
Abstract
Despite considerable advances in knowledge of the anatomy, ecology and evolution of early mammals, far less is known about their physiology. Evidence is contradictory concerning the timing and fossil groups in which mammalian endothermy arose. To determine the state of metabolic evolution in two of the earliest stem-mammals, the Early Jurassic Morganucodon and Kuehneotherium, we use separate proxies for basal and maximum metabolic rate. Here we report, using synchrotron X-ray tomographic imaging of incremental tooth cementum, that they had maximum lifespans considerably longer than comparably sized living mammals, but similar to those of reptiles, and so they likely had reptilian-level basal metabolic rates. Measurements of femoral nutrient foramina show Morganucodon had blood flow rates intermediate between living mammals and reptiles, suggesting maximum metabolic rates increased evolutionarily before basal metabolic rates. Stem mammals lacked the elevated endothermic metabolism of living mammals, highlighting the mosaic nature of mammalian physiological evolution.
Similar content being viewed by others
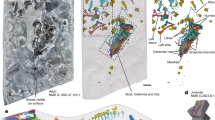


Introduction
Recent discoveries and analyses have revolutionized our knowledge of Mesozoic mammals, revealing novel aspects of their ecology1,2, development2,3, systematics2,3 and macroevolution4,5. However, details of physiology are more difficult to determine from fossils, and our knowledge of physiological evolution remains comparatively poor. Living mammals are endotherms, possessing the ability to control and maintain metabolically produced heat and have a substantially higher capacity for sustained aerobic activity than ectothermic animals6,7,8. The origin of endothermy is an important event in mammalian evolution, often noted as key to their success6,7,8. There are a number of competing evolutionary hypotheses for the origin of endothermy: (a) selection for higher maximum metabolic rates (MMRs) enhanced sustained aerobic activity6,9,10, (b) selection for higher basal metabolic rates (BMRs) enhanced thermoregulatory control11,12, or (c) MMRs and BMRs evolved in lockstep with each other7,8.
Direct evidence from living mammals to support these hypotheses is equivocal7. Recent analyses find no long-term evolutionary trend in BMR13 contradicting earlier suggestions of increasing BMR throughout the Cenozoic12 and so implying that the Middle Jurassic (∼170 Ma) most recent common ancestor (MRCA) of living mammals13 possessed a BMR within the range of present-day mammals. Several indirect indicators of metabolic physiology in fossil synapsids have been suggested but provide contradictory evidence for the timing of the origin of endothermy and its evolutionary tempo. These include: the presence of fibrolamellar long-bone histology14,15, first seen in the Early Permian synapsid Ophiacodon ∼300 million years (Ma) ago16; the presence of an infraorbital canal and lack of parietal foramen, used to infer facial whiskers, fur, lactation and endothermy in Early Triassic (∼245 Ma) cynodonts17; inferred maxillary nasal turbinates in the Late Permian (∼255 Ma) therapsid Glanosuchus, used to suggest that mammalian levels of endothermy evolved by the Late Triassic (∼210 Ma)18; a trend towards increased relative brain size initiated in Late Triassic non-mammaliaform cynodonts19 and the mammaliaform (stem mammal sensu Rowe20) Morganucodon21,22; and acquisition of a parasagittal gait in the Early Cretaceous (∼125 Ma) therian mammals Eomaia and Sinodelphys23. Several recent studies provide more quantitative links to physiological parameters. Oxygen isotopes were used to infer elevated thermometabolism in Middle–Late Permian (∼270–255 Ma) eucynodonts24, red blood cell size diminution in Late Permian (∼255 Ma) eutheriodontid therapsids was linked via two proxies to increased MMR25 and osteocyte lacuna shape correlations suggested “mammalian” resting metabolic rates in Permo-Triassic (∼250 Ma) dicynodonts26.
However, the inconsistency of these characters, in time and with respect to phylogeny27,28, along with re-assessments of function in relation to endothermy7,29,30, limit their use as conclusive indicators of modern mammalian levels of endothermy in fossil taxa. Such temporal and phylogenetic heterogeneity suggests that the evolution of mammalian endothermy followed a complex, mosaic pattern with different physiological aspects likely evolving independently, and at separate rates, towards current mammalian levels. Additionally, few of these physiological proxies are directly related to measurable aspects of metabolic rate.
To address these issues, we use two proxies to improve understanding of physiology at one of the most important nodes along this transition. We do so by estimating BMR and growth rate, and calculating a known proxy for MMR, for two of the earliest known mammaliaforms, Morganucodon and Kuehneotherium1,31. Using cementochronology to estimate maximum lifespan by counting growth increments in synchrotron radiation-based micro-computed tomographic (µCT) data of fossil dental cementum, we estimate that both taxa had significantly longer lifespans than extant mammals of comparable size. By regressing lifespan against BMR and postnatal growth rate in extant mammals and reptiles, we in turn estimate significantly lower values for both of these metrics for the earliest mammaliaforms. However, when we compare the blood flow index (the ratio between femoral nutrient foramina area and femur length that serves as a proxy for MMR) of Morganucodon with those of extant taxa, we find that Morganucodon had an intermediate value between living mammals and reptiles. These results suggest that basal mammaliaforms occupied a metabolic grade similar to living reptiles and had yet to achieve the endothermic physiology of living mammals.
Results
Lifespan as a proxy for mammaliaform physiology
We used maximum lifespan (i.e. the single longest known lifespan of a taxon) estimates for fossil mammaliaform taxa as a proxy for both BMR32 and postnatal growth rate33. In extant tetrapods, negative correlations exist between maximum lifespan and BMR32 and between maximum lifespan and growth rate33,34. In general, the longer a mammal’s lifespan, the lower its size-adjusted BMR and growth rate. Growth rates have been shown to correlate strongly with metabolic power in extant vertebrates, with endotherms growing an order of magnitude faster than ectotherms34,35. Maximum lifespan is an applicable value for fossil samples, as, unlike other metrics (e.g. 10% most long lived or mean lifespan of a cohort), it does not rely on cohort- or population-based statistics that fossil samples cannot fulfil36. This value is also less susceptible to extrinsic population-level factors on lifespan, such as disease or predation, and relates most closely to the physiological limit of lifespan of an organism. An accurate assessment of maximum lifespan in fossil mammals can therefore be used to estimate their metabolic potential.
To estimate mammaliaform lifespans, we used cementochronology. This well-established technique, which counts annual growth increments in tooth-root cementum, has been used to record lifespans in extant mammals37 with >70 species aged using this technique38. Cementum is a mineralized dental tissue surrounding the tooth root (Fig. 1a, b), attaching it to the periodontal ligament and anchoring the tooth within the alveolus37. Growth of cementum is continuous throughout life in extant mammals and seasonally appositional in nature, forming a series of increments of differing thickness and opacity when viewed in histological thin sections under light microscopy. The correlation between increment count and chronological age is well documented, with one thick and one thin increment deposited every year38. It has been shown that the thin, hyper-mineralized opaque increments record growth rate reduction in less favourable seasons39,40.

a, b Three-dimensional reconstructions of a Morganucodon right lower molar tooth NHMUK PV M 104134 (voxel size 2 μm, μCT) and b Kuehneotherium right lower molar tooth NHMUK PV M 21095 (voxel size 1.2 μm, PPC-SRµCT). Green = cementum. c, d Transverse PPC-SRµCT virtual thin sections (0.33 μm voxel size) of roots of c NHMUK PV M 104134 and d NHMUK M 27436. Red bracketed line highlights extent of cementum surrounding dentine. e, f Close-ups of boxes in c, d, with five and four circumferential light/dark increment pairs highlighted by red arrows, respectively. g Synchrotron nanotomographic virtual thin section of NHMUK PV M 104134 (30 nm voxel size) provides close-up of region close to box in e. Vertical red arrows = cementum increments; horizontal blue arrows, dashed blue lines and Sf = radial bands of Sharpey’s fibres; yellow dashed line and glT = granular layer of Tomes; green dashed line and hlH-S = hyaline layer of Hopewell-Smith. Scale bars represent 500 µm in a, b, 100 µm in c, d, 30 µm in e, f and 10 µm in g.
Despite this potential, cementochronology has not previously been attempted for fossil mammals older than the Pleistocene (2.6 Ma)41, because histological thin sections destroy fossils and provide only a restricted field of view. We overcame these problems by using propagation phase-contrast X-ray synchrotron radiation microtomography (PPC-SRµCT) to non-destructively image fossilized cementum increments42,43. The sub-micrometre resolution, fast-throughput and three-dimensional (3D) nature of PPC-SRµCT allows for large sample sizes and for increments to be imaged along their entire transverse and longitudinal trajectories in volumetric PPC-SRµCT data sets. Cementum increments are known to occasionally split and coalesce, creating errors in counts based on single, or limited numbers of, two-dimensional thin sections created for each tooth37,44 (Supplementary Fig. 1). PPC-SRµCT imaging and 3D segmentation of individual increments across extensive vertical distances within the cementum allowed us to confidently distinguish principal annual increments from any accessory increments created by lensing and coalescence (see “Methods”).
