
Abstract
Iron deficiency hampers photosynthesis and is associated with chlorosis. We recently showed that iron deficiency-induced chlorosis depends on phosphorus availability. How plants integrate these cues to control chlorophyll accumulation is unknown. Here, we show that iron limitation downregulates photosynthesis genes in a phosphorus-dependent manner. Using transcriptomics and genome-wide association analysis, we identify two genes, PHT4;4 encoding a chloroplastic ascorbate transporter and bZIP58, encoding a nuclear transcription factor, which prevent the downregulation of photosynthesis genes leading to the stay-green phenotype under iron-phosphorus deficiency. Joint limitation of these nutrients induces ascorbate accumulation by activating expression of an ascorbate biosynthesis gene, VTC4, which requires bZIP58. Furthermore, we demonstrate that chloroplastic ascorbate transport prevents the downregulation of photosynthesis genes under iron-phosphorus combined deficiency through modulation of ROS homeostasis. Our study uncovers a ROS-mediated chloroplastic retrograde signaling pathway to adapt photosynthesis to nutrient availability.
Similar content being viewed by others

Introduction
Chloroplasts are sites of photosynthesis, whose function requires numerous proteins encoded in the nuclear genome1. Although plants tightly orchestrate chloroplast-to-nucleus signaling (retrograde control), it is poorly understood at the mechanistic level. In addition, the adequate accumulation of nutrients such as iron (Fe) in chloroplasts is required for their optimal performance2,3. Up to 80% of Fe in leaves is located in the chloroplasts4,5, where its ability to donate and accept electrons plays a central role in electron transfer reactions6. Fe is found in all electron transfer complexes PSI, PSII, cytochrome b6f complex, and ferredoxins and is required for the biogenesis of cofactors such as hemes and iron–sulfur clusters7,8. Plants grown under Fe-deficient (−Fe) environments show chlorotic symptoms9, and compromised photosynthesis2,3. However, chlorotic leaves can also develop under high-phosphorus (P) conditions, despite replete Fe levels10, challenging the causal connection between Fe concentration and chlorophyll accumulation. Moreover, we recently reported that rice plants grown under a combined Fe and P deficiency (−Fe−P) do not exhibit a chlorotic phenotype11. These observations revealed a gap in our understanding of the interdependent effects of nutrient availability on regulating photosynthesis. Here, we addressed this issue through a combination of global gene expression analyses and genome-wide association studies (GWAS) to find expression quantitative trait loci (eQTLs) and uncovered a regulatory module that controls chlorophyll accumulation in response to Fe and P availability. This module involves an ascorbic acid (AsA) synthesis enzyme named VITAMINC4 (VTC4), a chloroplastic AsA transporter named PHOSPHATE TRANSPORTER 4;4 (PHT4;4), and a putative transcription factor named BASIC LEUCINE-ZIPPER 58 (bZIP58). The functioning of this module sheds light on the importance of chloroplast–nucleus communications under co-occurring nutrient deficiencies in controlling photosynthesis.
Results
Iron-induced chlorosis is independent of Fe status
We previously reported that Fe-deficiency-induced chlorosis depends on P availability in rice11. To investigate whether the interdependent effects of Fe and P availability on chlorosis are conserved across monocot and eudicot species, we phenotyped Arabidopsis thaliana Col-0 (eudicot) and Lemna gibba (monocot), along with Oryza sativa (monocot), under different regimes of Fe and P availability. Fe deficiency (−Fe) caused chlorosis in all three species, but only in the presence of P (−Fe+P) (Fig. 1a–c). Quantification of chlorophyll content confirmed that −Fe significantly reduced the accumulation of chlorophyll in all three species (Fig. 1d). However, under −Fe−P conditions, chlorophyll content was comparable to control (+Fe+P) conditions in these species (Fig. 1d). Next, we focused on Arabidopsis to gain insights into the physiological and molecular processes underlying the stay-green phenotype (lack of chlorosis development) under −Fe−P conditions. First, we asked whether the stay-green phenotype under −Fe−P is caused by an increase of Fe levels in shoots. Plants grown in −Fe+P conditions decreased total Fe in shoots by twofold compared to +Fe+P conditions (Supplementary Fig. 1). On the other hand, under +Fe−P conditions, Fe levels increased by 2.2-fold relative to +Fe+P conditions (Supplementary Fig. 1). Surprisingly, Fe levels in plants grown under −Fe−P were reduced and indistinguishable from the Fe levels in −Fe+P conditions (Supplementary Fig. 1). Therefore, the stay-green phenotype under −Fe−P appears independent of shoot Fe status. Furthermore, examination of bioavailable Fe2+ in leaves using the Turnbull/DAB (stains Fe2+)12,13 staining revealed enhanced accumulation of Fe under +Fe−P and reduced Fe under −Fe+P compared to control (+Fe+P) (Fig. 1e). Under −Fe−P, Fe accumulation was also reduced compared to control and indistinguishable from −Fe+P, indicating that the higher chlorophyll content under −Fe−P compared to −Fe+P is not due to an increase in bioavailable Fe (Fig. 1e). Similarly, quantification using the phenanthroline method14 detected no difference in bioavailable Fe2+ accumulation under −Fe+P and −Fe−P conditions (Fig. 1f). Taken together, these results show that the onset of chlorosis during −Fe requires sufficient P in the growth media, and that the “stay-green” phenotype under the combined −Fe−P deficiency is unlikely due to Fe nutritional status in leaves.

a–c Duckweed (Lemna gibba), rice (Oryza sativa cv Nipponbare), and Arabidopsis thaliana plants grown on media containing iron and phosphorus (+Fe+P), deficient in iron (−Fe+P), or deficient in both elements (−Fe−P). Representative images of L. gibba propagated for 28 days (a), 24-day-old rice (b) and 14-day-old A. thaliana (Col-0) (c) are shown. The experiment was repeated with similar results three times. d Mean chlorophyll accumulation in L. gibba, A. thaliana, and O. sativa grown under +Fe+P, −Fe+P, and −Fe−P conditions. Scale bars: 7 mm (a), 10 mm (b), and 5 mm (c). Data shown are from three experiments with 3–10 plants per experiment. Error bars represent 95% confidence intervals. FW: fresh weight. e Turnbull staining of Fe2+ in leaves of seedlings grown on +Fe+P for ten days and transferred to +Fe+P, +Fe−P, −Fe+P or −Fe−P conditions for 7 days. The experiment was repeated with similar results three times. f Mean soluble iron content in shoots of fourteen-day-old A. thaliana plants grown on agar plates containing +Fe+P, +Fe−P, −Fe+P, or −Fe−P. Data shown are from three experiments with ten plants per experiment. Error bars represent 95% confidence intervals. DW dry weight. For d and f, letters above bars represent statistically different means within each species-specific comparison at P < 0.05 (one-way ANOVA with a Duncan post hoc test). Source data are provided as a Source Data file.
Phosphorus availability modulates the effect of iron deficiency on chlorophyll accumulation and photosystem activity
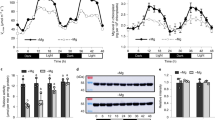
To understand the cause of chlorophyll reduction in response to −Fe, we first explored the timing of −Fe sensing and photosynthetic function response. Since −Fe affects chlorophyll accumulation and photosystem II (PSII) activity15,16, we monitored their kinetics over 172 h (h) (Fig. 2a, b and Supplementary Fig. 2A–C). Arabidopsis plants were first grown on +Fe+P media for 1 week, and then transferred to +Fe+P, −Fe+P, or −Fe−P conditions. −Fe+P caused a significant decrease in chlorophyll content, which was observable starting at 52 h after the transfer to −Fe+P (Fig. 2a). However, transfer to −Fe−P did not affect chlorophyll content, even at 172 h after the transfer (Fig. 2a). To determine how photosynthesis was affected, we measured Fv/Fm, which reflects the quantum yield of photochemistry and is a measure of PSII activity15,16. Plants under −Fe+P decreased Fv/Fm, which was observable starting at 52 h, indicative of compromised electron transport through PSII, and which coincides with the decrease of chlorophyll accumulation (Fig. 2b). By 172 h, PSII activity was substantially reduced under −Fe+P compared to +Fe+P. However, plants under −Fe−P showed slightly lower but stabilized Fv/Fm compared to those in +Fe+P (Fig. 2b). Similarly, non-photochemical quenching decayed as a function of time under −Fe+P whereas the decay was substantially less in −Fe−P (Supplementary Fig. 2D). These physiological characterizations showed that chlorophyll accumulation and photosystem activity were affected by −Fe, and both responses were P-dependent (Fig. 2a, b and Supplementary Fig. 2D).

