
Abstract
Core-fucosylation is an essential biological modification by which a fucose is transferred from GDP-β-L-fucose to the innermost N-acetylglucosamine residue of N-linked glycans. A single human enzyme α1,6-fucosyltransferase (FUT8) is the only enzyme responsible for this modification via the addition of an α-1,6-linked fucose to N-glycans. To date, the details of substrate recognition and catalysis by FUT8 remain unknown. Here, we report the crystal structure of FUT8 complexed with GDP and a biantennary complex N-glycan (G0), which provides insight into both substrate recognition and catalysis. FUT8 follows an SN2 mechanism and deploys a series of loops and an α-helix which all contribute in forming the binding site. An exosite, formed by one of these loops and an SH3 domain, is responsible for the recognition of branched sugars, making contacts specifically to the α1,3 arm GlcNAc, a feature required for catalysis. This information serves as a framework for inhibitor design, and helps to assess its potential as a therapeutic target.
Similar content being viewed by others

Introduction
Core-fucosylation is an essential biological modification of the N-glycan core in eukaryotes (except for plants and fungi) that is performed by the inverting α1,6-fucosyltransferase (FUT8)1. It is the most common type of fucose modification and occurs in the Golgi apparatus. FUT8 transfers a n l-fucose residue from GDP-β-l-fucose (GDP-Fuc) onto the innermost GlcNAc of N-glycan to form an α1,6-linkage2. Gene knockout of FUT8 in mice led to early postnatal death, severe growth retardation and emphysema-like changes in the lung, and revealed that this modification is crucial for the activation of growth factor receptors1,3. FUT8 is also involved in cell adhesion and cell migration processes by influencing turnover and expression levels of E-cadherin4 and the activity of α3/β1 integrin5. Core-fucose also effects N-glycan conformation by favoring an extended orientation of the α1,6 arm6. This can lead to subtle changes in glycan recognition by lectin receptors7. However, core-fucosylation is likely most known for its capacity to adjust the humoral immune response by altering antibody-dependent cellular cytotoxicity (ADCC). Thus, different strategies for reducing core-fucosylation in therapeutic monoclonal antibodies have been developed to enhance ADCC8. Notably, FUT8 is up-regulated in numerous types of cancer, suggesting that blocking its activity could be a promising strategy for improving anti-tumor immune responses9.
The acceptor specificity of FUT8 requires the presence of a terminal GlcNAc moiety on the α1,3 arm of the N-glycan but shows a higher degree of flexibility on the α1,6 arm10. This substrate preference does not necessarily demand the presence of a peptide/protein10. On the contrary, FUT8 also fucosylates high mannose N-glycans lacking a terminal GlcNAc moiety on the α1,3 arm. However, in these cases a peptide/protein moiety attached to the first GlcNAc via an N-glycosidic linkage11 is necessary.
The crystal structure of the human FUT8 (HsFUT8) apo form has been previously reported revealing that FUT8 is composed of a multi-domain enzyme that contains an N-terminal coiled-coil domain, a catalytic domain, which adopts a GT-B fold, and a C-terminal SH3 domain12. However, the lack of solved ternary complexes has impeded to obtain mechanistic insights into the glycosyl transfer reaction or reveal the molecular basis of the requirement for a terminal GlcNAc moiety on the α1,3 arm for optimal catalysis. Here, we have captured a ternary complex formed between the human FUT8 (HsFUT8), GDP and the biantennary complex N-glycan G0 (see below) by X-ray crystallography, uncovering the molecular basis of FUT8 catalysis and recognition of acceptor substrates.