Morganucodon and Kuehneotherium are shrew-sized insectivores1, which co-existed on a small landmass during the Early Jurassic marine transgression (Hettangian-early Sinemurian, ∼200 Ma) in what is now Glamorgan, South Wales, UK45 (Supplementary Notes 1 and 2 and Supplementary Data 1 and 2). Thousands of their bones and teeth were washed into karst fissures that have subsequently been revealed by quarrying. This provides a rare opportunity to analyse large samples of fossil material needed for confident estimation of maximum lifespan. Importantly, these are the earliest diphyodont taxa (Fig. 2), with a single replacement of non-molar teeth and no molar tooth replacement31, and so estimates of lifespan are accurate to the time of the measured tooth-root formation.

Red nodes highlight the divergence of major lineages; Node 1 = divergence of the Pelycosaur lineage; Node 2 = the Therapsida clade; Node 3 = the Cynodontia clade; Node 4 = the Mammaliaformes clade; Node 5 = the Mammalia clade; Node 6 = the Theria clade. Superscript numbers denote references in the main text. Single asterisk (*) denotes the uncertain phylogenetic affinities of Kuehneotherium within the Mammaliaformes clade31. Double asterisks (**) denote the uncertain phylogenetic affinities of Arboroharamiya3.
The fossil sample studied included both isolated teeth and mandibles with multiple teeth or roots in situ. We applied PPC-SRµCT to 87 Morganucodon specimens (52 isolated teeth, 35 dentaries, all Morganucodon watsoni) and 119 Kuehneotherium specimens (116 isolated teeth, 3 dentaries) (see “Methods”). From these, 34 Morganucodon and 27 Kuehneotherium specimens were sufficiently well preserved for three observers to independently estimate lifespan from cementum increments. These estimates were compared to validate their accuracy and precision (see “Methods”; Supplementary Data 1). The remainder showed physical and/or diagenetic damage that prevented increment measurement (Supplementary Fig. 2).
The cementum of Morganucodon and Kuehneotherium (Fig. 1a, b) is distinguished from dentine in our PPC-SRµCT data by a distinct boundary layer separating the two tissues. This lies external to the granular layer of Tomes of the dentine and is interpreted as the hyaline layer of Hopewell–Smith (Fig. 1c–g). Synchrotron nanotomographic imaging (30 nm isotropic voxel size) highlights individual Sharpey’s fibre bundles (linking cementum to the periodontal ligament in extant mammals) visible in several exceptionally preserved specimens, which can be traced radially through the cementum (Fig. 1g). Across the tooth-root transverse axis, cementum is ∼10–70 μm radial thickness and displays a series of contrasting light and dark circumferential increments representing different material densities (Figs. 1e–g and 3a–d). Higher-density increments (represented by greater greyscale values) are on average 2–3 μm radial thickness, and lower density increments are 1–3 μm radial thickness (Fig. 1c–g and 3a–d). Individual increments can be followed continuously both longitudinally and transversely through the entire scanned volume of a tooth root (Fig. 3e, f).

a, b Transverse virtual thin sections of PPC-SRµCT reconstructions (0.33 μm3 voxel size). a Morganucodon specimen NHMUK PV M 104127 showing a 55-μm-thick layer of cementum around the root dentine. b Kuehneotherium specimen UMZC Sy 141 showing a 32-μm-thick layer of cementum. c, d Detail of the cementum of c NHMUK PV M 104127 and d UMZC Sy 141. Cementum increments highlighted by 14 and 9 multi-coloured arrows, respectively. e, f 3D segmentations of the cementum increments of e NHMUK PV M 104127 and f UMZC Sy 141. The colour of each increment corresponds to the colours of each arrow in c, d, respectively. Scale bars represent 100 µm in a, b, 30 µm in c, d and 30 µm in e, f.
Increment count accuracy, tooth eruption sequence and timing
We tested the accuracy of cementum increment counts for predicting lifespan in fossils by additional PPC-SRµCT imaging and counting of increments in the cementum of several teeth along the tooth row in eight dentulous Morganucodon specimens with a range of teeth in situ (Table 1) and of growth increments (lines of arrested growth (LAGs)) in the periosteal region of the dentary bone in two of these (Fig. 4). In both specimens where dentary LAGs are found, counts are identical with the cementum increments in the teeth (p3–m2; Fig. 4). Also, comparisons between counts of cementum increments are identical across all four premolars (p1–p4) and the anterior molars (m1–m2), in all specimens where they occur together (Table 1). This agreement between p1–m2 teeth and dentary increment counts indicates that growth in both teeth and jaws was following the same, circum-annual rhythm, as previously reported for multiple extant mammal species37. We consider this to be strong support for the accuracy of lifespan estimates based on these increment counts.

a Four lines of arrested growth and a fifth incipient one are visible within the periosteal region of the dentary, each highlighted by three-dimensional segmented bands of differing colour corresponding to coloured arrows in the accompanying transverse PPC-SRµCT slice. Only LAGs persisting through the volume are segmented and highlighted. This pattern is mirrored in b the anterior root of the m1 tooth, c the posterior root of the same m1 tooth and d the anterior root of the m2 tooth. Scale bars represent 30 µm.
The increment counts along Morganucodon dentary toothrows can also provide information on eruption sequence and timing (see Supplementary Note 3 for more details). The first permanent premolar to the second molar all erupted within 1 year, with the first molar erupting prior to the third and fourth premolars (NHMUK PV M 27312; see Supplementary Note 3 for more details). The ultimate incisor (i4), the canine and the third molar erupted in the following year. We do not have information on eruption timing of more anterior incisors, or the fourth molar, and the fifth molar is only occasionally present. As we estimate that Morganucodon was long lived relative to comparatively sized extant mammals (see below), this pattern of most of the adult tooth row being in place during the first 2 years of life is also supportive of a relatively short (compared with its total lifespan) juvenile stage46 and determinate growth47. The absolute length of these stages in Morganucodon is, however, considerably longer than extant mammals of comparable body size37. Unfortunately, dentulous specimens of Kuehneotherium are rare, and there are no tooth rows with cementum increment counts in our sample.
Long lifespans, low BMR and growth rates
Cementum increment counts provide a minimum estimate of maximum lifespan of 14 years for Morganucodon and 9 years for Kuehneotherium (Figs. 3 and 5a and Supplementary Data File 1). These may underestimate true maximum lifespan, as any damage to outer cementum increments would reduce estimated maximum lifespan. One-way analysis of variance (ANOVA) comparisons of mean intra-observer coefficient of variation (CV) between our study and ten previous cementochronological studies of different extant mammal species (see “Methods”) with similar age ranges suggest that values for PPC-SRµCT data (Shapiro–Wilk W = 1) of Morganucodon (CV = 9.32) and Kuehneotherium (CV = 4.89) are significantly lower than previous thin section-based studies (W = 0.93; minimum extant CV = 14.2, mean CV = 21.8, standard deviation = 5.87; F = 11.12, p < 0.01; Supplementary Table 1).

a Histogram of lifespan estimates from cementum increment counts. b Log10 phylogenetic least squares (PGLS) biplot of mean body mass (g) against maximum wild lifespan (years) for extant mammals (n = 279), extant non-avian reptiles (n = 252) and fossil mammaliaforms. c Log10 PGLS biplot of mean body mass (g) against mass-specific standard metabolic rate (msSMR; mL O2 h−1 g−1) for extant mammals (n = 117) and extant reptiles (n = 55) and estimates for fossil mammaliaforms. d Log10 PGLS biplot of mean body mass (g) against postnatal growth rate constant K (days−1) for extant mammals (n = 115) and extant reptiles (n = 33) and estimates for fossil mammaliaforms. PGLS regression lines in b–d are shown for extant mammals (black) and extant reptiles (blue), 95% confidence intervals are represented by dashed lines and 95% predictor intervals by dotted lines. Source data are provided as a Source data file.