a, b Chlorophyll content and PSII activity (Fv/Fm) in response to iron and/or phosphate deficiency in A. thaliana. Seedlings were grown for 7 days in the presence of iron and phosphorus (+Fe+P) and transferred to three different media: +Fe+P, −Fe+P, or −Fe−P for 15, 28, 39, 52, 76, 100, 124, 148, and 172 h. a Mean chlorophyll data shown are from three experiments with eight plants per experiment. Error bars represent 95% confidence intervals. FW fresh weight. b Mean Fv/Fm data shown are from three experiments with 16 plants per experiment. Error bars represent the 95% confidence intervals. c Experimental design for transcriptomic studies on A. thaliana (Col-0) shoots. Plants were grown in +Fe+P media for 7 days before transfer to three different media conditions (+Fe+P, −Fe+P, or −Fe−P). Shoots were harvested for RNA extraction and sequencing at the 39, 52, and 76 h time points. d Global expression analysis of genes in response to −Fe+P and −Fe−P relative to +Fe+P. Numbers of genes displaying at least twofold change in their expression are shown for each condition. The 32 genes that were decreased specifically in −Fe+P but not in −Fe−P relative to +Fe+P at 52 h and 76 h (highlighted in purple) were used to perform genome-wide association studies. e A heatmap showing gene expression patterns of the 32 genes in −Fe+P and −Fe−P relative to control (+Fe+P) at 39, 52, and 76 h after transfer. LogFC log2 fold change. f Gene Ontology enrichment for the 32 genes whose mRNA abundance was specifically decreased by −Fe+P. FDR false-discovery rate. Source data are provided as a Source Data file.
Phosphorus and iron interaction regulates the expression of photosynthesis-related genes (PRGs)
Based on these findings, we selected three time points at 39, 52, and 76 h after the transfer of plants to +Fe+P, −Fe+P, or −Fe−P to conduct a global gene expression analysis in shoots (Fig. 2c and Supplementary Fig. 3A–F). We identified genes whose expression levels were either increased or decreased by −Fe+P relative to +Fe+P by at least twofold at a P value <0.05 (Fig. 2d and Supplementary Data 1). Even more genes were either upregulated or downregulated in −Fe−P conditions relative to +Fe+P (Fig. 2d and Supplementary Fig. 4A–C). A total of 673 and 2434 transcripts were specifically differentially regulated in response to −Fe+P or −Fe−P, respectively, supporting the existence of different signaling pathways under the two conditions (Supplementary Fig. 4A–C). To identify functions enriched in genes that were differently regulated by −Fe+P or −Fe−P, we performed Gene Ontology (GO) enrichment analysis. The common set of 52 genes that were specifically downregulated by −Fe−P at 52 h and 76 h after the transfer (Supplementary Fig. 5A and Supplementary Data 1) showed enrichment for ribosomal genes (Supplementary Fig. 5B) while upregulated genes (162 genes, Supplementary Fig. 5C, Supplementary Data 1) revealed enrichment for genes involved in cation transport, response to water, and ester hydrolysis (Supplementary Fig. 5D and Supplementary Data 1). On the other hand, GO analysis of the 32 genes specifically downregulated by −Fe+P but not affected by −Fe−P at 52 h and 76 h (Fig. 2e and Supplementary Data 1) revealed an enrichment of genes related to the chloroplast and photosynthesis-related processes (Fig. 2f), while upregulated genes (Supplementary Fig. 5E) were enriched for genes related to cellular respiration, oxidation–reduction process, and energy metabolism (35 genes; Supplementary Fig. 5F). Altogether, the transcriptomics analysis indicated that the control of chloroplast function is an integral component of the nuclear transcriptomic response to −Fe, which is dependent on P availability. We also learned that the photosynthesis-related phenotypes we observed under −Fe+P, but not under −Fe−P, could be due to the downregulation of key photosynthesis regulators.
PHT4;4-mediated AsA transport in the chloroplast prevents chlorosis in −Fe−P conditions
To decode the signaling pathways that control the expression of the photosynthesis genes in response to −Fe+P, we exploited natural variation in expression of the 32 genes that were downregulated by Fe deficiency in a P-dependent manner in a worldwide collection of A. thaliana accessions17. One way to identify mechanisms regulating the expression of photosynthesis-related genes (PRGs) could be to identify genetic factors associated with their natural variation of expression across Arabidopsis accessions. For this, we retrieved the published mRNA levels of the 32 photosynthesis genes in shoots of 727 Arabidopsis accessions17. Strikingly, their expression was generally positively associated with each other across the accessions, indicating that they are co-regulated (Supplementary Fig. 6A). Next, we performed principal component analysis (PCA) to reduce the dimensionality of expression data for these 32 genes. PC1 explained 89.5% of the variation in their expression (Supplementary Fig. 6B), which we then used to perform a genome-wide association study (GWAS) (Fig. 3a). Our GWA analysis detected 38 QTLs containing 145 candidate genes in total (Fig. 3a). In this study, we followed up two QTLs, the first one located on chromosome 1 (SNP4653399) containing nine candidate genes (AT1G13570 to AT1G13610) (Supplementary Fig. 7), and the second QTL located on chromosome 4 (SNP171674) containing six candidate genes (AT4G00355 to AT4G00400) (Supplementary Fig. 7). All of the candidate genes underlying these two QTLs were subjected to functional genetic analysis to identify the causal gene(s) that influence the expression of the PRGs and chlorophyll content.

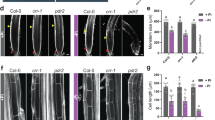
a A Manhattan plot for genome-wide association mapping using principal component 1 that explained 89.5% of expression variation of the 32 photosynthesis-related genes across 727A. thaliana accessions17. The five chromosomes are depicted by alternating light- and dark-blue colors. Dashed lines correspond to significant SNPs determined using an accelerated mixed model with adjustment for multiple tests using a FDR 5% threshold (blue) and Bonferroni α = 0.05 (red). The light-gray rectangle highlights a significant association located in an intergenic region (SNP: 4493712). Two significant associations that were followed up in this study are highlighted in red rectangles. b Representative images of wild-type Col-0, pht4;4, vtc4, and a line expressing genomic PHT4;4 in pht4;4 (pht4;4-CL) grown for 7 days in the presence of iron and phosphorus (+Fe+P) and transferred to three different media: +Fe+P, −Fe+P, or −Fe−P for 7 additional days. Scale bars: 7 mm. The experiment was repeated with similar results three times. c Total chlorophyll content in Col-0, pht4;4, vtc4, and PHT4;4-CL grown under +Fe+P, −Fe+P, −Fe−P, or −Fe+P+AsA. FW: fresh weight. Data shown are from ten plants conducted in three independent experiments. d Relative mRNA abundance of VTC genes (VTC1 (AT2G39770), VTC2 (AT4G26850), and VTC4 (AT3G02870)) in shoots of Col-0 plants grown on +Fe+P media for 7 days and transferred to +Fe+P, −Fe+P, or −Fe−P media for 52 h. Data shown are the means from 3 experiments. Error bars represent the 95% confidence interval. e Total ascorbic acid (AsA) content in Col-0, pht4;4, vtc4, and PHT4;4-CL plants grown for 7 days on +Fe+P media and transferred to +Fe+P, −Fe+P, or −Fe−P media for 52 h. Data shown are from three experiments, each with 16 plants. In box plots (c, e), center lines indicate sample medians; box limits indicate the 25th and 75th percentiles; whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles. For c–e, letters above bars or boxes represent statistically different means at P < 0.05 (one-way ANOVA with a Duncan post hoc test). Data points are plotted as open circles. Source data are provided as a Source Data file.
Analysis of mutants of genes underlying the QTL for chlorophyll accumulation on chromosome 4 (associated with SNP171674) showed that genetic inactivation of only one gene, AT4G00370 (PHT4;4), failed to stay green under −Fe−P conditions, and exhibited a remarkable decrease in chlorophyll content by comparison to wild-type plants (Fig. 3b, c and Supplementary Fig. 8). Introduction of the wild-type PHT4;4 allele into a pht4;4 mutant background complemented these phenotypes (Fig. 3b, c). PHT4;4 encodes a protein that transports inorganic phosphate (Pi) as well as ascorbic acid (AsA) to the chloroplast18. To test whether other chloroplastic Pi transporters18 influenced chlorophyll accumulation under combined Fe and P deficiency, we characterized mutant plants for PHT4;3, PHT4;5, and PHT4;6. Chlorophyll content was not distinguishable in any of these mutants compared to wild-type plants in −Fe−P conditions (Supplementary Fig. 9A). Furthermore, plants lacking key genes regulating Pi transport or homeostasis, the transcription factor PHOSPHATE RESPONSE 1 (PHR1) and the SPX DOMAIN GENE 1 (SPX1), showed similar chlorophyll content as wild-type plants (Supplementary Fig. 9B). Taken together, these results support the idea that the effects of pht4;4 mutants on chlorophyll content are likely mediated through its AsA transport activity.
