0.5 µl of the overnight culture and bacteria were collected when OD600 reached between 0.8–1. Bacteria were centrifuged for 10 min at 4000 × g and the pellet resuspended in MgCl2 10 mM. Bacterial density was adjusted to 0.2 (1 × 108 CFU/ml) prior to further dilutions. Bacterial infections were carried between 14:00–15:00, or a zeitgeber time of 6:00–7:00.
All inoculations of Arabidopsis leaves were performed by syringe-infiltration. Infiltrated plants were all kept under ambient humidity levels for 1–2 h to allow water to evaporate, then domed with a plastic unit to maintain high humidity (>95% RH), unless stated otherwise.
Bacterial growth in planta was monitored by harvesting infected Arabidopsis leaves, surface sterilizing in 80% ethanol and rinsing in sterile water twice. Leaf disks were taken from three leaves from the same plant (one per leaf; total of three leaf disks) using a cork borer (0.6 mm in diameter) and ground in sterile 10 mM MgCl2. Three biological replicates were performed for each experiment. Colony-forming units (CFU) were determined by making serial dilutions (100−10−6) and plating on LB plates containing 50 mg/l of rifampicin. Each dilution was plated in three technical replicates. Experiments were repeated at least three times and values from replicates are tabulated in the Source Data File.
ROS quantification
Leaf disks from 4-week-old Arabidopsis plants were collected using a 4 mm diameter biopsy punch and placed into white 96-well plates (Corning) containing 100 μl of distilled water for 16 h (overnight). Prior to ROS quantification, the water was removed and replaced with ROS assay solution (100 μM Luminol [Millipore-Sigma], 20 μg/ml horseradish peroxidase [Millipore-Sigma] with or without immune elicitors). Light emission was measured using a TECAN Spark® plate reader. Experiments were repeated three times and values from replicates are tabulated in the Source Data File.
Stomatal aperture assays
In LL and LD experiments, plants were sampled during the subjective day (i.e., light conditions). In DD experiments, plants were sampled from dark conditions. Leaves were cut at the base of the petiole and immediately immersed in the stomatal fixation solution (formaldehyde 4% and rhodamine 6 G 0.5 µM) for 1 min to stop stomatal movement. A quarter of each leaf was cut with a razor blade and stomata were observed by epifluorescence microscopy. Stomatal apertures were measured using OMERO software and the degree of the stomatal opening was measured as a ratio of stomatal width to length. Between 150 to 250 stomata were measured for each data point. Data points represented in stomatal aperture measurement figures originated from at least three leaves from three different plants. Data collection and analysis was performed by using double-blinded standards to avoid bias. Values from replicates are tabulated in the Source Data File.
Chemical treatments
To assess plant immune responses, Arabidopsis leaves were treated with flg22 (1 μM; BioBasic) by syringe-infiltration. For BTH protection assay, Arabidopsis leaves were syringe-infiltrated with BTH (50 μM; Millipore-Sigma).
RNA extraction and real-time quantitative PCR
RNA was extracted from frozen and ground leaf tissue using QIAZOL (QIAGEN) reagents, followed by on-column DNase treatment (QIAGEN), according to the manufacturer’s protocol. RNA purity was assessed with a spectrophotometer and quality by gel electrophoresis. cDNA was generated by using MMuLV-RT (Service de purification des protéines—Université de Sherbrooke).
Quantitative real-time PCR was performed with a Bio-Rad CFX96 machine. Each reaction contained 1X SYBR mix (Service de purification des protéines—Université de Sherbrooke), specific primers and a 1:20 dilution of 500 ng of cDNA stock. Amplification cycle protocols were as follow: 2 min at 95 °C; 40 cycles of 6 s at 95 °C and 30 s at 60 °C. Melting curves were verified at the end of 40 cycles for confirmation of primer specificities. All reactions were repeated in three technical and biological replicates. Average Cq values were normalized by ∆∆CT formula against ACTIN2 expression. Oligos used in this study can be found in Supplementary Table 1. Experiments were repeated at least three times and values from replicates are tabulated in the Source Data File.
Apoplast extraction
Three fully expanded leaves per plant from three plants were excised and apoplast extracted as previously described56, with some modifications. Briefly, the initial weight of freshly excised leaves was measured before being infiltrated with distilled water and weighed again once leaves were fully saturated with water. Leaves were centrifuged at 4000 rcf in 2 ml microcentrifugation tubes containing glass beads at the bottom to prevent tissue collapse during the centrifugation. Leaves were weighed post centrifugation and apoplast hydration determined as previously described56. Experiments were repeated at least three times and values from replicates are tabulated in the Source Data File.