We estimated body mass ranges of 10.7–25.0 g (mean 17.9 g) for Morganucodon and 14.9–32.7 g (mean 23.8 g) for Kuehneotherium (minimum mass estimates based on skull length21 and maximum mass estimates on dentary length48; see “Methods”). Maximum lifespan and mean body mass for the mammaliaforms were compared with published data for large samples of terrestrial, non-volant wild extant mammal (n = 278) and non-avian reptile (n = 256) species (“Methods”; Supplementary Data 3). Maximum wild lifespans of extant taxa were chosen for comparison with our fossil taxa, as these values are the closest analogue to our estimated lifespans, relative to captive lifespan values. To ensure robustness of our results, we additionally analysed maximum captive lifespans of extant taxa below 100 g, which show an average increase above maximum wild lifespans of approximately 3.43 and 4.38 years per taxon for mammals and reptiles, respectively (Supplementary Fig. 4). Broad results of statistical tests, and the overall conclusions of our study, are unchanged regardless of whether wild or captive data are used for analysis and comparisons between our fossil lifespans and the lifespans of extant taxa (see Supplementary Note 4 and Supplementary Figs. 1–6).
Phylogenetic generalized least squares (PGLS) regression of log10-transformed values shows that the fossil mammaliaforms fall within the range of extant reptiles and have longer maximum lifespans for their size and are further above the mammal regression mean, than all extant mammals under 4 kg (the long-lived and secondarily dwarfed49 mouse lemur Microcebus murinus is closest). Only the short-beaked echidna Tachyglossus aculeatus, a monotreme with long lifespan and low metabolic rate, exceeds the distance above the mammalian mean for Kuehneotherium, but not for Morganucodon (Fig. 5b). One-way phylogenetic analysis of covariance (ANCOVA) comparisons show that regression slopes for extant mammals and reptiles are statistically similar (p = 0.35) but their means are significantly separated (p = 0.036), with reptiles on average living 18.3 years longer than mammals of the same body mass.
To estimate BMR, we used PGLS and recovered significant correlations between log10-transformed values of maximum wild lifespan and mass-specific standard metabolic rate (msSMR; measured in mL O2 h−1 g−1 and analogous with BMR in extant mammals—SMR was used as BMR cannot be measured in reptiles50) from published data for 117 extant mammals and 55 extant reptiles (“Methods”; Supplementary Data 3 and Fig. 6a). Using the correlation between maximum wild reptile lifespan and msSMR and plotting our mammaliaforms directly onto this regression line, we estimated a reptile-derived msSMR of 0.055 mL O2 h−1 g−1 (Morganucodon) and 0.08 mL O2 h−1 g−1 (Kuehneotherium) (Fig. 6a). We additionally used the correlation between maximum wild mammal lifespan and msSMR and estimated a mammal-derived msSMR of 0.36 mL O2 h−1 g−1 for Morganucodon and 0.46 mL O2 h−1 g−1 for Kuehneotherium (Fig. 6a). When log10 PGLS is used to regress these estimates against body mass, both mammaliaforms fall outside the 95% predictor interval (PI) of the mammalian data and within the reptile range of msSMR, regardless of whether mammaliaform msSMR is estimated from reptilian or mammalian data (Fig. 5c). This suggests that the mammaliaforms had significantly lower msSMR values when compared to extant mammals of similar size. The comparably sized mammal (<100 g) of lowest msSMR is the marsupial Dasycercus cristicauda, with a maximum wild lifespan of 7 years and msSMR of 0.63 mL O2 h−1 g−1 (Fig. 5c).

a Log10 phylogenetic least squares (PGLS) biplot between maximum wild lifespan (years) and mass-specific standard metabolic rate (msSMR; mL O2 h−1 kg1) for extant mammals (n = 117) and reptiles (n = 55). b Log10 PGLS biplot between maximum wild lifespan (years) and postnatal growth rate constant K (days−1) for extant mammals (n = 115) and reptiles (n = 31). PGLS regression means for each clade (black lines for mammals, blue lines for reptiles, dashed lines denote 95% confidence intervals) are used to estimate msSMR and K for mammaliaforms Morganucodon and Kuehneotherium, with dashed brackets denoting their 95% confidence intervals. Source data are provided as a Source data file.
We estimated growth rates using PGLS correlations between maximum wild lifespan and growth rate33,34 from published data from 115 extant mammals and 30 extant reptiles (“Methods”; Supplementary Data 3 and Fig. 6b). From mammal data, we estimate growth rate constants K (days−1—see “Methods”) of 1.085−2 days−1 (Morganucodon) and 1.474e−2 days−1 (Kuehneotherium). From reptile data, we estimated K = 4.91−4 days−1 (Morganucodon) and K = 6.65−4 days−1 (Kuehneotherium) (Fig. 6b). Log10 PGLS regression against body mass again places both mammaliaforms outside the mammalian 95% PI and within the reptile growth rate range, whether estimated from mammalian or reptilian data (Fig. 5d). The lowest growth rate of any <100 g extant mammal is K = 3.24e−2 days−1 for the Mongolian gerbil Meriones unguiculatus.
In summary, our estimates of maximum lifespan provided by tomographic imaging of cementum increments in Morganucodon and Kuehneotherium are significantly longer than the maximum wild lifespan of any extant mammal of comparable body mass. These lifespans provide estimates of SMR/BMR and growth rate that are significantly lower than comparably sized extant mammals and instead correspond to those of extant reptiles.
Femoral blood flow shows intermediate MMR
To compare our fossil mammaliaform BMR estimates with MMR, we used a second proxy directly linked to MMR51. The ratio between nutrient foramen area and femur length has been used as an index for relative blood flow (Qi) through the femur during and after metabolically demanding exercise (Qi = rf4/L, where rf = foramen radius and L = femur length), previously shown to correlate well with MMR51. From µCT data of the six most complete Morganucodon femoral diaphyses available, we segmented all nutrient foramina (Fig. 7a) and estimated their area by measuring their minimal radii (see “Methods”). Kuehneotherium could not be included as no suitable femoral specimens are known.

a 3D reconstruction of Morganucodon femur using µCT (specimen UMZC EoPC 19_6, voxel size of 4 μm), with all identifiable foramina segmented and highlighted (red, green, gold and blue). b Log10 biplot of mean body mass (g) against estimated blood flow index (Qi; mm3) for extant non-avian reptiles (n = 22), extant varanid reptiles (n = 8), extant mammals (n = 69) and Morganucodon. GLS regression lines in b are shown for extant mammals (black), extant non-varanid reptiles (blue) and extant varanids (purple), with 95% confidence intervals represented by dashed lines. Scale bars in a represent 1 mm. Source data are provided as a Source data file.
We estimated a Qi of 3.829e−7 mm3 for Morganucodon and compared this with published and new data (“Methods”) for extant mammals (n = 69) and reptiles (n = 30). The latter includes varanids (n = 8), which in the absence of mammalian predators fill an active hunting niche and tend to have mammalian MMR levels while retaining reptilian BMR levels51 (Supplementary Table 2). One-way ANCOVA comparisons show that means of GLS regression slopes for extant mammals and non-varanid reptiles are significantly different (p < 0.01) while the slopes are similar (p = 0.16). Log10 GLS regression of body mass and Qi shows that Morganucodon is further above (higher Qi for its mass) the non-varanid reptile mean than all non-varanid reptiles (phylogenetically informed statistical comparisons were not used here due to the non-significant lambda values showing no phylogenetic signal in the taxa used, see “Methods”). However, Morganucodon is also slightly further from the mammalian mean than the non-varanid reptile mean and considerably closer to small non-varanid reptile species data points than those of small mammalian species (Fig. 7b). This intermediate Qi, and so inferred intermediate MMR, suggests that, while retaining typical reptilian BMR and growth rates, Morganucodon had MMR above non-varanid reptiles but not as high as mammals or actively foraging varanid reptiles.
Discussion
We have used two quantitative proxies to determine the metabolic status of early mammaliaforms. Relatively long lifespans for both Morganucodon and Kuehneotherium result in SMR/BMR and growth rate estimates equivalent to modern reptiles of comparable size and indeed at the higher lifespan/lower BMR/slower growth end of the reptile scale for Morganucodon. This is true whether we compare our fossil estimates to wild lifespans of extant taxa or estimate fossil “captive” lifespans and compare them to captive values for extant taxa. In contrast, femoral blood flow estimates (Qi) suggest that the MMR of Morganucodon was intermediate between extant non-varanid reptiles and mammals. We therefore infer that in Morganucodon increased MMR (and so also absolute aerobic capacity (AAC) = MMR – BMR) was initially selected for before BMR and that the MMR-first hypothesis6 is the best-supported model for the evolution of mammalian endothermy. We suggest that at least Morganucodon, if not also Kuehneotherium, occupied a metabolic grade approaching extant varanids: able to undergo longer bouts of aerobically demanding activity than non-varanid reptiles but not capable of sustaining either mammalian levels of aerobic activity or the elevated thermometabolism exhibited by living endotherms.