To test our hypothesis about the role of AsA in preventing chlorosis under −Fe−P conditions, we first assessed how Fe and P availabilities regulate the expression of VITAMIN C (VTC) enzymes involved in AsA biosynthesis in plants19. Our RNA-seq analysis indicated that −Fe−P caused a two- to threefold increase in VTC1 (AT2G39770), VTC2 (AT4G26850), and VTC4 (AT3G02870) expression, which we confirmed using qRT-PCR (Fig. 3d and Supplementary Fig. 10A). However, −Fe in the presence of P (−Fe+P) caused about a twofold decrease in the mRNA abundance of VTC4 (Fig. 3d and Supplementary Fig. 10A). VTC4 is the final enzyme in the AsA biosynthesis pathway19. This prompted us to test the effect of the absence of VTC4 on chlorophyll accumulation under −Fe+P and −Fe−P conditions. Under −Fe−P, mutants with a vtc4 null allele were still chlorotic, similarly to pht4;4 and in contrast to wild-type plants (Fig. 3b, c). These data show that AsA contributes to preventing chlorosis in −Fe−P conditions.
Next, we tested whether the chlorotic phenotype is due to variations in AsA levels. In wild-type, AsA levels decreased significantly under −Fe+P at 52 h after the transfer relative to control (+Fe+P), whereas no change was detected under −Fe−P (Fig. 3e), suggesting that AsA levels were associated with −Fe-mediated chlorosis. To test whether AsA levels were associated with chlorosis in general, we measured AsA levels in AsA synthesis (vtc4) mutant plants. Under +Fe+P, vtc4 plants accumulated 35% less AsA than wild-type plants, and AsA levels remained unchanged in response to −Fe+P or −Fe−P stress (Fig. 3e). However, vtc4 plants did not show the chlorotic phenotype under +Fe+P, which indicated that the level of AsA contributed to the chlorotic phenotype specifically under −Fe and this contribution was dependent on P availability. In addition, the AsA transporter (pht4;4) mutants showed similar AsA levels as the wild type even though pht4;4 plants were still chlorotic in −Fe−P (Fig. 3e). To determine whether AsA accumulation in the cell or its transport to the chloroplast is associated with the development of chlorotic phenotype in −Fe+P, we tested the effect of an exogenous supply of AsA in wild type, vtc4, and pht4;4 plants (Fig. 3b, c). Exogenous AsA alleviated the chlorosis caused by −Fe+P in wild-type and vtc4 mutant plants. However, pht4;4 mutants failed to stay green under −Fe+P+AsA conditions (Fig. 3b, c), indicating that the transport of AsA to the chloroplast is required for −P-mediated “stay-green” phenotype under Fe deficiency. Our results showed that −P prevents the downregulation of VTC4 by −Fe and associated changes in AsA accumulation, and that the PHT4;4-mediated transport of AsA to chloroplasts is required for the maintenance of chlorophyll content under combined deficiency of Fe and P.
We next asked whether PHT4;4-mediated AsA transport to the chloroplast is important for regulation of the PRGs that were specifically downregulated by −Fe in a P-dependent manner. First, we tested the effects of PHT4;4 inactivation on the expression of these photosynthesis-related genes using qRT-PCR (Fig. 4a and Supplementary Fig. 10B, and Supplementary Data 2). While −Fe+P significantly downregulated the mRNA abundance of these genes in wild-type plants (Col-0), −Fe−P prevented this response (Fig. 4a and Supplementary Fig. 10B, and Supplementary Data 2). Furthermore, adding AsA to −Fe+P mimicked −Fe−P response in preventing downregulation of the photosynthesis genes (Fig. 4a, Supplementary Fig. 10B, and Supplementary Data 2). While under −Fe+P PRGs are expressed in pht4;4 to similar levels to that of wild-type plants, PRGs were downregulated in pht4;4 plants under −Fe−P as well as −Fe+P supplemented with AsA (Fig. 4a and Supplementary Fig. 10B). Taken together, these data indicate AsA regulates the expression of PRGs in a pht4;4-dependent manner.

a Average relative expression of 32 PRGs in Col-0 (red), pht4;4 (light blue), and bzip58 (brown) in plants grown for 7 days in the presence of iron and phosphorus (+Fe+P) and transferred to +Fe+P, −Fe+P, −Fe−P, or −Fe−P + AsA for 76 h. Letters above markers represent statistically different means at P < 0.05 (one-way ANOVA with a Duncan post hoc test). Error bars = 95% CI. Data for all PRGs were averaged from three independent experiments. Relative expression level of PRGs was determined relative to Ubiquitin 10. b Relative mRNA abundance of bZIP58 in shoots of Col-0 plants grown for 7 days on +Fe+P and transferred to +Fe+P, −Fe+P, −Fe−P, or −Fe−P+H2O2 for 76 h. Data shown are the means from three experiments. Error bars represent 95% confidence intervals. c Relative mRNA abundance of VTC4 in the shoots of Col-0 and bzip58 mutants grown in the presence of +Fe+P for 7 days and transferred to +Fe+P, −Fe+P, or −Fe−P for 76 h. Data shown are the means three experiments. Error bars represent 95% confidence intervals. d Confocal microscopy images of p35S::bZIP58::GFP expressing plants grown for 7 days under +Fe+P. Scale bars: 20 μm. The experiment was repeated with similar results three times. e Representative images of Col-0, bzip58, and a line expressing genomic bZIP58 in bzip58 mutants (bzip58-CL) grown for 7 days in +Fe+P and transferred to +Fe+P, −Fe+P, −Fe−P, or −Fe+P+AsA for 7 additional days. Scale bars: 7 mm. The experiment was repeated with similar results three times. f Total chlorophyll content in Col-0, bzip58, and bZIP58-CL plants grown for 7 days in +Fe+P and transferred to +Fe+P, −Fe+P, −Fe−P, or −Fe+P+AsA for 7 days. FW: fresh weight. Data shown are from four experiments. g Accumulation of H2O2 (a type of ROS) in shoots of Col-0, pht4;4 and bzip58 plants grown for 7 days in +Fe+P and transferred to +Fe+P, −Fe+P, or −Fe−P for 52 h. Data shown from 12 experiments. In box plots (f, g), center lines show sample medians; box limits indicate the 25th and 75th percentiles; whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles. For b, c, f, and g, letters above bars or boxes represent statistically different means at P < 0.05 (one-way ANOVA with a Duncan post hoc test). Data points are plotted as open circles. Source data are provided as a Source Data file.
bZIP58 regulates the expression of photosynthesis-related genes (PRGs) in a Fe-dependent manner
We next sought to determine how −Fe+P affected expression of the 32 PRGs. To look for potential transcriptional regulators of these genes, we screened candidate genes from the GWAS analysis. We found that bZIP58 (AT1G13600) (Fig. 3a and Supplementary Figs. 6 and 7), a putative transcription factor, underlies one of the strongest QTL peaks (Fig. 3a). To characterize bZIP58 further, we first asked whether Fe and P deficiencies affected its expression. Fe limitation strongly downregulated bZIP58 in shoots, and P limitation alleviated the repression of bZIP58 caused by Fe deficiency (Fig. 4b and Supplementary Fig. 10A). bZIP58 expression was not altered by +Fe−P, indicating that P availability alone does not influence bZIP58 expression (Supplementary Fig. 10C). In comparison to shoots, we found a much weaker downregulation of bZIP58 by −Fe in roots (Supplementary Fig. 10D). Given that AsA supplementation prevented the development of chlorosis caused by Fe limitation, we next asked how AsA would influence the expression of bZIP58, and whether bZIP58 regulates the expression of VTC4 (Fig. 4c). Remarkably, AsA prevented the downregulation of bZIP58 by −Fe+P. This led us to examine the contribution of bZIP58 in regulating the −Fe+P-specific PRGs under +Fe+P, −Fe+P, and −Fe−P conditions (Fig. 4a, Supplementary Fig. 10B, and Supplementary Data 2). Mutants with the bzip58 null allele showed a remarkable constitutive decrease in the expression of these 32 PRGs (Fig. 4a, Supplementary Fig. 10B, and Supplementary Data 2). bZIP58 localizes to the nucleus (Fig. 4d), which is consistent with a role as a transcription factor. Furthermore, to ask whether bZIP58 can directly regulate any of the 32 PRGs, we performed transactivation assays using bZIP58 as an effector and the promoters of five randomly selected PRGs (PHOTOSYSTEM II SUBUNIT T (PSBTN); PHOTOSYSTEM II 5 kD, PLASTID TRANSCRIPTIONALLY ACTIVE 16 (PTAC16), LIGHT HARVESTING COMPLEX PHOTOSYSTEM II (LHCB4.1) and PHOTOSYSTEM II REACTION CENTER W (PSBW)) fused to GUS as reporters (Supplementary Fig. 11A) showed that bZIP58 can activate the promoter activity of three of the five genes tested (PSBTN, PHOTOSYSTEM II 5 kD, and PTAC16) (Supplementary Fig. 11B). Therefore, bZIP58 can directly activate the expression of some PRGs. These results demonstrate that bZIP58 is a key gene to regulate the expression of PRGs in a Fe-dependent manner.