Results
Core-fucosylation of synthetic N-glycans by HsFUT8
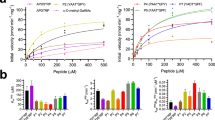
HsFUT8 was successfully cloned, expressed and purified to homogeneity (Methods) to enable functional and structural characterization. The activity of HsFUT8 on 5-aminopentyl G0 was monitored by mass spectrometry that showed a mass increase of 146 Da corresponding to the incorporation of a fucose moiety into the glycan in the presence of GDP-Fuc (Supplementary Fig. 1). Then, we evaluated the HsFUT8 acceptor substrate specificity on a microarray of synthetic N-glycans by expanding an earlier array version used by us in the study of C. elegans FUT8 (CeFUT8)13,14 (Fig. 1, Supplementary Table 1). The HsFUT8 showed identical substrate acceptor preferences to the reported ones for CeFUT8 (Fig. 2), and verified prior reports describing the importance of a terminal GlcNAc moiety in the α1,3 arm as a requirement for catalysis and a great promiscuity towards modifications on the α1,6 arm10.

a Pictogram representation of N-glycan structures included on the glycan array. b Specification of glycosidic bond configurations. c Monosaccharide symbol nomenclature according to Consortium of Functional Glycomics recommendations. See the following link for further information, http://www.functionalglycomics.org/static/consortium/Nomenclature.shtml.

a Aleuria aurantia lectin (AAL-555) binding before and after incubation with recombinant HsFUT8 at two different concentrations (650 nM and 3.5 μM) on a selection of glycan structures. b AAL-555-binding profile on a full glycan array before treatment with recombinant HsFUT8. c AAL-555-binding assay after treatment with recombinant HsFUT8 at 650 nM enzyme concentration and d AAL-555-binding assay after treatment with recombinant HsFUT8 at 3.25 μM enzyme concentration. Histogram bars show the average fluorescence RFU (relative fluorescence units) values for four replicate spots on the array. Error bars represent the standard deviation of the average RFU values on the same microarray. N-glycans that show binding towards AAL after incubation with HsFUT8 are highlighted in blue.
Architecture of the HsFUT8-GDP-G0 complex
Further, we successfully obtained C2 crystals of HsFUT8 in a complex with GDP and G0 glycan (Table 1). The resulting crystals allowed us to solve the structure at high resolution (1.95 Å) and interpret the density map (Table 1). The asymmetric unit (AU) of C2 crystals contains two molecules of HsFUT8 that are arranged as dimers with two molecules from the neighboring AU by direct contact between their corresponding N-terminal coiled-coil domains (Fig. 3). The root mean square deviation (RMSD) between both molecules belonging to chain A and B in the AU is 0.44 Å on 452 equivalent Cα atoms (Fig. 4; hereafter we will only discuss molecule A because it does not contain any disordered regions). As expected, the analysis of the HsFUT8 structure with the DALI server15 revealed structural homology to two fucosyltransferases, namely the Arabidopsis thaliana FUT1 (AtFUT1; PDB entries 5KX6, 5KOP, 5KOR, 5KWK and 5KOE16,17) and the human and C. elegans PoFUT2 (HsPoFUT2 and CePoFUT2; PDB entries 4AP5, 4AP6 and 5FOE18,19,20). Although FUT8 is very distant to AtFUT1 and HsPoFUT2/CePoFUT2 in terms of acceptor substrates, the server rendered good scores implying that they superimposed fairly well (RMSDs of ~3.2 and ~5.0 Å between AtFUT1 and HsFUT8, and AtFUT1 and HsPoFUT2/CePoFUT2 crystal structures, respectively; the superimposed residues ranged from 429 to 459 residues).

One HsFUT8 molecule (A in gray) from one asymmetric unit forms a dimeric structure with another molecule (B* in blue) from the other asymmetric unit* through their corresponding coiled-coil domains.

a Ribbon structure of HsFUT8 with GDP (orange carbon atoms) and G0 (green carbon atoms). The coiled-coil, catalytic and SH3 domains are colored in gray, red and orange, respectively. The interdomain α3 and loop β10–β11 are colored in blue and aquamarine, respectively. Disulfide bridges are indicated as yellow sulfur atoms. The C-terminal loop is colored in black. (inset) Electron density maps are FO–FC (blue) contoured at 2.2σ for GDP and G0. The labels of the sugar units (A–G) are also shown. b Surface representation of the HsFUT8-GDP-G0 complex. c Superposition of HsFUT8 apo (pink) and substrate-bound (gray) forms. This figure also depicts the conformational changes visualized in residues 365–378 and 428–444. These regions are represented as aquamarine (apo form) and blue (substrate-bound form).