Evidence from non-mammalian synapsids (including changes in gait7, long bone histology15 and development of secondary osteological features correlated with increased metabolic rate17,18) indicate unquestionable changes in physiology from pelycosaur- to mammaliaform-grade taxa. Determinate growth47 and reduction of dental replacement (diphyodonty) in basal mammaliaforms permitted more precise occlusion52, which has been considered a key innovation in the development of mammalian endothermy by enabling increased assimilation and higher metabolism53. However, determinate growth and diphyodonty appear to have preceded the appearance of modern mammalian levels of endothermy, at least in Morganucodon and Kuehneotherium. We therefore suggest that the development of precise occlusion in basal mammaliaforms52 may be more associated with dietary specialization and niche partitioning1.
Comparison of our results to those of other recent studies of physiology in fossil synapsids supports the hypothesis of a complex, mosaic pattern for the evolution of endothermy, with different characters being selected for at different rates through time, and with respect to phylogeny. For example, the size diminution associated with the cynodont–mammaliaform transition54 may have reversed the evolutionary trajectory of some previous histological proxies for endothermy55, contributing to the complex, contradictory patterns observed. Our study also suggests that more work is needed to compare fossil and extant ectothermic and endothermic taxa directly in order to better understand their relative metabolic properties. Many previous studies rely on simple binary divisions, such as the presence/absence of fibrolamellar bone and/or respiratory nasal turbinates. These proxies cannot represent accurately the complex series of physiological characteristics that range between “ectothermy” and “endothermy” and are frequently distributed homoplastically across the synapsid phylogeny, individually and with respect to each other. Other studies provide relative data such as preserved apatite oxygen isotopes24 that allow comparisons with co-habiting ectothermic taxa but cannot be directly compared to extant data and so do not suggest where the studied fossil taxa fall in the metabolic spectrum of extant vertebrates. However, our results are compatible with recent work on living mammals, suggesting that the BMR of the Middle Jurassic (∼170 Ma) mammalian MRCA was comparable to present-day values13. This indicates that evolution towards modern-day mammalian endothermy occurred during the 25 million year-long Early Jurassic and suggests that the mammalian mid-Jurassic adaptive radiation4,5 was driven by this or vice versa.
In conclusion, our data offer a direct link to measurable aspects of endothermy, such as BMR and MMR, at a key point in mammalian evolution. Further work applying these methods to additional Mesozoic mammaliaforms and mammals, and comparison with evidence from other physiological characteristics, will allow the evolutionary tempo and mode of multiple aspects of mammalian physiology to be determined. The early mammaliaforms Morganucodon and Kuehneotherium possessed surprisingly low, reptile-like metabolic rates plus a mixture of plesiomorphic and derived characters7 relating to life history and physiology. Ultimately, we can no longer assume that the endothermic metabolism of living mammals had evolved in the earliest mammaliaforms.
Methods
Choice of fossil specimens
All Morganucodon and Kuehneotherium specimens are from the Early Jurassic St Brides Island fissure suite, from Glamorgan, South Wales (UK)1,56,57. Morganucodon specimens used for the cementum analysis are from the collections of the Natural History Museum, London, UK (NHMUK), Kuehneotherium specimens are from the collections of the NHMUK and University Museum of Zoology Cambridge, UK (UMZC) and the Morganucodon femora specimens are from the collections of UMZC. The Morganucodon specimens are all one species, M. watsoni, from Pontalun 3 fissure, excavated in 1962–1963 in Pontalun quarry (now known as Lithalun)45,56. Kuehneotherium is not abundant in Pontalun 3 fissure and the specimens used here for cementum analysis are from three Glamorgan fissures in Pontalun and Pant quarries: Pontalun 3, Pant 2, and Pant 4. Pontalun 3 and Pant 2 fissures have a relatively impoverished fauna, and Pant 4 has a more diverse biota45 (Supplementary Data 2). Kuehneotherium from Pontalun 3 fissure is Kuehneotherium praecursoris57, but those from the Pant fissures are considered to be different species, based on small molar differences58. All specimens from Pontalun 3 fissure in the NHMUK (Morganucodon and Kuehneotherium) were prepared by immersing dried blocks of clay matrix in hot tap water, with the addition of dilute hydrogen peroxide or sodium hexametaphosphate (Calgon) if required, but those in the UMZC collection were prepared with 10% acetic acid. Kuehneotherium specimens from Pant quarry (Pant 2 and Pant 4) were collected by the Kermack team from University College London, between 1955 and 1978, and are from a harder matrix, which was prepared with 15% acetic acid58.
The Morganucodon and Kuehneotherium specimens used for cementum analysis are either isolated teeth or dentary specimens with a range of teeth in situ. The lower dental formula of Morganucodon is i4:c1:p4:m4–5 and for Kuehneotherium it is i?:c1:p6:m61. In Morganucodon, the majority of isolated teeth measured were lower second molars (m2) since they are easily identified, are relatively large, have robust separated roots and, as anterior diphyodont molars, they should erupt relatively early and are not replaced, therefore offering a near-complete record of life history. However, i4, c, p3, p4, m1, m3 and m4 teeth were also studied in dentulous Morganucodon specimens. Dentulous Kuehneotherium specimens are extremely rare and so isolated teeth were scanned in almost all cases. For Kuehneotherium, all appropriate teeth were chosen; the distinctive p5 and p6, and a range of lower and upper molars, which can be identified to anterior, mid or posterior tooth row on the degree of triangulation58.
Tomographic imaging of cementum
Pilot scans of two Morganucodon lower second molars (NHMUK PV M 104131 and NHMUK PV M 104132) were carried out in 2011 on the nanotomography imaging beamline ID22 at the European Synchrotron Radiation Facility (ESRF), Grenoble, France (project EC 1064). For ID22, we used the following experimental settings for computed tomographic (CT) imaging: X-ray energy of 29.6 keV, 1999 projections over a 180° rotation, 0.5 ms exposure time, 321 nm voxel size, 405 mm sample-to-detector distance, diamond window, a 20-µm-thick LSO scintillator doped with Tb, and 1.5 mm Al filter, single propagation distance tomography.
During a 4-day experiment at the ID19 beamline of the ESRF (18/04/2014-22/04/2014), 71 additional Morganucodon specimens (52 isolated teeth and 19 dentaries) and 2 pilot Kuehneotherium specimens were scanned (project ES 152). A single harmonic U13 undulator was used as the X-ray source, delivering a pink X-ray beam with peak energy at 26.5 keV, with a 1.4 mm Al filter used to cut the background lower energies. The detector was a microscope optic system coupled to a sCMOS sensor (PCO edge 5.5), mounted with a 10-µm thick GGG:Eu scintillator. Scans were performed using single propagation distance tomography (15 mm sample-to-detector propagation distance), an exposure time of 250–300 ms, 2499 angular projections over a 360° scan and at voxel sizes of 280, 347 and 700 nm.
Subsequently, 117 additional Kuehneotherium specimens (116 isolated teeth and one dentary) and 12 additional Morganucodon dentary specimens were scanned during a 3-day experiment at the TOMCAT tomographic beamline of the Swiss light Source (SLS), Villigen, Switzerland (13/04/2015-16/04/2015). The beam was set at an energy of 20 keV using a double multilayer monochromator, a LSO:Tb scintillator and a pco.EDGE 5.5 detector. Samples were scanned using single propagation distance tomography (14 mm sample-to-detector propagation distance), an exposure time of 150 ms and 1500 angular projections over a 180° scan at a voxel size of 330 nm. A Kuehneotherium lower molar (UMZC Sy 141) was imaged at 1.2 μm voxel size (with an exposure time of 150 ms and 1500 angular projections over 180°) to provide the 3D volume presented in Fig. 1b.
Three juvenile Morganucodon dentary specimens, with roots from final deciduous premolars (NHMUK PV M 27312, NHMUK PV M 27474 and NHMUK PV M 27475), and an older individual with extensive molar wear (NHMUK PV M 27465), were scanned during a 3-day experiment at the TOMCAT beamline of the SLS (07/03/2016-10/03/2016). The beam energy was set at 21 keV using a double multi-layer monochromator. Samples were scanned using single propagation distance tomography (14 mm sample-to-detector propagation distance), an exposure time of 200 ms and 1601 angular projections over a 180° scan at a voxel size of 330 nm.
CT reconstructions of the above tomographic data were generated using a filtered back-projection algorithm coupled with “Paganin-style” single distance phase retrieval59 algorithms developed in-house at the respective beamlines60,61. For data from ID19, β = 8.1 × 10−8, δ = 9.8 × 10−9. For data from TOMCAT, β = 3.7 × 10−8, δ = 1.7 × 10−10.