Taken together, these findings support the idea that bZIP58 is a key regulator of PRGs, and its absence could alter chlorophyll accumulation regardless of Fe and P availability. Genetic inactivation of bZIP58 indeed causes a constitutive decrease in chlorophyll content, and the mutant line is chlorotic (Fig. 4e, f). The expression of the bZIP58 gene in bzip58 plants complements the constitutive chlorosis phenotype, and the complemented line responds to Fe and P availability similarly to wild-type plants (Fig. 4e, f). Furthermore, AsA supplementation could not rescue the chlorotic phenotype of bzip58 mutants (Fig. 4e, f), indicating that bZIP58 lies downstream of AsA action. In addition, bZIP58 was partially required to induce VTC4 expression under −Fe−P conditions (Fig. 4c). These data show that bZIP58 controls the expression of PRGs and is transcriptionally regulated in response to −Fe depending on P availability, likely by mediating the perception of AsA.
Plastid ROS influences chlorophyll content and PRGs expression in −Fe+P conditions
We next asked how AsA in the chloroplast could affect photosynthesis. AsA has an antioxidizing action that detoxifies reactive oxygen species (ROS) through its scavenging properties20, thus making ROS a potential signaling molecule21,22,23 capable of modulating the expression of PRGs through bZIP58. To test this hypothesis, we first measured the relative amount of ROS accumulation in shoots of wild-type, pht4;4, and bzip58 plants under various Fe and P availability. −Fe+P caused a twofold increase in ROS accumulation in shoots of wild-type plants, which partially depended on P availability (Fig. 4g). pht4;4 plants displayed comparable ROS accumulation to that of the wild-type under +Fe+P and −Fe+P. However, pht4;4 plants accumulated significantly higher ROS than wild-type plants under −Fe−P (Fig. 4g). In addition, bzip58 mutant plants displayed a constitutive increase in ROS accumulation (Fig. 4g). To check whether ROS in turn can regulate the expression of bZIP58, we quantified bZIP58 expression in response to foliar application of H2O2. ROS treatment caused a 4-fold decrease in bZIP58 transcript accumulation, and this effect was independent of the availability of Fe or P in the medium (Fig. 4b and Supplementary Fig. 10C). Our results supported the role of ROS as a plastid signal to influence chlorophyll content and PRGs expression in −Fe+P conditions. To test this idea, we expressed bZIP58 in bzip58 (Line A1) and bzip58xpht4;4 mutant plants (Line B1) under the control of a ROS-responsive promoter of a gene called JUNGBRUNNEN1 (JUB1)24 (Fig. 5a). −Fe induced the expression of JUB1p::bZIP58 by 2.5-fold in bzip58 and by fourfold in bzip58xpht4;4 (Fig. 5a). While WT and bzip58-CL lines were chlorotic and showed repression of PRGs in −Fe+P conditions (Fig. 4e, f), the expression of JUB1p::bZIP58 enhanced chlorophyll accumulation and expression of PRGs in the bzip58 mutant (Fig. 5b, c and Supplementary Fig. 12). These responses were exacerbated in bzip58xpht4;4 double mutant expressing JUB1p::bZIP58 (Fig. 5b, c and Supplementary Fig. 12). Greatly enhanced induction of JUB1p::bZIP58 by −Fe in bzip58xpht4;4 compared to bzip58 indicates the sensitivity of this ROS-responsive promoter to PHT4;4-dependent plastid signal. Remarkably, AsA supplementation in −Fe+P prevented the induction of JUB1p::bZIP58 in the bzip58 mutant but not in bzip58xpht4;4 double mutant (Fig. 5b, c and Supplementary Fig. 12). Consistently, Line A1 was chlorotic, which was associated with repression of PRGs, and Line B1 accumulated chlorophyll under −Fe+P+AsA conditions and exhibited high expression levels of PRGs (Fig. 5b–d and Supplementary Fig. 12). These results demonstrate that PHT4;4-mediated AsA transport to the chloroplast regulates the expression PRGs in −Fe+P through stress-induced ROS in the chloroplast that is perceived by bZIP58.

a Relative mRNA abundance of bZIP58 in shoots of plants expressing JUNGBRUNNEN1p::bZIP58 in the bzip58 (Line A1) and bzip58xpht4;4 (Line B1) mutant backgrounds. Plants were grown in the presence of iron and phosphorus (+Fe+P) for 7 days and transferred to +Fe+P, −Fe+P, or −Fe+P+AsA for 76 h. Data shown are the means from five biologically independent samples. Error bars indicate 95% confidence intervals. Asterisks indicate statistically different bZIP58 expression in the treatment compared to the control (+Fe+P) (P value < 0.01, two-sided Student’s t test). b Representative images of wild-type Col-0, Line A1 and Line B1 grown for 1 week under +Fe+P and transferred to three different media conditions (+Fe+P, −Fe+P, or −Fe+P+AsA) for 2 additional weeks. Scale bars: 7 mm. The experiment was repeated with similar results three times. c Total chlorophyll content in Col-0, Line A1 and Line B1 grown under +Fe+P, −Fe+P, or −Fe+P+AsA. FW: fresh weight. Data shown are from ten plants representing three independent experiments. Center lines show the medians; box limits indicate the 25th and 75th percentiles; whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles. d Relative mRNA abundance of PRGs genes (PSBTN (AT3G21055), AT1G5140, and PTAC16 (AT3G46780)) in shoots of Col-0, bzip58, Line A1, bzip58 pht4;4, and Line B1. Plants were grown under +Fe+P for 7 days and transferred to +Fe+P, −Fe+P, or −Fe+P+AsA for 76 h. Data shown are the means from five biologically independent samples. Error bars indicate 95% confidence intervals. For c, d, letters above bars or boxes represent statistically different means at P < 0.05 (one-way ANOVA with a Duncan post hoc test).
Discussion
Fe is an important micronutrient that plays crucial roles in plant growth and development. In nature, plants may experience Fe deficiency due to low Fe supply and alkaline pH (e.g., pH greater than 7.0 makes Fe unavailable to plants)25,26. Fe deficiency causes chlorosis27,28 and affects root growth29. Great progress has been achieved in understanding how plants respond to Fe deficiency, particularly the molecular mechanisms for Fe uptake in the roots and its distribution throughout the plant30. However, how plants sense and respond to Fe deficiency to control photosynthesis remains poorly understood. Fe is an essential cofactor in photosystem complexes and is present in almost all the components of the electron transport chain in the chloroplast6. Here, we describe a signaling pathway orchestrating communication between the chloroplast and the nucleus to control the expression of photosynthesis-related genes (PRGs) and chlorosis in Fe-deficient environments (Fig. 6). Modulation of this newly discovered pathway could have a direct impact on plant growth in the field by improving plant photosynthetic activity while reducing nutrient supply.

Fe deficiency (−Fe+P) causes a decrease in the expression of bZIP58 that is central to controlling the transcription of nuclear-encoded photosynthetic genes. P limitation under Fe deficiency (−Fe−P) prevents this downregulation of bZIP58 and induces VTC4. The induction of VTC4 expression requires bZIP58, whose effect could be direct or indirect, represented here by “X”. We propose that induction of VTC4 increases ascorbic acid in the chloroplast mediated by PHT4;4. We hypothesize that the increase of ascorbic acid level prevents ROS accumulation, thus maintaining the expression of bZIP58 and its downstream photosynthesis genes and leading to the “stay-green” phenotype. The figure was created with BioRender.com.