The structure of HsFUT8 complexed to GDP and G0 depicts the three aforementioned domains and how they are connected with each other. While the catalytic domain is connected to the coiled-coil domain by α3, the SH3 domain is connected to the catalytic domain by the loop β10–β11 (Fig. 4a, Supplementary Fig. 2). While in general, densities for GDP and the rest of the G0 molecule are very well defined, we only observe partial density for the GlcNAc moiety on the α1,6 arm (GG0). Of special note is the anti-ψ conformation for the core-chitobiose GlcNAc moieties (glycosidic linkage involving units AG0–BG0) of the N-glycan in the bound state (see below). This conformer is less stable relative to the typical syn-ψ conformation found in solution21 (see inset in Fig. 4a). In addition, the Manα1-6Man linkage of G0 adopts an ‘extended gg’ conformer22, with ψ and ω values of 174.7° and 50.4°, respectively. The structure also reveals that GDP is partly buried and located exclusively in the catalytic domain, while the inner part and the α1,3/α1,6 arms of G0 are located in the catalytic domain and the exosite formed by the loop β10–β11 and SH3 domain, respectively (Fig. 4b). The presence of exosites for intimate binding to N-glycans was also observed in distant glycosyltransferases (GTs) and glycosyl hydrolases such as MGAT2 and MAN2A1, respectively23. Our structure also exhibits two striking conformational changes relative to the apo form (RMSD of 0.56 Å on 436 equivalent Cαs; Fig. 4c). Residues 428–444 (formed by a loop and α10) experience a large conformational change (the largest distance, close to 22 Å, was observed for Gly437 in both forms) that allows the partial closure of the GDP-Fuc-binding site and which protects the more hydrophobic part of the molecule (the guanosine moiety) from the solvent. Significantly, during the transition from the apo to the substrate-bound forms, a rearrangement of the α10 takes place in which α10apo-form (residue 436–442) becomes unstructured and another α10, comprising residues 430–435, is formed. Furthermore, the loop β6–α8 (residues 365–378) is partly disordered in the apo form and suffers a conformational change in the presence of both ligands, leading to an ordered loop that contributes to the formation of both the GDP-Fuc and the acceptor binding sites (Fig. 4c).
The sugar nucleotide-binding site
The HsFUT8-binding site is large and set up by the GDP-Fuc and the N-glycan binding sites (Fig. 5a, b). The guanine moiety of GDP establishes CH–π interactions with Ala436, Val450 and Val471 while the guanosine moiety is tethered via hydrogen bonds to Tyr250/His363/Asp453 side chains and Tyr220/Thr408 backbones. The pyrophosphate interacts with Gly221/Cys222/Gln470 backbones, Lys369/Arg365/Ser469 side chains and a glycerol moiety. The presence of positively charged residues like Arg365 and Lys369 is typically found in GT-B fold GTs, which do not bind to metal ions24. These residues replace the metal present in GT-A fold GTs and stabilize the pyrophosphate groups and the right conformations of sugar nucleotides for catalysis24. The glycerol moiety is a fucose mimic that occupies a similar position as found for fucose (see Methods, Fig. 5b, c, Supplementary Fig. 3), and further interacts with Arg365 and Glu373 side chains, and AG0 OH6, the site of fucosylation in G0 (Fig. 5b). The GDP-Fuc coordinates were obtained from the crystal structure of CePoFUT1 complexed to GDP-Fuc (PDB entry: 3ZY6 (ref. 25)) and the resulting HsFUT8-GDP-Fuc-G0 complex was further minimized using molecular mechanics (MM) calculations, as shown in Methods. Two hydroxyl groups of the glycerol molecule occupy positions that mimic fucose OH4 and the endocyclic oxygen. In addition, one of these two hydroxyl groups is also found in interaction distance of Arg365 and mimics the conserved interaction observed by the fucose endocyclic oxygen and Arg365 (Fig. 5b, c, Supplementary Fig. 3). Glu373 side chain further interacts with AG0 OH6 and Lys369 side chain, which helps stabilize the negative charge of Glu373 carboxylate. This could improve the catalytic base character of Glu373 for deprotonating the AG0 OH6 (Fig. 5a, b). The importance of Arg365, Lys369 and Glu373 for catalysis was previously confirmed by Ala substitutions by site-directed mutagenesis of these residues, which rendered the resulting mutants fully inactive12. The location of Arg365, Lys369 and Glu373 in the mobile loop β6–α8 demonstrates the pivotal relevance of this loop in binding to the ligands and catalysis. Markedly, structure of HsFUT8 with GDP-Fuc (see Methods) and G0 could demonstrate the inversion at the anomeric carbon of fucose with respect to GDP-Fuc (Fig. 5c). Therefore, these results are compatible with an SN2 single-displacement reaction mechanism, which is deployed by most inverting GTs24.