Of the 71 Morganucodon molar specimens imaged at beamline ID19 in 2014, 4 were additionally imaged at the nano-imaging beamline ID16A of the ESRF synchrotron (project ES 152). These were imaged using holotomography62 from four propagation distances. Holograms were recorded using a charge-coupled device detector with an effective pixel size of 3 µm and a 23-µm-thick GGG:Eu scintillator at both 17 keV and 33.6 keV. The selected voxel sizes were 10, 25, 30 and 130 nm. The number of angular projections recorded over 180° varied between 1200 and 2000 and the exposure times were set at 250–800 ms. To generate the image containing the virtual section in Fig. 1g with 30 nm voxel size, the four focus-to-sample distances were 2.65, 13.19, 15.36 and 19.87 mm and the sample-to-detector distance was 1.2648 m.
To produce the 3D model presented in Fig. 1a, a Morganucodon lower molar (NHMUK PV M 104134) was imaged using μCT at the University of Helsinki in March 2013. μCT was performed using a Nanotom 180 NF (phoenix X-ray Systems & Services GmbH) with a CMOS detector (Hamamatsu Photonics) and a high-power transmission-type X-ray nanofocus source with a tungsten anode. A total of 900 angular projections were collected for a 180° rotation, at an exposure time of 1840 ms and a voxel size of 2 × 2 × 2 μm3. The raw projection data were reconstructed using filtered back-projection by the reconstruction software datos|x rec, supplied by the system manufacturer.
Increment counting and creation of virtual thin sections
Cementum increments were counted in CT data using modifications to the techniques suggested by the Cementochronology Research Program38 to take into account the 3D nature of the PPC-SRµCT cementum data. First, the cementum was visually inspected throughout the entire volume of each scan, in transverse PPC-SRµCT slices using ImageJ/Fiji63 to distinguish between specimens that could be confidently interpreted as preserving cementum increments or those that were too badly affected by diagenesis for increment counting. Phase-contrast imaging of incremental features is understood to be prone to recurrent destructive interference patterns from Fresnel diffraction that create periodic blurring at differing frequencies when they are scanned using inappropriate experimental parameters (principally X-ray energy, sample-to-detector propagation distance and voxel size). However, our parameters produce blurring frequencies that are too narrow (approximately 500–900 nm) to significantly affect the contrast between cementum increments (1–3 μm radial thickness) (Tafforeau, personal observation). Therefore, no significant masking of increments from Fresnel diffraction blurring should be expected in our data. In specimens that preserved increments, volumes were inspected by eye to identify regions of highest increment contrast with no lensing and/or coalescence between increments. Increments identified in these regions were followed by eye throughout the entire cementum tissue surrounding these regions, both longitudinally and transversely through the root, in order to distinguish between principal increments and accessory increments formed by lensing and coalescence of primary increments in discrete portions of the tissue (Supplementary Fig. 1). Primary increments were distinguished as those that persisted vertically through the entire scanned region of cementum, whereas accessory increments lasted only for short periods before coalescing into neighbouring increments (Supplementary Fig. 1).
Once regions of highly contrasting primary cementum increments had been identified, virtual thin sections of these regions were created. This was performed by isolating ten transverse PPC-SRµCT slices through each region and summing their greyscale values using the “Sum slices” option of the “Z projection” tool in ImageJ/Fiji to create a new image of increased contrast between dark and light cementum increments and reduced image noise. Between three and five virtual thin sections were created for all specimens with readable cementum increments. For each virtual thin section, increments were counted manually by three different observers: Observer One (E.N.) had considerable experience in counting cementum increments (>100 specimens studied); Observer Two (K.W.) had training in counting cementum increments (30 specimens studied under guidance from Observer One) and experience in studying growth patterns in PPC-SRµCT data of long-bones; Observer Three (C.N.) had no prior experience in counting increments or studying growth patterns. Each observer studied virtual thin sections blind, after collections of virtual thin sections were numbered and randomized between specimens using the RAND function in Microsoft Excel. For each observer, the final increment count for every specimen was determined as the maximum number counted in all of its virtual thin sections.
Once each observer had counted increments in every virtual thin section (Supplementary Fig. 3 and (Supplementary Data 1) and the precision between their increment counts was compared by calculating the CV (Eq. 1; Supplementary Table 1):
3D modelling of cementum increments
3D modelling was performed, using the Avizo image analysis software (Avizo 8.0; Thermo Fisher Scientific), on a subsample of teeth comprising the first tooth imaged using PPC-SRµCT (NHMUK PV M 104131) and the specimens that provided the highest cementum increment counts for each fossil taxon (NHMUK M 104127 for Morganucodon and UMCZ Sy 141 for Kuehneotherium). Original tomographic data were downsampled in each axis by a factor of two to decrease manual processing time while retaining sufficient spatial resolution to preserve the cementum increments. Principal increments originally identified by eye were manually traced in each PPC-SRµCT slice and assigned to different materials in the “label field” tool-kit. Models were subsequently created and analysed using the “surface view” feature of Avizo. This allowed the pattern of incrementation to be viewed with a 3D perspective, to test whether increments defined in two-dimensional slices were true principal increments or diffuse accessory increments (Supplementary Fig. 1).
Fossil body mass estimation and choice of extant taxa
Body mass for our fossil taxa was estimated using two techniques, based on scaling between single cranial dimensions and measured body mass in extant mammals. Maximum body mass estimates were made using the scaling relationship (Eq. 2) between dentary length (mm) and body mass (g) published for extant marsupial mammals of small body mass and subsequently used to estimate body mass in several Mesozoic fossil taxa from the Late Jurassic Morrison Formation and elsewhere48:
Dentary lengths are from published CT reconstructions1 (20 mm for Morganucodon and 21.9 mm for Kuehneotherium), resulting in body mass estimates of 25.0 g and 32.7 g respectively. However, this estimate may be an overestimate for Kuehneotherium as it has a longer, more gracile, dentary relative to Morganucodon, due to the differing feeding ecologies of the two taxa1.
Second, minimum body mass estimates were calculated using the scaling relationship (Eq. 3) between skull length (mm) and body mass (g) found for 64 extant species of small “lipotyphlan” insectivores64 and used to estimate the body masses of several Mesozoic mammaliaforms21,64,65.
Due to a lack of complete, diagnostic cranial material for UK samples of Morganucodon and Kuehneotherium, their skull lengths were estimated using a scaling relationship between dentary length and skull length calculated for Morganucodon oehleri from Rowe et al. Figure S121 (skull length (mm) = 1.0458 × jaw length (mm)). Following this relationship, we used dentary length1,21 to estimate a skull length of 21.0 mm for M. watsoni and 22.9 mm for Kuehneotherium, which results in body mass estimates of 10.7 and 14.9 g, respectively (as above, this may be an overestimate for Kuehneotherium). Body masses presented in Figs. 5 and 7 are mean values of the two estimates (17.9 g for Morganucodon and 23.8 g for Kuehneotherium).
Extant data
Information on maximum wild lifespan and mean body mass was obtained for 278 extant terrestrial mammal species (body mass: mean = 70.2 kg, standard deviation = 395 kg; wild lifespan: mean = 10.8 years, standard deviation = 11.2 years) and 256 extant terrestrial non-avian reptile species (body mass: mean = 3.6 kg, standard deviation = 20 kg; wild lifespan: mean = 12 years, standard deviation = 14 years) (Supplementary Data File 3). Information on maximum captive lifespan was obtained for 644 extant terrestrial mammal species (body mass: mean = 62.2 kg, standard deviation = 324 kg; captive lifespan: mean = 15.5 years, standard deviation = 11.5 years) and 866 extant reptile species (body mass: mean = 2.4 kg, standard deviation = 15.9 kg; captive lifespan: mean = 15.3 years, standard deviation = 11.6 years). Information for flying or gliding taxa (including birds) was not included as their body masses are known to be secondarily reduced to aid their lifestyle and so may distort observed overall relationships between lifespan and body mass. In particular, birds and bats live on average three-to-four times as long as terrestrial mammals of similar body mass66. Similarly, marine mammals were not included as their environment positively affects body mass, allowing significantly higher body masses than terrestrial taxa67. Mammalian wild and captive lifespan data were obtained from the primary literature. The majority of these data originated from a download from the online databases of the Max Planck Institute (https://www.demogr.mpg.de/longevityrecords/0203.htm) on 10/03/2019. The majority of reptile wild and captive lifespans were obtained from the supplementary information of Scharf et al.68. Body mass estimates were obtained from an online Ecological Archives database69 (http://www.esapubs.org/archive/ecol/E084/094/metadata.htm), the AnAge database (https://genomics.senescence.info/species/) and published data50,68. Although the Max Planck Institute data are entitled “longevity records”, it notes in the introduction that this is equivalent to “highest documented age”. The authors of the database believe that, despite any analytical and conceptual shortcomings, the records are useful in a number of comparative and disciplinary contexts, including demographic, gerontological, ecological and evolutionary. Similarly, the AnAge database states that maximum longevity is synonymous with maximum lifespan.