Salicylic acid extraction and quantification
Fully expanded 4-week-old Arabidopsis leaves were harvested and weighed for fresh weight calculation and immediately flash-freeze in liquid nitrogen. Tissues were ground with a plastic pestle and phytohormones were extracted overnight using 0.5–1 ml of ice-cold extraction buffer (methanol: water [80:20 v/v], 0.1% formic acid, 0.1 g/l butylated hydroxytoluene and 100 nM ABA-d6 as an internal standard). Extracts were filtered using centrifugal filter units.
Filtered extracts were quantified using an Acquity Ultra Performance Liquid Chromatography system (Waters Corporation, Milford, MA) as described previously10. SA was quantified based on a standard curve to calculate sample concentration (nM), which was converted to ng using the molecular weight of each specific compound and the extraction volume used. All data was normalized to initial fresh weight in grams. Experiments were repeated three times and values from replicates are tabulated in the Source Data File.
Statistical analysis
Statistical analyses were performed in GraphPad Prism 8.0 for all RT-qPCR experiments, photon counts, and SA quantification. For all other experiments (bacterial counts and stomatal assays), the bioinformatic software R was used. p values greater than 0.05 were considered non-significant. Sample sizes, statistical tests used, and p values are stated in figure legends. All statistical test assumptions such as normality and homoskedasticity were tested. When not respected, non-parametric equivalent tests were performed. In multiple comparison tests, Bonferroni corrections were used. Data points represented in stomatal aperture measurement figures originated from at least three leaves from three different plants.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Source data are provided with this paper.
References
Cheng, Y. T., Zhang, L. & He, S. Y. Plant-microbe interactions facing environmental challenge. Cell Host Microbe 26, 183–192 (2019).
Nishad, R., Ahmed, T., Rahman, V. J. & Kareem, A. Modulation of plant defense system in response to microbial interactions. Front. Microbiol. 11, 1298 (2020).
Han, G.-Z. Origin and evolution of the plant immune system. N. Phytol. 222, 70–83 (2019).
Yuan, M., Ngou, B. P. M., Ding, P. & Xin, X.-F. PTI-ETI crosstalk: an integrative view of plant immunity. Curr. Opin. Plant Biol. 62, 102030 (2021).
Toruño, T. Y., Stergiopoulos, I. & Coaker, G. Plant-pathogen effectors: cellular probes interfering with plant defenses in spatial and temporal manners. Annu. Rev. Phytopathol. 54, 419–441 (2016).
Collmer, A. et al. Pseudomonas syringae Hrp type III secretion system and effector proteins. PNAS 97, 8770–8777 (2000).
Lee, J. et al. Type III secretion and effectors shape the survival and growth pattern of Pseudomonas syringae on leaf surfaces. Plant Physiol. 158, 1803–1818 (2012).
Gentzel, I. et al. Dynamic nutrient acquisition from a hydrated apoplast supports biotrophic proliferation of a bacterial pathogen of maize. Cell Host Microbe 30, 502–517.e4 (2022).
Hu, Y. et al. Bacterial effectors manipulate plant abscisic acid signaling for creation of an aqueous apoplast. Cell Host Microbe 30, 518–529.e6 (2022).
Roussin-Léveillée, C. et al. Evolutionarily conserved bacterial effectors hijack abscisic acid signaling to induce an aqueous environment in the apoplast. Cell Host Microbe 30, 489–501.e4 (2022).
Xin, X.-F. et al. Bacteria establish an aqueous living space in plants crucial for virulence. Nature 539, 524–529 (2016).
Xin, X.-F., Kvitko, B. & He, S. Y. Pseudomonas syringae: what it takes to be a pathogen. Nat. Rev. Microbiol. 16, 316–328 (2018).
Melotto, M., Underwood, W., Koczan, J., Nomura, K. & He, S. Y. Plant stomata function in innate immunity against bacterial invasion. Cell 126, 969–980 (2006).
Melotto, M., Zhang, L., Oblessuc, P. R. & He, S. Y. Stomatal defense a decade later. Plant Physiol. 174, 561–571 (2017).
Panchal, S. et al. Coronatine facilitates Pseudomonas syringae infection of Arabidopsis leaves at night. Front. Plant Sci. 7, 880 (2016).
Toum, L. et al. Coronatine inhibits stomatal closure through guard cell-specific inhibition of NADPH oxidase-dependent ROS production. Front. Plant Sci. 7, 1851 (2016).
Carvalho, S. D. & Castillo, J. A. Influence of light on plant-phyllosphere interaction. Front. Plant Sci. 9, 1482 (2018).