a Complete GDP-Fuc and G0 binding sites of the HsFUT8-GDP-G0 complex. The residues forming the GDP-Fuc/AG0/BG0 and CG0/DG0/EG0/FG0/GG0 binding sites are depicted as gray and aquamarine/orange carbon atoms, respectively. GDP, G0 and glycerol are shown as orange, green and blue carbon atoms, respectively. Hydrogen bond interactions are shown as dotted black lines. b, c Close-up view of the binding site region of the HsFUT8-GDP-G0 and HsFUT8-GDP-Fuc-G0 complexes showing the essential residues (Arg365, Lys369 and Glu373) that are major players in the plausible SN2 single-displacement reaction mechanism. Note the proximity and the orientation of the AG0 OH6 to the anomeric carbon (4.48 Å) which is compatible with the inversion of the configuration during the reaction.
The N-glycan-binding site
Unlike the intimate recognition of GDP by HsFUT8, G0 displays less contacts with the protein (Fig. 5a), in line with previous SPR data in which the binding of GDP-Fuc was ~40-fold stronger than the binding of G0 to the enzyme26 (Fig. 5a). AG0 is recognized through hydrogen bonds formed between AG0 OH1 and Gly217 backbone, AG0 acetamide NH and Asp295 side chain, and AG0 OH3 and Asp295/Lys216 side chains. BG0 is tethered via interactions formed between BG0 OH6 and Glu373 side chain, and BG0 acetamide carbonyl group and Gln470 side chain. The unusual anti-ψ conformation of AG0 and BG0, which is imposed by the enzyme, is a strict requirement for the AG0 OH6 fucosylation site facing GDP-Fuc (Fig. 5a). Catalysis can only take place in the anti-ψ conformation even though this conformation is less energetically favorable than the solution-phase syn-ψ conformation (−8.9 versus −9.5 kcal/mol, estimated by MM calculations for the disaccharide containing AG0–BG0 units).
The sugars forming the branching part of G0 (CG0, DG0, EG0, FG0 and GG0) are located in the exosite formed by the loop β10–β11 and the SH3 domain (Fig. 5a). CG0 establishes a CH–π interaction with Tyr498 and provides one hydrogen bond between CG0 OH4 and Asp495 side chain. DG0 is poorly recognized by a lonely hydrogen bond between DG0 OH6 and the Glu373 backbone. However, the terminal GlcNAc moiety in the α1,3 arm, EG0, is the most intimately recognized monosaccharide residue of all branched sugars. EG0 established three hydrogen bonds that are formed between EG0 OH6 and His535 side chain, EG0 OH2 and Asp494 backbone, and EG0 acetamide NH and Asp495 side chain. In addition, the EG0 acetamide methyl group is engaged in a hydrophobic interaction with Ala374 (Fig. 5a). The contacts that EG0 establishes with the enzyme could explain the strict requirement for a GlcNAc moiety in this position. This unit improves the binding of the α1,3 arm, and in turn, provides stabilization of the entire molecule in the bound state. On the contrary, the branching sugars on the α1,6 arm are far less well recognized, which provides a plausible explanation for the larger promiscuity towards the α1,6-mannose branch10. The FG0 endocyclic oxygen and FG0 OH4 establish two hydrogen bonds with Gln502 side chain and Val531 backbone, respectively, while other potential interactions between GG0 and HsFUT8 (GG0 carbonyl group and GG0 OH3 with Gly501 and Gln502 backbones, respectively) cannot be confirmed due to the less well-defined density in this sugar residue (Figs. 4a and 5a). The addition of a bisecting GlcNAc in G0 would preclude fucosylation by FUT8 because this residue is likely to impose a significant steric hindrance to the complex (Fig. 6), explaining why N-glycans with bisecting GlcNAc are not substrates of FUT8 (ref. 27).