Data on SMR for extant mammals (n = 117; mean = 915; standard deviation = 3018) and reptiles (n = 55; mean = 62.3; standard deviation = 159) was obtained from a published electronic appendix50. SMRs for mammals were included in this data set only if measured under basal metabolic conditions50 and so are synonymous with BMRs.
Data on the postnatal growth rate constant K was obtained upon request for extant mammals32 (n = 115, mean = 0.03, standard deviation = 0.03) and reptiles (n = 33, mean = 0.002, standard deviation = 0.003) from previous publications70,71. The growth rate constant K is a magnitude free measure of growth rate, measured in days−1, that is considered the best measure for comparing growth rates between multiple species72.
Physiological metrics (lifespan/body mass/SMR/K) were compared using PGLS regression in the “R” statistical environment with the “ape”, “geiger”, “nlme” and “phytools” packages installed. For each regression, phylogenetic subsets of squamate and mammal taxa were downloaded from https://vertlife.org (100 trees per subset, birth-death node-dated completed tree distribution for mammals) representing the phylogenetic relationships of every taxon in the respective subsample. For crocodilian taxa, a phylogeny was manually constructed in R following the time-calibrated phylogeny of Oaks73 (using node mean age values from the species tree/90 My maximum analysis) and added to the base of the squamate phylogeny to produce a reptile clade. Each subset was investigated to find the tree that produced the highest PGLS F values and this tree was then input into the “corPagel” covariance structure for the “gls()” function to produce a phylogenetically informed regression model between the respective metrics.
The relationships between body mass and maximum lifespan were compared between extant mammals and extant reptiles using phylogenetic ANCOVA following Smaers et al.74 using the “gls.ancova” function in “R” with the same libraries installed as for PGLS. The individual phylogenetic subsets representing the mammal and reptile samples were combined with connecting branches connecting the Last Common Ancestors (LCAs) of the crocodilian, squamate and mammal clades. The LCA node connecting squamates and crocodilians was dated at 275.9 Ma using the mean of minimum and maximum node ages from Benton et al.75; similarly, the LCA node connecting reptiles (squamates and crocodiles) and mammals was dated at 325.5 Ma75. The mammal and reptile clades were then distinguished using two factor elements, and the influence of these factors upon the fit of the data to a series of models of increasing complexity was tested. First, a baseline model was produced consisting of one slope and one intercept for the entire data set. Three models were then compared to this baseline; the first varied in slope between mammals/reptiles, keeping their intercepts constant; the second allowed the intercept to vary between mammals/reptiles, keeping their slopes constant; and the final model allowed both slope and intercept to vary between mammals/reptiles. Comparisons between these models allowed the determination of their improvements upon the baseline model. Phylogenetic ANCOVA identified a lambda value of <0.001 between body mass and Qi for reptiles and mammals, and so non-phylogenetically informed GLS and ANCOVA were used to compare these metrics.
μCT study of femoral nutrient foramina
Nutrient foramina in the femora of Morganucodon and 11 extant mammal taxa of comparable size were imaged using μCT in the X-Ray Micro-Imaging Laboratory, University of Helsinki (Supplementary Table 2). We used μCT data of femoral foramina rather than photographic images of the foramina51 because many of the Morganucodon foramina were filled with sediment, making their photographic measurement difficult or impossible, whereas the differential density of sediment and fossilized bone in μCT data allowed visualization and analysis. Six Morganucodon femur specimens were selected for analysis from the collections of the Cambridge University Museum of Zoology based on their relative completeness. All femora (both fossil and extant) were scanned using a Bruker Skyscan 1272 μCT scanner at 70 kVp source tension and with a 0.5 mm Al filter. For each scan, 1125–1800 angular projections were collected during a 180° rotation, at an exposure time of 1344 ms. One fossil specimen was scanned at an isotropic voxel size of 4 μm (Fig. 7a) and five at 5 μm. All femora from extant taxa were scanned at 4 μm voxel sizes. The raw projection data were reconstructed using filtered back-projection with the Feldkamp algorithm by the Bruker reconstruction software “NRecon” (Version 1.7.1.0).
Analysis of μCT femora data was conducted using the Avizo image analysis software (Version 9.3.0; Thermo Fisher Scientific). Foramina were located using the “Orthoslice” tool, to scroll through transverse CT slices of each femur. Once located, the foramen was imaged in 3D using the “Volume rendering” tool. This allowed the minimum diameter, and so minimum radius (in cm), of the foramen aperture to be assessed and measured using the “3D line measurement” tool. Minimum diameter was used by Seymour et al.51, in the study of foramina in small mammals, as it is not always possible to see the direction of nutrient vessel penetration. Resulting foramina radii were used to generate estimates of blood flow index (Qi) following the method outlined in Seymour et al.51. If multiple foramina were found in a single specimen, then the radii were summed following the assumption that this represents the total entry/exit potential of nutrient circulation through the femur51. This sum radius, or the single radius in specimens with single foramina (both abbreviated to r; measured in cm), was then used along with the length of the femur (L; measured in cm) to estimate Qi as follows (Eq. 4):
Although Morganucodon femoral elements are relatively common in the Hirmeriella fissure suite, almost all are incomplete76. We here chose the six most complete femoral specimens known from the Pontalun 3 fissure, from where the Morganucodon teeth imaged using PPC-SRµCT originated. Specimens chosen (UMZC EoPC 19_1 to EoPC 19_6) preserve at least two-thirds of the femoral shaft. Length was estimated from the minimum mid-shaft diaphysial width of each element, using the scaling relationship between the two measurements established for a reconstructed Morganucodon femur created by concatenating three incomplete femora by P.G.G. (Eq. 5):
Statistics
One-way ANOVA comparison of intra-observer CV between cementochronological studies; Shapiro–Wilk normality test: PPC-SRµCT data W = 1, p = 1, histological data W = 0.93, p = 0.41; test statistics F = 11.12, degrees of freedom (df) = 10, Cohen’s effect size d = 3.13, p = 0.00728. Phylogenetic ANCOVA comparison of PGLS regression slopes for lifespan against body mass in mammals (log10 lifespan = 0.26(log10 body mass) + 0.16; 95% confidence interval (CI) = 0.05; r2 = 0.69) and reptiles (log10 lifespan = 0.26(log10 body mass) + 0.60; 95% CI = 0.08); r2 = 0.46); slopes are statistically similar (F = 0.868, p = 0.352) while means are significantly separated (F = 4.44, df = 529, partial eta squared effect size = 0.32, p = 0.036). PGLS regression of mammalian lifespan against msSMR: log10 msSMR = −0.237(log10 lifespan) – 0.083; 95% CI = 0.07; r2 = 0.59, p < 0.001. PGLS regression of reptilian lifespan against msSMR: log10 msSMR = −0.83(log10 lifespan) – 0.31; 95% CI = 0.255; r2 = 0.43, p < 0.01. PGLS regression of mammalian lifespan against growth constant K; log10 K = −0.692(log10 lifespan) – 1.171; 95% CI = 0.101; r2 = 0.66, p < 0.01. PGLS regression of reptilian lifespan against growth constant K; log10 K = −0.69(log10 lifespan) – 2.523; 95% CI = 0.339; r2 = 0.43, p < 0.01. One-way ANCOVA comparison of OLS regression slopes for Qi against body mass in extant mammals (log10(Qi) = 0.513 × log10(body mass) – 6.104) and non-varanid reptiles (log10(Qi) = 0.685 × log10(body mass) – 8.139); slopes are statistically similar (F = 2, p = 0.16) while means are significantly different (F = 87.6, df = 89, partial eta squared effect size = 0.50, p = 7.4E−15).
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The tomographic data that support the findings of this study are available from the corresponding authors upon reasonable request. Virtual thin sections used for cementum and bone increment counts are available at the University of Southampton’s PURE data repository as data number D1506 (https://doi.org/10.5258/SOTON/D1506). Physiological and phylogenetic data are from online databases of the Max Planck Institute (https://www.demogr.mpg.de/longevityrecords/0203.htm), an online Ecological Archives database (http://www.esapubs.org/archive/ecol/E084/094/metadata.htm), the AnAge database (https://genomics.senescence.info/species/), the VertLife online project (https://vertlife.org) and the literature (references in Supplementary Data file 3) and are provided in Supplementary Tables, as Supplementary Data files, and as a part of the Source data provided with this paper.
References
Gill, P. G. et al. Dietary specializations and diversity in feeding ecology of the earliest stem mammals. Nature 512, 303–305 (2014).
Luo, Z. X. Transformation and diversification in early mammal evolution. Nature 450, 1011–1019 (2007).