Generally, it has been assumed that chlorosis in Fe-deficient environments is due to Fe deficiency in leaves26. However, optimal photosynthesis in plants subjected to simultaneous Fe and P deficiency, as described in this study, challenges this assumption. There are two possibilities to explain the absence of chlorosis under −Fe−P conditions. First, Fe and P interactions can influence their bioavailability, and a joint deficiency of Fe and P may increase bioavailable Fe for photosynthesis. However, we did not find evidence to suggest that levels of bioavailable Fe change in the leaves of plants grown under −Fe−P using our methods; more sensitive methods of quantification with higher spatial resolution would be needed to further reinforce our conclusion. A second possibility is that P availability modulates Fe-deficiency signaling pathways that control photosynthesis. This hypothesis is clearly supported by our global gene expression analysis, which revealed that the downregulation of 32 PRGs by −Fe is prevented by simultaneous Fe and P deficiency. A similar transcriptional response was observed for bZIP58, a key transcriptional regulator of PRG expression we discovered in this study.
Fe and P are known to interact to regulate root growth13,31. However, our understanding of the molecular basis of Fe and P signaling crosstalk to regulate photosynthesis remains fragmentary. Therefore, identifying key molecular mechanisms improving plant photosynthesis under nutrient-limited conditions is of great importance. Using expression GWAS, we found that a chloroplastic Pi and AsA transporter PHT4;418 controls Fe-deficiency-induced chlorosis. It is noteworthy that our results show that PHT4;4 expression is not affected by Fe deficiency (Supplementary Fig. 10A). Given its dual-transport activity, one could ask which target of transport, Pi or AsA, is the cause of the persistent chlorotic phenotype in the pht4;4 mutant under the −Fe−P treatment. As far as Pi transport is concerned, our results show that genetic inactivation of other plastid Pi transporters (PHT4;3, PHT4;5, and PHT4;6)32 or key Pi signaling pathway components (SPX1 and PHR1)33 respond similarly to wild-type plants under −Fe+P or −Fe−P conditions. Our findings thus support the idea that PHT4;4 affects photosynthesis through its AsA transport activity. In line with this, AsA supplementation fully prevents the development of chlorosis in wild-type plants and not in pht4;4 plants, indicating that this phenotype depends on PHT4;4-mediated AsA transport to the chloroplast.
In our study, we measured AsA content 52 h after Fe-deficiency treatment, and we found a significant decrease in AsA accumulation in leaves. AsA levels have been shown to increase in response to 2–3 weeks Fe-deficiency treatments in Brassica napus leaves and sugar beet roots34,35. However, 3–7 days Fe-deficiency treatment in Arabidopsis exhibited a slight reduction in AsA accumulation in shoots36. It is intriguing to find a consistent reduction of AsA during the early phase of Fe-deficiency. One possibility is that AsA reduction could be one of the earliest signs of Fe-deficiency stress. Supporting this hypothesis is a recent finding that an early response to salt stress is repression of AsA biosynthesis by ABI4-mediated repression of VTC237. Identifying the upstream factors of the signaling pathway we discovered, including AsA biosynthesis, will be an important future direction.
Fe deficiency causes a decrease in electron flux in the thylakoid membranes38, and leads to an increase of ROS compared to +Fe conditions39,40. Several abiotic stresses enhance ROS accumulation in the chloroplast, a major site of ROS production22,41,42. Recently, ROS was proposed to act as a retrograde signal to influence plant responses to abiotic stresses, particularly for plant tolerance to salt stress43. This raises an exciting question of whether chloroplastic ROS could act as a retrograde signal under Fe deficiency to regulate the expression of nuclear PRGs. Our study indeed shows that, under −Fe conditions, ROS acts as a plastidic signal to influence the expression of PRGs and chlorophyll content via the repression of bZIP58. This effect becomes clearer when we express bZIP58 under the control of the ROS-inducible promoter of JUNGBRUNNEN1 (JUB1)24. The induction of JUB1p::bZIP58 by −Fe+P enhanced chlorophyll accumulation and expression of PRGs in the bzip58 mutant, and these responses were exacerbated in bzip58xpht4;4 double mutant. These new results demonstrate that Fe-deficiency-based chlorosis and PRG repression is mediated by chloroplastic ROS controlling the expression of the nuclear transcription factor bZIP58.
Taken together, this study identifies and validates a signaling pathway involved in the regulation of photosynthesis under combined Fe and P stresses (Fig. 6). This newly identified mechanism includes genes encoding chloroplastic (PHT4;4) and nuclear (bZIP58) proteins that prevent the repression of core photosynthesis genes and associated chlorosis under Fe and P co-deficiency. Furthermore, we provide evidence that bZIP58 regulates the expression of AsA biosynthesis genes and, using pht4;4 mutant plants, we reveal that AsA transport into the chloroplast is important for preventing chlorosis under Fe and P co-deficiency. We also show that in −Fe conditions, the increase of plastidic ROS levels is perceived at the nucleus as a signal to downregulate the expression of bZIP58 and downstream photosynthesis genes, which results in chlorosis. These results provide fundamental new insights into chlorophyll accumulation and photosynthesis under Fe limitation and identify a signaling pathway that may coordinate plastid–nuclear communication as a means to adapt photosynthesis to nutrient availability.
Methods
Plants and growth conditions
Seeds of Arabidopsis thaliana wild-type (ecotype Columbia, Col-0, CS60000) and knockout mutant lines SALK_139877 (AT1G13570), SALK_150849 (AT1G13580), SALK_063177 (AT1G13590), N571881 (AT1G13600), SALK_087271 (AT1G13605), SALK_130208 (AT1G13607), SALK_023173 (AT1G13608), SAIL_1243_E04 (AT1G13609), SAIL_897_D11 (AT1G13610), N469134 (At4g00355), SALK_128714 (AT4G00360), N469134 (AT4G00370), SAIL_842_E09 (AT4G00380), N866595 (At4g00390), SAIL_633_E10 (At4g00400), SALK_077222 (AT3G02870) SALK_053900 (AT3G46980), SALK_114708 (AT5G20380), SAIL_809_B01 (AT5G44370), SALK_067629 (AT4G28610), and SALK_039445 (AT5G20150) were obtained from the Nottingham Arabidopsis Stock Centre (NASC). Homozygous mutant lines were confirmed by PCR using the primers listed in Supplementary Table 1. bZIP58 complemented lines (bZIP58-CL) were generated by expressing 3896 bp genomic DNA containing bZIP58 in the bzip58 mutant background (NASC, N571881). Complementation of pht4;4 mutant plants (PHT4;4-CL) was obtained by expressing 6450 bp genomic DNA containing PHT4;4 in the pht4;4 mutant background (NASC, N469134). The bzip58xpht4;4 double mutant was obtained through the genetic crossing. Arabidopsis plants were grown on control (+Fe+P) plates containing 1.249 mM KH2PO4; 0.25 mM Ca(NO3)2; 0.5 mM KNO3; 1 mM MgSO4; 100 μM FeSO4.7H2O; 30 μM H3BO3; 1 μM ZnCl2; 10 μM MnCl2; 1 μM CuCl2; 0.1 μM (NH4)6Mo7O24; and 50 μM KCl; 0.05% 2-(N-morpholino)ethanesulfonic acid (MES), without sucrose supplementation, and 0.8% washed agar. The agar was washed three times with 50 mM EDTA, pH 5.7, with continuous stirring for 16 h, then washed 6 times with Milli-Q de-ionized water for 2 h to reduce mineral contamination44. P-deficient media contained 12.49 µM KH2PO4 (+Fe−P). Fe-free media was obtained by omitting FeSO4.7H2O from the growth media (−Fe+P). P- and Fe-deficient media contained 12.49 µM KH2PO4 (+Fe−P), and no FeSO4.7H2O (−Fe−P). Seeds were stratified at 4 °C for 3 days and grown on vertical agar plates in a growth chamber with 22 °C, 24 h of light at 100 µmol m−2 s−1 fluorescent illumination. Lemna gibba (duckweed) plants used during this study were obtained from Duckweeds stock center (stock number 29-DWC131) at Rutgers University (USA). Duckweed plants were grown in 1× Schenk & Hildebrandt (SH) hydroponic medium containing 0.05% 2-(N-morpholino)ethanesulfonic acid (MES) and 1% sucrose, and pH adjusted to 5.7. For experiments with duckweeds, P-deficient and Fe-deficient media contained 1% NH4H2PO4 and 1% FeSO47H2O, respectively, of 1X SH media. Media were changed every 7 days. The growth condition was 22 °C and 24 h of light at 80 µmol m−2 s−1. Rice (Oryza sativa cv Nipponbare) plants were grown hydroponically in 0.25X Yoshida media45 under light/dark cycle of 14/10 h, and temperature of 28/25 °C. Single (−P or −Fe) and combined (−P−Fe) nutrient deficiency stresses were applied to 10 day-old plants. NaH2PO4 (0.33 mM) and Fe-NaEDTA (0.04 mM) present in the complete media were omitted in the P- and/or Fe-deficient media.