N-glycans carrying a bisecting GlcNAc are not substrates of HsFUT8 likely due to a steric clash between the bisecting GlcNAc with Ala532 (distance of 4.46 Å between the Ala532 backbone and CG0 OH4). Note that the bisecting GlcNAc would be bound to CG0 via the CG0 OH4, and it is this GlcNAc who would have a steric clash with Ala352. It is also likely that the addition of the bisecting GlcNAc causes conformational changes to the neighboring branching sugar moieties, which could contribute to disrupt the binding to HsFUT8.
Discussion
FUT8 is the only enzyme responsible for core-fucosylation on mammalian N-glycoproteins. Despite many years of research on the enzymology and chemistry of FUT8, its reaction mechanism remained elusive and the requirement for a terminal GlcNAc moiety on the α1,3 arm of the N-glycan for optimal fucosylation was not well understood. The structure of the HsFUT8 apo form provided the architecture of this enzyme, which is formed by three different domains12. However, this structure did not offer any clues for the reaction mechanism or substrate recognition. Here, we have determined the crystal structure of a functional HsFUT8 in complex with GDP and G0. Intriguingly, the structure reveals that residues 436–442, in which one loop and α10 are present, and the loop β6–α8 suffer large conformational changes in the presence of the ligands forming the GDP-Fuc and acceptor binding sites. Though conformational changes of loops have been visualized in inverting and retaining GTs24,28,29, the motion here of the loop β6–α8 in the presence of ligands is unique because this loop brings in key residues that recognize both GDP and G0. In particular, Arg365, Lys369 and Glu373 contribute not only to the binding but also to catalysis, with Glu373 acting as the catalytic base for AG0 OH6. This is compatible with an SN2 single-displacement reaction mechanism. Generally, GT-A or GT-B inverting GTs following an SN2 inverting mechanism contain an amino acid acting as a catalytic base24 (typically Glu, Asp, or His). However, in most structures, these residues acting as catalytic residues are already located in the binding site in the absence of ligands24. Hence and to our knowledge, FUT8 exemplifies a unique case in which the catalytic base is brought to the binding site in the presence of ligands (note that the distance of Glu373 Cαs of the apo and substrate-bound form is ca. 18 Å; Fig. 7).

This figure also depicts the conformational changes visualized in Glu373 (in aquamarine and blue for the apo and substrate-bound form, respectively).
Previously and based on computational studies, HsFUT8 was suggested to follow an SN2 mechanism in which the β-phosphate was acting as the catalytic base26. Similarly, the β-phosphate acting as the catalytic base was proposed for the inverting CePoFUT125, though in this case an SN1 mechanism was suggested. In both enzymes, there was no basic residue present in the active site that could act as an assisting base. However, and based on our crystal structure, it is now clear that the authors could not predict the motion of the loop β6–α8, which brings the essential catalytic base Glu373. This demonstrates that it is imperative to obtain ternary complexes for these enzymes to elucidate their catalytic mechanism. Our SN2 inverting mechanism is also reinforced by the previous elucidation of ternary complexes for the closest structural homologs, AtFUT1 (ref. 17) and CePoFUT2 (ref. 18). For both enzymes, residues such as Asp and Glu, respectively, were also acting as the catalytic bases.