Shah, A. et al. A comprehensive review on the influence of light on signaling cross-talk and molecular communication against phyto-microbiome interactions. Crit. Rev. Biotechnol. 41, 370–393 (2021).
Genoud, T., Buchala, A. J., Chua, N.-H. & Métraux, J.-P. Phytochrome signalling modulates the SA-perceptive pathway in Arabidopsis. Plant J. 31, 87–95 (2002).
Karpinski, S., Gabrys, H., Mateo, A., Karpinska, B. & Mullineaux, P. M. Light perception in plant disease defence signalling. Curr. Opin. Plant Biol. 6, 390–396 (2003).
Gangappa, S. N. & Kumar, S. V. DET1 and COP1 modulate the coordination of growth and immunity in response to key seasonal signals in Arabidopsis. Cell Rep. 25, 29–37.e3 (2018).
Roeber, V. M., Schmülling, T. & Cortleven, A. The photoperiod: handling and causing stress in plants. Front. Plant Sci. 12, 781988 (2022).
Karapetyan, S. & Dong, X. Redox and the circadian clock in plant immunity: a balancing act. Free Radic. Biol. Med. 119, 56–61 (2018).
Griebel, T. & Zeier, J. Light regulation and daytime dependency of inducible plant defenses in Arabidopsis: phytochrome signaling controls systemic acquired resistance rather than local defense. Plant Physiol. 147, 790–801 (2008).
Zhang, C. et al. Crosstalk between the circadian clock and innate immunity in Arabidopsis. PLoS Pathog. 9, e1003370 (2013).
Bhardwaj, V., Meier, S., Petersen, L. N., Ingle, R. A. & Roden, L. C. Defence responses of Arabidopsis thaliana to infection by Pseudomonas syringae are regulated by the circadian clock. PLoS ONE 6, e26968 (2011).
Zheng, X.-Y. et al. Spatial and temporal regulation of biosynthesis of the plant immune signal salicylic acid. Proc. Natl Acad. Sci. USA 112, 9166–9173 (2015).
Wang, W. et al. Timing of plant immune responses by a central circadian regulator. Nature 470, 110–114 (2011).
Matthews, J. S. A., Vialet-Chabrand, S. & Lawson, T. Role of blue and red light in stomatal dynamic behaviour. J. Exp. Bot. 71, 2253–2269 (2020).
de Torres Zabala, M., Bennett, M. H., Truman, W. H. & Grant, M. R. Antagonism between salicylic and abscisic acid reflects early host-pathogen conflict and moulds plant defence responses. Plant J. 59, 375–386 (2009).
Meguro, A. & Sato, Y. Salicylic acid antagonizes abscisic acid inhibition of shoot growth and cell cycle progression in rice. Sci. Rep. 4, 4555 (2014).
Moeder, W., Ung, H., Mosher, S. & Yoshioka, K. SA-ABA antagonism in defense responses. Plant Signal Behav. 5, 1231–1233 (2010).
Cortleven, A. et al. Photoperiod stress in Arabidopsis thaliana induces a transcriptional response resembling that of pathogen infection. Front. Plant Sci. 13, 838284 (2022).
Görlach, J. et al. Benzothiadiazole, a novel class of inducers of systemic acquired resistance, activates gene expression and disease resistance in wheat. Plant Cell 8, 629–643 (1996).
Kim, J. H. et al. Increasing the resilience of plant immunity to a warming climate. Nature 607, 1–6 (2022).
Liu, Z. et al. Phytocytokine signalling reopens stomata in plant immunity and water loss. Nature 605, 332–339 (2022).
Aung, K., Jiang, Y. & He, S. Y. The role of water in plant–microbe interactions. Plant J. 93, 771–780 (2018).
Peng, Z. et al. Xanthomonas translucens commandeers the host rate-limiting step in ABA biosynthesis for disease susceptibility. Proc. Natl Acad. Sci. USA 116, 20938–20946 (2019).
Zhang, D., Tian, C., Yin, K., Wang, W. & Qiu, J.-L. Postinvasive bacterial resistance conferred by open tomata in rice. Mol. Plant Microbe Interact. 32, 255–266 (2019).
Clayton, E. Water soaking of leaves in relation to development of the wildfire disease of tobacco. J. Agric. Res. 52, 239–269 (1936).
Velez-Ramirez, A. I., van Ieperen, W., Vreugdenhil, D. & Millenaar, F. F. Plants under continuous light. Trends Plant Sci. 16, 310–318 (2011).
Li, Z., Bonaldi, K., Uribe, F. & Pruneda-Paz, J. L. A localized Pseudomonas syringae infection triggers systemic clock responses in Arabidopsis. Curr. Biol. 28, 630–639.e4 (2018).