Plasmid construction and plant transformation
The JUNGBRUNNEN1 (JUB1)24 promoter and The bZIP58 coding region were amplified using PCR using primers listed in Supplementary Table 1. The PCR product was and then cloned into the binary vector pCAMBIA1301 using PstI and BamHI restriction enzymes. The PstI cut site was used for the insertion of the JUB1 promoter upstream of the bZIP58 coding region. The constructs were transformed into Agrobacterium tumefaciens strain GV3101 and then used for Arabidopsis transformation via the floral dip method46. Transgenic plants were selected by antibiotic resistance, and homozygous descendants of hemizygous T2 plants segregating 3:1 for antibiotic resistance:sensitivity were used for analysis.
Iron concentration measurement
Arabidopsis seeds were germinated and grown in the control (+Fe+P) media for 7 days, and then transferred to +Fe+P, iron-deficient (−Fe+P), phosphate-deficient (+Fe−P), or iron and phosphate-deficient (−Fe−P) conditions and grown for 7 additional days. Plants were harvested and shoot samples were dried at 70 °C for 3 days. Total iron was extracted by acid digestion in 1N nitric acid using MARSX (CEM) microwave digester. A 1:10 dilution of the digested material was used to quantify total iron with inductively coupled plasma atomic emission spectrometry (ICP-OES).
Analysis of photosystem II activity
Photosystem II (PSII) activity was defined as the maximum quantum yield of the primary quinone acceptor PSII, which was estimated by the ratio of variable fluorescence (Fv) and maximal fluorescence (Fm) of the chlorophyll, Fv/Fm47. Arabidopsis wild-type (Col-0) seeds were germinated and grown in control (+Fe+P) for 7 days then transferred to three different media: +Fe+P, iron-deficient (−Fe+P), and iron- and phosphate-deficient (−Fe−P) conditions for 0 h (time of the transfer), 15 h, 28 h, 39 h, 52 h, 76 h, 100 h, 124 h, 148 h, and 172 h. Plates containing the seedlings were dark-adapted for 30 min followed by a very short (160 µs) exposure to a blue measuring beam to determine the minimal fluorescence (F0). The intensity of the detecting and the continuous illumination used was of 156 µE m−2 s−1. A saturating light flash (2600 µE m−2 s−1, 250 ms) was applied to measure the maximum fluorescence (Fm). Kinetics were normalized to the maximum fluorescence (Fm). The maximum quantum yield of Photosystem II (Fv/Fm = (Fm – F0)/Fm) was measured for each growth condition, and non-photochemical quenching was calculated using (Fm/Fm’) − 147.
Chlorophyll content measurement
Seeds of Arabidopsis genotypes were germinated and grown in control (+Fe+P) media for 7 days then transferred to three different media: +Fe+P, iron-deficient (−Fe+P), and iron- and phosphate-deficient (−Fe−P) conditions. Fresh leaves (~30 mg) were incubated in 2.5 mL of 80% acetone overnight in the dark at 4 °C. Total chlorophyll content was measured using a UV–VIS spectrophotometer (Beckman Coulter, DU 530). The absorbance of the supernatant was measured at 645 and 633 nm. The concentration of total chlorophyll was calculated using the following equation48: 20.31 A645 + 8.05 A663/FW [μg g−1] (FW: fresh weight of tissue in grams).
Ascorbic acid content determination
Seeds of Arabidopsis genotypes were germinated and grown in control (+Fe+P) media and then transferred to +Fe+P, −Fe+P, or −Fe−P media for 76 h. Ascorbic acid (AsA) content was measured by a colorimetric assay49. Briefly, shoots were collected and homogenized in ice-cold 6% trichloroacetic acid (TCA) (Sigma-Aldrich). The homogenate was centrifuged at 6000 rpm for 25 min at 4 °C, and the supernatant was collected. In the supernatant, Fe3+ (ferric ion) is reduced by AsA to the Fe2+ (ferrous ion) that, when coupled with 2,2-dipyridyl, forms a complex with a characteristic absorbance at 525 nm49. A standard curve was generated using known concentrations of AsA made in 6% TCA to determine the AsA concentration. Blanks were prepared using only 6% TCA. AsA concentration was expressed as µmol g−1 fresh weight.
Hydrogen peroxide quantification
Seeds of Arabidopsis genotypes were germinated and grown in control (+Fe+P) media and then transferred to +Fe+P, −Fe+P, or −Fe−P media for 76 h. Hydrogen peroxide (H2O2) (Sigma-Aldrich) was quantified as described previously50,51. Fresh shoot tissues (0.2 g) were homogenized with 0.1% (w/v) TCA and were centrifuged at 12,000× g for 15 min at 4 °C. In total, 0.5 ml of supernatant was added to 0.5 ml of 10 mM potassium phosphate buffer (pH 7.0) and 1 ml of 1 M potassium iodide. The absorbance of the reaction mixture was measured at 390 nm. The amount of H2O2 was calculated using a standard curve prepared from known concentrations of H2O2 ranging from 0.1 to 1 mM.
RNA sequencing and analysis
Arabidopsis wild-type (Col-0) plants were grown in control (+Fe+P) media for 7 days and transferred to three different media: control (+Fe+P), iron deficiency (−Fe+P), and iron and phosphate deficiency (−Fe−P) conditions. Shoots were collected at 39, 52, and 76 h after the transfer. For RNA-seq experiments, three biological replicates were prepared for each time point (39, 52, and 76 h) and each condition (+Fe+P, −Fe+P, and −Fe−P) for a total of 27 samples. Total RNA was extracted from these samples using RNeasy Plant Mini Kit (QIAGEN) using the RLT buffer supplemented with 2-mercaptoethanol, and RNA quality was verified using an Agilent 2100 BioAnalyzer. The mRNAs were subsequently isolated using magnetic KAPA Biosystems oligo-dT beads from KAPA Biosystems (Roche) and then used for library construction using the KAPA Biosystems RNA HyperPrep Kit (Roche). To index the libraries, we used adapters from the KAPA Biosystems Single-Indexed Adapter Set A + B (Roche). Before pooling the libraries, we monitored their quality and concentrations using an Agilent 2100 BioAnalyzer, Qubit dsDNA HS Assay Kit (Thermo Fisher Scientific), and the KAPA Library Quantification Kit (Roche). Pooled libraries were then sequenced using the NextSeq 500 System at the Stanford Functional Genomics Facility (Stanford, CA). Raw reads were demultiplexed and aligned to the TAIR10 genome annotation using HISAT252 on the Galaxy web platform53. Finally, mapped read counts were used to perform normalization and differential expression analysis on R using the DESeq254 and TxDB.Athaliana.BioMart.plantsmart27 (Bioconductor) packages. In DESeq2, P values from the Wald test were corrected for multiple hypothesis testing using the Benjamini and Hochberg method. A transcript was considered differentially expressed if the adjusted P value <0.05. Volcano plots were generated using the EnhancedVolcano package (version 1.6.0) (Bioconductor) with a default cut-off of log2(fold change) >|2| and adjusted P value <10e−6. DEGs having a P value of 0 were converted to 10−1 × lowest nonzero P value.
Turnbull staining
Arabidopsis plants (Col-0 genotype) were grown on 0.5× MS medium. Fourteen days after germination, plants were vacuum infiltrated with Turnbull staining solution (4% (v/v) HCl and 4% (w/v) K-ferricyanide) for 15 min and then incubated for 45 min at room temperature on an orbital shaker55,56. To inhibit peroxidases and redox reactions, the plants were washed with distilled water and treated for 1 h in methanol containing 0.065% (w/v) sodium azide and 0.3% (v/v) H2O2. After washing with 0.1 M potassium phosphate buffer (pH 7.4) followed by washing with sterile water, the plants were placed on a slide and observed under a microscope (Olympus BX61).