Han, G., Mao, F., Bi, S., Wang, Y. & Meng, J. A Jurassic gliding euharamiyidan mammal with an ear of five auditory bones. Nature 551, 451–456 (2017).
Wilson, G. P. et al. Adaptive radiation of multituberculate mammals before the extinction of dinosaurs. Nature 483, 457–460 (2012).
Close, R. A., Friedman, M., Lloyd, G. T. & Benson, R. B. Evidence for a mid-Jurassic adaptive radiation in mammals. Curr. Biol. 25, 2137–2142 (2015).
Bennett, A. F. & Ruben, J. A. Endothermy and activity in vertebrates. Science 206, 649–654 (1979).
Kemp, T. S. The origin of mammalian endothermy: a paradigm for the evolution of complex biological structure. Zool. J. Linn. Soc. 147, 473–488 (2006).
Clarke, A. & Pörtner, H. O. Temperature, metabolic power and the evolution of endothermy. Biol. Rev. 85, 703–727 (2010).
Koteja, P. Energy assimilation, parental care and the evolution of endothermy. Proc. R. Soc. B 267, 479–484 (2000).
Hopson, J. A. The role of foraging mode in the origin of therapsids: implications for the origin of mammalian endothermy. Fieldiana Life Earth Sci. 49, 126–149 (2012).
Farmer, C. G. Reproduction: the adaptive significance of endothermy. Am. Nat. 162, 826–840 (2003).
Lovegrove, B. G. The evolution of endothermy in Cenozoic mammals: a plesiomorphic-apomorphic continuum. Biol. Rev. 87, 128–162 (2012).
Avaria-Llautureo, J., Hernández, C. E., Rodríguez-Serrano, E. & Venditti, C. The decoupled nature of basal metabolic rate and body temperature in endotherm evolution. Nature 572, 651–654 (2019).
Gross, W. Die typen des mikroskopischen knochenbaues bei fossilen stegoccphalen und reptilien. Anat. Embryol. 103, 731–764 (1934).
Ray, S., Botha, J. & Chinsamy, A. Bone histology and growth patterns of some nonmammalian therapsids. J. Vert. Paleontol. 24, 634–648 (2004).
Shelton, C. D. & Sander, P. M. Long bone histology of Ophiacodon reveals the geologically earliest occurrence of fibrolamellar bone in the mammalian stem lineage. C. R. Palevol. 16, 397–424 (2017).
Benoit, J., Manger, P. R. & Rubidge, B. S. Palaeoneurological clues to the evolution of defining mammalian soft tissue traits. Sci. Rep. 6, 25604 (2016).
Hillenius, W. J. Turbinates in therapsids: evidence for Late Permian origins of mammalian endothermy. Evolution 48, 207–229 (1994).
Benoit, J., Fernandez, V., Manger, P. R. & Rubidge, B. S. Endocranial casts of pre-mammalian therapsids reveal an unexpected neurological diversity at the deep evolutionary root of mammals. Brain Behav. Evol. 90, 311–333 (2017).
Rowe, T. B. Definition, diagnosis, and origin of Mammalia. J. Vert. Paleontol. 8, 241–264 (1988).
Rowe, T. B., Macrini, T. E. & Luo, Z. X. Fossil evidence on origin of the mammalian brain. Science 332, 955–957 (2011).
Hoffman, E. A. & Rowe, T. B. Jurassic stem–mammal perinates and the origin of mammalian reproduction and growth. Nature 561, 104–108 (2018).
Kielan-Jaworowska, Z. & Hurum, J. H. Limb posture in early mammals: sprawling or parasagittal. Acta Palaeontol. Pol. 51, 393–406 (2006).
Rey, K. et al. Oxygen isotopes suggest elevated thermometabolism within multiple Permo-Triassic therapsid clades. eLife 6, e28589 (2017).
Huttenlocker, A. K. & Farmer, C. G. Bone microvasculature tracks red blood cell size diminution in Triassic mammal and dinosaur forerunners. Curr. Biol. 27, 48–54 (2017).
Olivier, C., Houssaye, A., Jalil, N. E. & Cubo, J. First palaeohistological inference of resting metabolic rate in an extinct synapsid, Moghreberia nmachouensis (Therapsida: Anomodontia). Biol. J. Linn. Soc. 121, 409–419 (2017).
Crompton, A. W., Owerkowicz, T., Bhullar, B. A. & Musinsky, C. Structure of the nasal region of non–mammalian cynodonts and mammaliaforms: speculations on the evolution of mammalian endothermy. J. Vertebrate Paleontol. 37, e1269116 (2017).
Rodrigues, P. G. et al. Digital cranial endocast of Riograndia guaibensis (Late Triassic, Brazil) sheds light on the evolution of the brain in non-mammalian cynodonts. Hist. Biol. 31, 1195–1212 (2019).
Hayes, J. P. & Garland, T. Jr The evolution of endothermy: testing the aerobic capacity model. Evolution 49, 836–847 (1995).
Köhler, M., Marín-Moratalla, N., Jordana, X. & Aanes, R. Seasonal bone growth and physiology in endotherms shed light on dinosaur physiology. Nature 487, 358–361 (2012).
Kielan-Jaworowska, Z., Cifelli, R. L. & Luo, Z. X. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure (Columbia University Press, New York, 2004).
Hulbert, A. J., Pamplona, R., Buffenstein, R. & Buttemer, W. A. Life and death: metabolic rate, membrane composition, and life span of animals. Physiol. Rev. 87, 1175–1213 (2007).
Magalhães, J. P. D., Costa, J. & Church, G. M. An analysis of the relationship between metabolism, developmental schedules, and lifespan using phylogenetic independent contrasts. J. Gerontol. A Biol. Sci. Med. Sci. 62, 149–160 (2007).
Werner, J., Sfakianakis, N., Rendall, A. D. & Griebeler, E. M. Energy intake functions and energy budgets of ectotherms and endotherms derived from their ontogenetic growth in body mass and timing of sexual maturation. J. Theor. Biol. 444, 83–92 (2018).
Grady, J. M., Enquist, B. J., Dettweiler-Robinson, E., Wright, N. A. & Smith, F. A. Evidence for mesothermy in dinosaurs. Science 344, 1268–1272 (2014).
Dong, X., Milholland, B. & Viig, J. Evidence for a limit to human lifespan. Nature 538, 257–259 (2016).
Klevezal, G. Recording Structures of Mammals (CRC Press, Boca Raton, FL, 1995).
Naji, S. et al. Cementochronology, to cut or not to cut? Int. J. Paleopathol. 15, 113–119 (2016).
Lieberman, D. E. Life history variables preserved in dental cementum microstructure. Science 261, 1162–1164 (1993).
Stock, S. R. et al. Cementum structure in Beluga whale teeth. Acta Biomater. 48, 289–299 (2017).
Stutz, A. J. Polarizing microscopy identification of chemical diagenesis in archaeological cementum. J. Archaeol. Sci. 29, 1327–1347 (2002).
Newham, E. Exploring the Use of X–ray Tomography for the Quantification of Cementum Growth Patterns Across the Mammal Phylogeny. PhD thesis, Univ. Southampton (2018).
Le Cabec, A., Tang, N. K., Ruano Rubio, V. & Hillson, S. Nondestructive adult age at death estimation: visualizing cementum annulations in a known age historical human assemblage using synchrotron X‐ray microtomography. Am. J. Phys. Anthropol. 168, 25–44 (2019).
Wittwer–Backhofen, U., Gampe, J. & Vaupel, J. W. Tooth cementum annulation for age estimation: results from a large known-age validation study. Am. J. Phys. Anthropol. 123, 119–129 (2004).
Whiteside, D. I., Duffin, C. J., Gill, P. G., Marshall, J. E. & Benton, M. J. The Late Triassic and Early Jurassic fissure faunas from Bristol and South Wales: stratigraphy and setting. Palaeontol. Pol. 67, 257–287 (2016).
Luo, Z.-X., Kielan-Jaworowska, Z. & Cifelli, R. L. Evolution of dental replacement in mammals. Bull. Carnegie Mus. Nat. Hist. 36, 159–175 (2004).
O’Meara, R. N. & Asher, R. J. The evolution of growth patterns in mammalian versus nonmammalian cynodonts. Paleobiology 42, 439–464 (2016).
Foster, J. R. Preliminary body mass estimates for mammalian genera of the Morrison Formation (Upper Jurassic, North America). PaleoBios 28, 114–122 (2009).
Yoder, A. D. et al. Geogenetic patterns in mouse lemurs (genus Microcebus) reveal the ghosts of Madagascar’s forests past. Proc. Natl Acad. Sci. USA 113, 8049–8056 (2016).
White, C. R., Phillips, N. F. & Seymour, R. S. The scaling and temperature dependence of vertebrate metabolism. Biol. Lett. 2, 125–127 (2005).