Zhou, M. et al. Redox rhythm reinforces the circadian clock to gate immune response. Nature 523, 472–476 (2015).
Roberts, M. R. & Paul, N. D. Seduced by the dark side: integrating molecular and ecological perspectives on the influence of light on plant defence against pests and pathogens. N. Phytol. 170, 677–699 (2006).
Yang, J., Li, C., Kong, D., Guo, F. & Wei, H. Light-mediated signaling and metabolic changes coordinate stomatal opening and closure. Front. Plant Sci. 11, 601478 (2020).
Pridgeon, A. J. & Hetherington, A. M. ABA signalling and metabolism are not essential for dark-induced stomatal closure but affect response speed. Sci. Rep. 11, 5751 (2021).
Weatherwax, S., Ong, M. S., Degenhardt, J., Bray, E. & Tobin, E. The interaction of light and abscisic acid in the regulation of plant gene expression. Plant Physiol. https://doi.org/10.1104/pp.111.2.363 (1996).
Lievens, L., Pollier, J., Goossens, A., Beyaert, R. & Staal, J. Abscisic acid as pathogen effector and immune regulator. Front. Plant Sci. 8, 587 (2017).
Cagnola, J. I. et al. Long-day photoperiod enhances jasmonic acid-related plant defense. Plant Physiol. 178, 163–173 (2018).
Shimizu, S., Yamauchi, Y. & Ishikawa, A. Photoperiod following inoculation of Arabidopsis with Pyricularia oryzae (syn. Magnaporthe oryzae) influences on the plant-pathogen interaction. Int. J. Mol. Sci. 22, 5004 (2021).
Audenaert, K., De Meyer, G. B. & Höfte, M. M. Abscisic acid determines basal susceptibility of tomato to Botrytis cinerea and suppresses salicylic acid-dependent signaling mechanisms. Plant Physiol. 128, 491–501 (2002).
Spence, C. A., Lakshmanan, V., Donofrio, N. & Bais, H. P. Crucial roles of abscisic acid biogenesis in virulence of rice blast fungus Magnaporthe oryzae. Front. Plant Sci. 6, 1082 (2015).
Yang, Y.-X. et al. RNA-seq analysis reveals the role of red light in resistance against Pseudomonas syringae pv. tomato DC3000 in tomato plants. BMC Genomics 16, 120 (2015).
Pham, V. N., Kathare, P. K. & Huq, E. Phytochromes and phytochrome interacting factors. Plant Physiol. 176, 1025–1038 (2018).
Yamada, K., Saijo, Y., Nakagami, H. & Takano, Y. Regulation of sugar transporter activity for antibacterial defense in. Arabidopsis. Sci. 354, 1427–1430 (2016).
Gentzel, I., Giese, L., Zhao, W., Alonso, A. P. & Mackey, D. A simple method for measuring apoplast hydration and collecting apoplast contents. Plant Physiol. 179, 1265–1272 (2019).
Acknowledgements
This study was supported by two Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grants respectively to P.M. and I.L.-L., as well as by a Fonds de Recherche du Québec—Nature et Technologies (FRQ-NT) Team Grant and NOVA-NSERC Alliance Grant to P.M. (PI) and I.L.-L. (co-PI). I.L.-L. is also supported by a Canada Research Chair T2. G.L. was supported by an Excellence Scholarship from the Faculty of Science of Université de Sherbrooke and by a Canada Graduate Scholarship (NSERC-MSc). C.R.-L. was supported by a VoiceAge excellence fellowship and an FRQ-NT PhD scholarship. We are grateful to Philippe Venne (Chemistry Department, Université de Sherbrooke) for assistance with UPLC-MS analyses.
Author information
Authors and Affiliations
Contributions
C.R.-L. and P.M. conceived the study. G.L. and C.R.-L. performed all experiments, data analysis and the creation of figures. S.B. and E.F. contributed to some experiments and data collection. I.L.-L. contributed to the statistical analysis. G.L. and C.R.-L. wrote the manuscript with support from P.M. and I.L.-L. All authors revised and agreed with the content written in this manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Jian Hua, Yukun Liu and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lajeunesse, G., Roussin-Léveillée, C., Boutin, S. et al. Light prevents pathogen-induced aqueous microenvironments via potentiation of salicylic acid signaling. Nat Commun 14, 713 (2023). https://doi.org/10.1038/s41467-023-36382-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-023-36382-7
This article is cited by
-
Extracellular niche establishment by plant pathogens
Nature Reviews Microbiology (2024)
-
How plants manage pathogen infection
EMBO Reports (2023)
-
The effectors of Phytophthora infestans impact host immunity upon regulation of antagonistic hormonal activities
Planta (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