At the level of the N-glycan-binding site, four major structural features are responsible of the selectivity and recognition of HsFUT8 on N-glycans: (a) the less stable anti-ψ conformation for the core-chitobiose GlcNAc moieties of the N-glycan in the bound state is a pre-requisite for core-fucosylation, since it provides the AG0 OH6 in close contact to GDP-Fuc; (b) the CH–π interaction between Tyr498 and CG0 is likely crucial for a right orientation of the branching sugars; (c) the existence of an exosite formed by the loop β10–β11 and SH3 domain is responsible for branching sugar recognition and, in particular, for the α1,3 arm GlcNAc binding. SH3 domains are small protein domains of about 60 amino acid residues present in a large number of proteins and well-known for driving protein–protein interactions30. The fact that residues from the HsFUT8 SH3 domain recognize sugar moieties implies the broad versatility of these domains in recognition of different molecules; and (d) the most intimately recognition for the α1,3 arm GlcNAc explains why FUT8 requires the presence of a terminal GlcNAc moiety on the α1,3 arm of the N-glycan for optimal fucosylation. This ligand-binding model also explains our own data using HsFUT8 on a microarray of synthetic N-glycans and prior reports10,13,14. However, our structure does not offer clues on how FUT8 fucosylates less optimally high mannose N-glycans linked to glycoproteins and peptides that lack a terminal GlcNAc on the α1,3-branch and which will require further structural studies.
In conclusion, the structure reported here shows how FUT8 recognizes N-glycans and provides information on the enzymatic mechanism. Our structure may assist in the development of inhibitors with a potential value for cancer therapy or as an alternative for the production of non-fucosylated antibodies with enhanced ADCC.
Methods
Glycan preparation
For the preparation of the biantennary complex A2 N-glycan (named here as G0), sialyl glycopeptide (SGP) predominantly glycosylated with A2G2S2 glycan was isolated from hen's egg yolk according to a published procedure31 and further enzymatically and chemically processed. Treatment of the glycopeptide with acetic acid at 80 °C removed sialic acid residues32 and deglycosylation with PNGase F (Asparia Glycomics) produced a mix of mono and bis-galactosylated glycans33. The obtained mixture (110 mg, 66.8 µmol) was dissolved in 50 mM citric phosphate buffer (8 mL, pH = 5). Aspergillus niger β-galactosidase (8.9 U/mg, 14.2 mg, in 1260 µL buffer) and an aqueous solution of 0.02% NaN3 were added. The reaction mixture was incubated at 37 °C and the reaction progress monitored via MALDI-TOF MS. After complete degalactosylation, MeOH (5 mL) was added and the reaction mixture centrifuged at 4500 r.p.m. MeOH was removed under reduced pressure and the supernatant was loaded onto a Bond elute graphite cartridge (10 g) and eluted with H2O (250 mL), 25% MeOH/H2O (500 mL), 50% MeOH/H2O (250 mL). The collected fractions of 50% MeOH were analyzed by MALDI-TOF MS and fractions containing pure product were pooled and lyophilized. The pure product G0 was obtained as a white solid (69.1 mg, 52.5 µmol, 79%). 1H NMR (500 MHz, D2O, 298 K): δ 5.10 (s, 1H, αH-1GlcNAc-1), 5.02 (s, 1H, H-1αMan-2), 4.83 (s, 1H, H-1αMan-3), 4.68 (s, 1H, H-1βMan-1), 4.62–4.59 (m, 1H, βH-1GlcNAc-1), 4.54–4.50 (m, 1H, H-1βGlcNAc-2), 4.46 (d, J = 8.4 Hz, 2H, H-1βGlcNAc-3, H-1βGlcNAc-4), 4.16 (s, 1H, H-2αMan-2), 4.10 (s, 1H, H-2αMan-3), 4.02 (s, 1H, H-2αMan-1), 3.90–3.31 (m, 39H), 1.99 (s, 3H, CH3,GlcNAc), 1.96 (s, 6H, CH3,GlcNAc), 1.95 (s, 3H, CH3,GlcNAc). 13C NMR (126 MHz, D2O, 298 K): δ 174.8, 174.7, 174.6 (4× CO), 101.3 (C1βGlcNAc-2), 100.4 (C1βMan-1), 99.5 (C1βMan-2, C1βGlcNAc2, C1βGlcNAc3), 97.0 (C1βMan-3), 94.8 (βC1GlcNAc-1), 90.6 (αC1αGlcNAc-1), 79.6, 79.5, 79.2, 76.4 (C2Man-3), 76.3 (C2Man-1), 75.8, 74.3, 73.5, 73.4, 73.2, 72.8, 72.4, 72.0, 69.9 (C2Man-2), 69.2, 67.3, 65.7, 61.6, 60.6, 59.9, 55.3 (C2GlcNAc-4), 54.9 (C2GlcNAc-3), 53.6 (C2GlcNAc-1, C2GlcNAc-2), 22.3, 22.2, 21.8 (4× CH3). HRMS (MALDI-TOF): calcd for C50H84N4O36Na [M + Na]+ 1339.475, found 1339.515.