Real-time quantitative reverse-transcription PCR
Seeds of Arabidopsis wild type (Col-0), bzip58, and pht4;4 mutant plants were germinated and grown for 7 days in control (+Fe+P) media, and then transferred to +Fe+P, Fe+P, or −Fe−P. Shoot tissues were collected at 76 h after the transfer and then used for total RNA extraction as described in ref. 57. Each experiment was conducted with 16 plants, and 4–6 plants were pooled for RNA extraction, resulting in 3–4 biological replicates. Two micrograms of the total RNA were used for reverse transcription (Promega) to synthesize cDNA using oligo(dT) primer (Promega). Real-time quantitative reverse-transcription PCR (qRT-PCR) was performed as described in ref. 57 using 384-well plates with a LightCycler 480 Real-Time PCR System (Roche diagnostics). The Ubiquitin 10 mRNA (UBQ10: At4g05320) was used as a control to calculate the relative mRNA level of each gene. The primers used in this study are listed in Supplementary Table 1.
Genome-wide association studies (GWAS)
Gene expression data of the 32 genes that were specifically downregulated by −Fe+P but not by −Fe−P relative to +Fe+P were downloaded from leaf expression data of 727 Arabidopsis accessions17. Normalized RNA-seq read counts of these genes were used to perform principal component analysis, and contributions of the accessions to PC1 that explained 89.5% of the expression variance of the 32 genes were used to run genome-wide association (GWA) analysis. GWA mapping was performed using 1001 genomes SNP data58 as implemented in the web application GWAPP59. Bonferroni correction (α = 0.05) and false-discovery rate (FDR) at 5%60 were implemented to account for multiple hypothesis tests.
In planta transactivation assay
The in planta transactivation assay61 was performed in Nicotiana benthamiana. Promoters of PHOTOSYSTEM II SUBUNIT T (PSBTN); PHOTOSYSTEM II 5 kD, PLASTID TRANSCRIPTIONALLY ACTIVE 16 (PTAC16), LIGHT HARVESTING COMPLEX PHOTOSYSTEM II (LHCB4.1), PHOTOSYSTEM II REACTION CENTER W (PSBW) were fused to the reporter gene encoding β-glucuronidase (GUS) using Gateway cloning. The coding sequence of bZIP58 was inserted downstream of the CaMV 35S promoter (35S::bZIP58). The 35S::C-YFP construct was used as a negative effector control61. Each construct was transformed into A. tumefaciens. Cultures inoculated with positive clones were grown overnight at 28 °C, pelleted through centrifugation, and washed four times with infiltration buffer (10 mM MgCl2, 10 mM MES (pH 5.6), and 100 μM acetosyringone). Fully expanded leaves of 5- to 6-week-old tobacco plants were co-infiltrated with the effector construct and reporter construct (OD600 of 0.8). One leaf per plant was infiltrated for each plasmid combination and three independent infiltrations per combination were performed. Leaves were collected 3 days after infiltration and the infiltrated areas in each leaf were excised and pooled into one sample. To measure GUS enzymatic activity, 100 μg of total protein extract was incubated at 37 °C in GUS assay solution (2 mM 4-methylumbelliferyl β-d-glucuronide (Gold Biotechnology); 100 μl of the reaction was transferred to 1.9 ml of 0.2 M carbonate (Na2CO3) stop solution at the 0, 30, 60, 90, and 120 min time points. GUS activity was measured using the Dyna Quant 200 fluorometer (Hoefer) and activity values were calculated following the β-glucuronidase (GUS) Fluorescent Activity Detection Kit (Sigma-Aldrich) protocols. For each promoter, the effect of bZIP58 on GUS activity was normalized to the effect of C-YFP on GUS activity to obtain a relative GUS enzymatic activity measurement. The primers used to clone the promoters of the photosynthesis-related genes are listed in Supplementary Table 1.
Statistical analysis
Box plots were generated using a web-based application “BoxPlotR”62. Statistical analyses of the data were performed using analysis of variance (ANOVA). One-way ANOVA with a Duncan post hoc test and two-way analysis of variance (ANOVA) and Tukey’s honest significant difference (HSD) test were used to compare mean values. For all the statistical analyses, the difference was considered statistically significant when the test yielded a P value <0.05.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Data supporting the findings of this work are available within the paper and its Supplementary Information and Source Data files. The datasets and plant materials generated and analyzed during this study are available from the corresponding author H.R. upon request. Transcriptome data were deposited in NCBI’s Gene Expression Omnibus (GEO) under the project number GSE163190. Source data are provided with this paper.
References
Myouga, F. et al. The chloroplast function database: a large-scale collection of Arabidopsis Ds/Spm-or T-DNA-tagged homozygous lines for nuclear-encoded chloroplast proteins, and their systematic phenotype analysis. Plant J. 61, 529–542 (2010).
Chen, Y. & Barak, P. Iron nutrition of plants in calcareous soils. Adv. Agron. 35, 217–240 (1982).
Carstensen, A. et al. The impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant Physiol. 177, 271–284 (2018).
Terry, N. & Abadía, J. Function of iron in chloroplasts. J. Plant Nutr. 9, 609–646 (1986).
Terry, N. & Low, G. Leaf chlorophyll content and its relation to the intracellular localization of iron. J. Plant Nutr. 5, 301–310 (1982).
Lill, R. Function and biogenesis of iron–sulphur proteins. Nature 460, 831 (2009).
Briat, J.-F., Dubos, C. & Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 20, 33–40 (2015).
Balk, J. & Pilon, M. Ancient and essential: the assembly of iron–sulfur clusters in plants. Trends Plant Sci. 16, 218–226 (2011).
Marschner, H. Mineral Nutrition of Higher Plants. 2nd edn. (Academic Press, Elsevier, 1995).
DeKock, P. C., Hall, A. & Inkson, R. H. E. Active iron in plant leaves. Ann. Bot. 43, 737–740 (1979).
Saenchai, C. et al. The involvement of OsPHO1; 1 in the regulation of iron transport through integration of phosphate and zinc deficiency signaling. Front. Plant Sci. 7, 396 (2016).
Meguro, R. et al. Nonheme-iron histochemistry for light and electron microscopy: a historical, theoretical and technical review. Arch. Histol. Cytol. 70, 1–19 (2007).
Müller, J. et al. Iron-dependent callose deposition adjusts root meristem maintenance to phosphate availability. Dev. Cell 33, 216–230 (2015).
Cassin, G., Mari, S., Curie, C., Briat, J.-F. & Czernic, P. Increased sensitivity to iron deficiency in Arabidopsis thaliana overaccumulating nicotianamine. J. Exp. Bot. 60, 1249–1259 (2009).
Bertamini, M., Nedunchezhian, N. & Borghi, B. Effect of iron deficiency induced changes on photosynthetic pigments, ribulose-1, 5-bisphosphate carboxylase, and photosystem activities in field grown grapevine (Vitis vinifera L. cv. Pinot noir) leaves. Photosynthetica 39, 59–65 (2001).
Liu, W. et al. The ethylene response factor AtERF4 negatively regulates the iron deficiency response in Arabidopsis thaliana. PLoS ONE 12, e0186580 (2017).
Kawakatsu, T. et al. Epigenomic diversity in a global collection of Arabidopsis thaliana accessions. Cell 166, 492–505 (2016).
Miyaji, T. et al. AtPHT4; 4 is a chloroplast-localized ascorbate transporter in Arabidopsis. Nat. Commun. 6, 5928 (2015).
Torabinejad, J., Donahue, J. L., Gunesekera, B. N., Allen-Daniels, M. J. & Gillaspy, G. E. VTC4 is a bifunctional enzyme that affects myoinositol and ascorbate biosynthesis in plants. Plant Physiol. 150, 951–961 (2009).
Noctor, G. & Foyer, C. H. Ascorbate and glutathione: keeping active oxygen under control. Annu. Rev. Plant Biol. 49, 249–279 (1998).
Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 194, 7–15 (2011).
Shapiguzov, A., Vainonen, J., Wrzaczek, M. & Kangasjärvi, J. ROS-talk–how the apoplast, the chloroplast, and the nucleus get the message through. Front. Plant Sci. 3, 292 (2012).
Shaikhali, J. & Wingsle, G. Redox-regulated transcription in plants: emerging concepts. AIMS Mol. Sci. 4, 301 (2017).
Wu, A. et al. JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 24, 482–506 (2012).
Anderson, W. B. Diagnosis and correction of iron deficiency in field crops‐an overview. J. Plant Nutr. 5, 785–795 (1982).
Korcak, R. F. Iron deficiency chlorosis. Hort. Rev. 9, 133–186 (1987).
Hansen, N. C., Jolley, V. D., Naeve, S. L. & Goos, R. J. Iron deficiency of soybean in the North Central US and associated soil properties. Soil Sci. Plant Nutr. 50, 983–987 (2004).