Seymour, R. S., Smith, S. L., White, C. R., Henderson, D. M. & Schwarz-Wings, D. Blood flow to long bones indicates activity metabolism in mammals, reptiles and dinosaurs. Proc. R. Soc. B 279, 451–456 (2011).
Jäger, K. R., Gill, P. G., Corfe, I. & Martin, T. Occlusion and dental function of Morganucodon and Megazostrodon. J. Vertebr. Paleontol. https://doi.org/10.1080/02724634.2019.1635135 (2019).
Kemp, T. S. The Origin and Evolution of Mammals (Oxford University Press, Oxford, 2005).
Lautenschlager, S., Gill, P. G., Luo, Z. X., Fagan, M. J. & Rayfield, E. J. The role of miniaturization in the evolution of the mammalian jaw and middle ear. Nature 561, 533–537 (2018).
Botha–Brink, J., Codron, D., Huttenlocker, A. K., Angielczyk, K. D. & Ruta, M. Breeding young as a survival strategy during Earth’s greatest mass extinction. Sci. Rep. 6, 24053 (2016).
Kermack, K. A., Mussett, F. & Rigney, H. W. The lower jaw of Morganucodon. Zool. J. Linn. Soc. 53, 87–175 (1973).
Kermack, D. M., Kermack, K. A. & Mussett, F. The Welsh pantothere Kuehneotherium praecursoris. Zool. J. Linn. Soc. 47, 407–423 (1968).
Gill, P. G. Kuehneotherium from the Mesozoic Fissure Fillings of South Wales. PhD thesis, Univ. Bristol (2004).
Paganin, D., Mayo, S. C., Gureyev, T. E., Miller, P. R. & Wilkins, S. W. Simultaneous phase and amplitude extraction from a single defocused image of a homogeneous object. J. Microsc. 206, 33–40 (2002).
Marone, F., Studer, A., Billich, H., Sala, L. & Stampanoni, M. Towards on-the-fly data post-processing for real-time tomographic imaging at TOMCAT. Adv. Struct. Chem. Imaging 3, 1 (2017).
Mirone, A., Brun, E., Gouillart, E., Tafforeau, P. & Kieffer, J. The PyHST2 hybrid distributed code for high speed tomographic reconstruction with iterative reconstruction and a priori knowledge capabilities. Nucl. Instrum. Methods Phys. Res. B 324, 41–48 (2014).
Mokso, R. & Cloetens, P. Nanoscale zoom tomography with hard X rays using Kirkpatrick-Baez optics. Appl. Phys. Lett. 90, 144104 (2007).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2001).
Luo, Z. X., Crompton, A. W. & Sun, A. L. A new mammaliaform from the Early Jurassic and evolution of mammalian characters. Science 292, 1535–1540 (2001).
Ji, Q., Luo, Z. X., Yuan, C. X. & Tabrum, A. R. A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science 311, 1123–1127 (2006).
Munshi–South, J. & Wilkinson, G. S. Bats and birds: exceptional longevity despite high metabolic rates. Ageing Res. Rev. 9, 12–19 (2010).
Chikina, M., Robinson, J. D. & Clark, N. L. Hundreds of genes experienced convergent shifts in selective pressure in marine mammals. Mol. Biol. Evol. 33, 2182–1292 (2016).
Scharf, I. et al. Late bloomers and baby boomers: ecological drivers of lifespan in squamates and the tuatara. Glob. Ecol. Biogeogr. 24, 396–405 (2015).
Smith, F. A. et al. Body mass of late Quaternary mammals. Ecology 84, 3403 (2003).
Werner, J. & Griebeler, E. M. Allometries of maximum growth rate versus body mass at maximum growth indicate that non–avian dinosaurs had growth rates typical of fast growing ectothermic sauropsids. PLoS ONE 9, e88834 (2014).
Case, T. J. On the evolution and adaptive significance of postnatal growth rates in the terrestrial vertebrates. Q. Rev. Biol. 53, 243–282 (1978).
Zullinger, E. M., Ricklefs, R. E., Redford, K. H. & Mace, G. M. Fitting sigmoidal equations to mammalian growth curves. J. Mammal. 65, 607–636 (1984).
Oaks, J. R. A time-calibrated species tree of Crocodylia reveals a recent radiation of the true crocodiles. Evolution 65, 3285–3297 (2011).
Smaers, J. B. & Rohlf, F. J. Testing species’ deviation from allometric predictions using the phylogenetic regression. Evolution 70, 1145–1149 (2016).
Benton, M. J. et al. Constraints on the timescale of animal evolutionary history. Palaeontol. Electron. 18, 1–106 (2015).
Jenkins, F. A. & Parrington, F. R. The postcranial skeletons of the Triassic mammals Eozostrodon, Megazostrodon and Erythrotherium. Philos. Trans. R. Soc. Lond. B 273, 387–431 (1976).
Acknowledgements
We acknowledge the European Synchrotron Radiation Facility, Grenoble, France for provision of synchrotron radiation facilities on beamlines ID19 and ID16A (project ES152) and thank Peter Cloetens for assistance in using beamline ID16A. We also acknowledge the Paul Scherrer Institut, Villigen, Switzerland for provision of synchrotron radiation beamtime at beamline TOMCAT of the Swiss Light Source (project 20141278). The research leading to these results has received funding from the European Community’s Seventh Framework Programme (FP7/2007-2013) under grant agreement no. 312284 (for CALIPSO). We thank Keijo Hämäläinen for his help in the initial stages of the synchrotron imaging. This project was part-funded by a Natural Environmental Research Council studentship and an Engineering and Physical Sciences Research Council studentship, awarded to E.N. and P.S. (Grant number NE/R009783/1), and we also thank the Academy of Finland for part-funding the project. Thank you to the Natural History Museum London for contributing to travel for P.B. via the Departmental Investment Fund for Earth Sciences. We thank Ginko Investments Ltd. for funding for materials and travel and the University of Bristol Bob Savage memorial fund for travel for E.N. Many thanks to the Natural History Museum, London, University Museum of Zoology, Cambridge and the Finnish Museum of Natural History, Helsinki, Finland for loans of specimens. facilitated by Martha Richter, Rob Asher, Matt Lowe and Martti Hildén. For assistance with laboratory work and materials, we thank Tom Davies, Wendy Dirks, Dani Schmidt, Remmert Schouten, Pedro Viegas, John Cunningham & Duncan Murdock. Discussions: Roger Benson, Chris Dean, Wendy Dirks, Jim Hopson, Fabien Lafuma, Thomas Martin, Rachel O’Meara, Stephen Naji, Tanya Smith and Emily Rayfield.
Author information
Authors and Affiliations
Contributions
I.J.C. and P.G.G. conceived and designed the project. E.N., P.G.G., P.B., K.R. and I.J.C. selected, prepared and curated specimens. E.N., P.G.G., P.B., V.F., D.H., T.K., A.K., A.P., P.S., H.S., P.T., B.Z.-P. and I.J.C. performed the synchrotron experiments. E.N., A.K. and I.J.C. performed the microCT experiments. E.N. processed and E.N., C.N. and K.W. analysed the synchrotron data. E.N. and I.J.C. analysed the micro-CT data. E.N., P.G.G. and I.J.C. discussed the interpretations. E.N. wrote the first draft and created all figures; E.N., P.G.G. and I.J.C. wrote the manuscript; all authors provided a critical review of the manuscript and approved the final draft. Authors M.J.B., V.F., N.J.G., D.H., J.J., T.K., A.K., C.N., A.P., K.R., K.R.B., P.S., H.S., P.T., K.W. and B.Z.-P. contributed equally to this work and are listed in alphabetical order. This article originated as a Master’s thesis (University of Bristol), then a PhD thesis (University of Southampton), performed by E.N. and supported and supervised by P.G.G., P.S., N.J.G., J.J., K.R.B., M.J.B. and I.J.C.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Communications thanks Zhe-Xi Luo and the other anonymous reviewer for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Newham, E., Gill, P.G., Brewer, P. et al. Reptile-like physiology in Early Jurassic stem-mammals. Nat Commun 11, 5121 (2020). https://doi.org/10.1038/s41467-020-18898-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-020-18898-4
This article is cited by
-
Global latitudinal gradients and the evolution of body size in dinosaurs and mammals
Nature Communications (2024)
-
New evidence from high-resolution computed microtomography of Triassic stem-mammal skulls from South America enhances discussions on turbinates before the origin of Mammaliaformes
Scientific Reports (2024)
-
Jurassic fossil juvenile reveals prolonged life history in early mammals
Nature (2024)
-
Remarks on the functional morphology of the earliest mammals
Mammalian Biology (2024)
-
Agency as an Inherent Property of Living Organisms
Biological Theory (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