Marsh, H. V. Jr, Evans, H. J. & Matrone, G. Investigations of the role of iron in chlorophyll metabolism. II. Effect of iron deficiency on chlorophyll synthesis. Plant Physiol. 38, 638 (1963).
Gruber, B. D., Giehl, R. F. H., Friedel, S. & von Wirén, N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 163, 161–179 (2013).
Kobayashi, T. & Nishizawa, N. K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 63, 131–152 (2012).
Ward, J. T., Lahner, B., Yakubova, E., Salt, D. E. & Raghothama, K. G. The effect of iron on the primary root elongation of Arabidopsis during phosphate deficiency. Plant Physiol. 147, 1181–1191 (2008).
Guo, B. et al. Functional analysis of the Arabidopsis PHT4 family of intracellular phosphate transporters. N. Phytologist 177, 889–898 (2008).
Puga, M. I. et al. SPX1 is a phosphate-dependent inhibitor of phosphate starvation response 1 in Arabidopsis. Proc. Natl Acad. Sci. 111, 14947–14952 (2014).
Tewari, R. K., Hadacek, F., Sassmann, S. & Lang, I. Iron deprivation-induced reactive oxygen species generation leads to non-autolytic PCD in Brassica napus leaves. Environ. Exp. Bot. 91, 74–83 (2013).
Zaharieva, T. B. & Abadía, J. Iron deficiency enhances the levels of ascorbate, glutathione, and related enzymes in sugar beet roots. Protoplasma 221, 269–275 (2003).
Ramírez, L., Bartoli, C. G. & Lamattina, L. Glutathione and ascorbic acid protect Arabidopsis plants against detrimental effects of iron deficiency. J. Exp. Bot. 64, 3169–3178 (2013).
Kakan, X. et al. Ascorbic acid modulation by ABI4 transcriptional repression of VTC2 in the salt tolerance of Arabidopsis. BMC Plant Biol. 21, 1–12 (2021).
Yabuta, Y. et al. Light regulation of ascorbate biosynthesis is dependent on the photosynthetic electron transport chain but independent of sugars in Arabidopsis. J. Exp. Bot. 58, 2661–2671 (2007).
Roncel, M. et al. Iron deficiency induces a partial inhibition of the photosynthetic electron transport and a high sensitivity to light in the diatom Phaeodactylum tricornutum. Front. Plant Sci. 7, 1050 (2016).
Zhai, L. et al. At ROP 6 is involved in reactive oxygen species signaling in response to iron-deficiency stress in Arabidopsis thaliana. FEBS Lett. 592, 3446–3459 (2018).
Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 141, 391–396 (2006).
Lodeyro, A. F. et al. Suppression of reactive oxygen species accumulation in chloroplasts prevents leaf damage but not growth arrest in salt-stressed tobacco plants. PLoS ONE 11, e0159588 (2016).
Zhuang, Y. et al. EGY3 mediates chloroplastic ROS homeostasis and promotes retrograde signaling in response to salt stress in Arabidopsis. Cell Rep. 36, 109384 (2021).
Haydon, M. J. et al. Vacuolar nicotianamine has critical and distinct roles under iron deficiency and for zinc sequestration in Arabidopsis. Plant Cell 24, 724–737 (2012).
Yoshida, S., Forno, D. A. & Cock, J. H. Laboratory Manual for Physiological Studies of Rice. (International Rice Research Institute, 1976).
Clough, S. J. & Bent, A. F. Floral dip: a simplified method for Agrobacterium‐mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743 (1998).
Baker, N. R. Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 59, 89–113 (2008).
Zhang, J., Han, C. & Liu, Z. Absorption spectrum estimating rice chlorophyll concentration: preliminary investigations. J. Plant Breed. Crop Sci. 1, 223–229 (2009).
Gillespie, K. M. & Ainsworth, E. A. Measurement of reduced, oxidized and total ascorbate content in plants. Nat. Protoc. 2, 871–874 (2007).
Alexieva, V., Sergiev, I., Mapelli, S. & Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 24, 1337–1344 (2001).
Gechev, T., Mehterov, N., Denev, I. & Hille, J. A simple and powerful approach for isolation of Arabidopsis mutants with increased tolerance to H2O2-induced cell death. Methods Enzymol. 527, 203–220 (2013).
Kim, D., Langmead, B. & Salzberg, S. L. HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12, 357–360 (2015).
Afgan, E. et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 46, W537–W544 (2018).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Roschzttardtz, H., Conéjéro, G., Curie, C. & Mari, S. Identification of the endodermal vacuole as the iron storage compartment in the Arabidopsis embryo. Plant Physiol. 151, 1329–1338 (2009).
Meguro, R. et al. The presence of ferric and ferrous iron in the nonheme iron store of resident macrophages in different tissues and organs: histochemical demonstrations by the perfusion-Perls and-Turnbull methods in the rat. Arch. Histol. Cytol. 68, 171–183 (2005).
Rouached, H. et al. Differential regulation of the expression of two high-affinity sulfate transporters, SULTR1. 1 and SULTR1. 2, in Arabidopsis. Plant Physiol. 147, 897–911 (2008).
The 1001 Genomes Consortium. 1,135 genomes reveal the global pattern of polymorphism in Arabidopsis thaliana. Cell 166, 481–491 (2016).
Seren, Ü. et al. GWAPP: a web application for genome-wide association mapping in Arabidopsis. Plant Cell 24, 4793–4805 (2012).
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. 57, 289–300 (1995).
Bossi, F. et al. Systematic discovery of novel eukaryotic transcriptional regulators using sequence homology independent prediction. BMC Genomics 18, 1–20 (2017).
Spitzer, M., Wildenhain, J., Rappsilber, J. & Tyers, M. BoxPlotR: a web tool for generation of box plots. Nat. Methods 11, 121 (2014).
Acknowledgements
The authors thank the members of the Rhee lab; Benoit Lacombe and HONUDE team (INRAe) for their comments on the manuscript and helpful discussions. The authors are grateful to Jean-Francois Briat, Stéphane Mari and Diego Melo-Almeida for helpful discussion on Fe staining in Arabidopsis leaves. We thank the ICP-MS/TIMS Facility within Stanford University for assistance with the ICP-MS measurements, and the Stanford Functional Genomics Facility for assistance with RNA sequencing (Stanford, CA). The GWAS analysis was made possible using expression data generated by Kawakatsu et al.17. This work was funded in part by the “Institut National de la Recherche Agronomique—Montpellier—France” INRA, the AgreeenSkills Plus, and Michigan State University (USA) to H.R. as well as by the Carnegie Institution for Science, Brigitte Berthelemot, National Science Foundation (IOS-1546838, IOS-1026003), and the U.S. Department of Energy, Office of Science, Office of Biological and Environmental Research, Genomic Science Program grant nos. DE-SC0018277, DE-SC0008769, and DE-SC0020366 to S.Y.R. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
S.Y.R. and H.R. conceived the project. Experiments were designed by S.Y.R., H.R., H.N., and Z.S. and mainly carried out by H.N. S.C. performed and analyzed experiments related to photosystem II activity. bZIP58-GFP localization, ascorbic acid quantification, and hydrogen peroxide assays were conducted by H.C. and N.B. RNA-seq data were generated and analyzed by Y.D. Gene Ontology analysis was performed by K.Z. Z.S. performed the genome-wide association mapping. H.R. and K. L.P. performed the qRT-PCR analyses, generated plasmid constructs, the homozygote mutants, and the complemented mutant lines. S.Y.R., H.R., and Z.S. wrote the paper with input from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nam, HI., Shahzad, Z., Dorone, Y. et al. Interdependent iron and phosphorus availability controls photosynthesis through retrograde signaling. Nat Commun 12, 7211 (2021). https://doi.org/10.1038/s41467-021-27548-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-021-27548-2
This article is cited by
-
Transcriptome analysis of Pennisetum americanum × Pennisetum purpureum and Pennisetum americanum leaves in response to high-phosphorus stress
BMC Plant Biology (2024)
-
Phosphorus deficiency alleviates iron limitation in Synechocystis cyanobacteria through direct PhoB-mediated gene regulation
Nature Communications (2024)
-
Recent advances in research on phosphate starvation signaling in plants
Journal of Plant Research (2024)
-
Exploring the interplay between angiosperm chlorophyll metabolism and environmental factors
Planta (2024)
-
Molecular characterization and expression pattern of Rubisco activase gene GhRCAβ2 in upland cotton (Gossypium hirsutum L.)
Genes & Genomics (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