Activity assay on 5-aminopentyl G0
G0 containing a C5 linker at the anomeric position of the reducing end (5-aminopentyl G034) (1 nmol), guanosine 5′-diphospho-β-L-fucose (GDP-Fuc) (2 nmol) and HsFUT8 (18 µM) in Tris 25 mM, 150 mM NaCl pH = 7.5 were incubated at room temperature (rt) for 5 h. MALDI-TOF MS analysis of the reaction mixture employing a solution of 2,5-dihydroxybenzoic acid (DHB, 5 mg/mL in acetonitrile) as matrix showed the formation of fucosylated 5-aminopentyl G0. Control experiments lacking HsFUT8 and GDP-Fuc were performed showing the unaltered 5-aminopentyl G0 by MALDI-TOF MS (Supplementary Fig. 1).
Glycan array analysis
HsFUT8 screening on glycan microarrays was performed as previously described with slight modifications13. Each subarray was incubated with a solution (200 µL) containing two different concentrations of FUT8 (650 nM and 3.25 µM), GDP-Fuc (0.5 mM), MnCl2 (10 mM) in Tris 25 mM, 150 mM NaCl pH = 7.5 at rt overnight. The supernatant was removed and the slide was washed with PBS and water. The introduction of Fuc residues by the action of FUT8 was probed by incubation with Aleuria aurantia lectin (Vector laboratories, Burlingame, CA, USA) labeled with Alexa Fluor™ 555 NHS succinimidyl ester (Thermo Fisher, AAL-555, 60 µg/mL in binding buffer: Tris 25 mM, 150 mM NaCl pH = 7.5 containing 0.01% Tween-20) at rt for 1 h. The microarray slide was washed with binding buffer, and water and fluorescence was analyzed on an Agilent G265BA microarray scanner system (Agilent Technologies, Santa Clara, USA). Quantification was performed with ProScanArray® Express (Perkin Elmer, Shelton, USA) and Microsoft Excel software. The average of mean RFU values after background subtraction and standard deviation for four replicate spots was represented as histograms employing GraphPad Prism 6 software.
Expression and purification of HsFut8
The DNA sequence encoding amino acid residues 68–575 of the HsFUT8 was codon optimized and synthesized by GenScript (USA) for expression in HEK293 cells (see Supplementary Table 2 for the codon optimized sequence). The DNA, containing at the 5′-end a recognition sequence for KpnI, and at the 3′ end a stop codon and a recognition sequence for XhoI, was cloned into a modified pHLSec containing after the secretion signal sequence a 12xHis tag, a superfolder GFP35 and a Tobacco Etch Virus (TEV) cleavage site, rendering the vector pHLSec-12His-GFP-TEV-HsFUT8. Both the synthesis of the HsFUT8 construct and the engineered pHLSec together with the cloning of HsFUT8 into pHLSec-12His-GFP-TEV were performed by GenScript.
pHLSec-12His-GFP-TEV-HsFUT8 was transfected into HEK293F cell line (Thermo Fisher Scientific) as described below. Cells were grown in suspension in a humidified 37 °C and 8% CO2 incubator with rotation at 125 r.p.m. Transfection was performed at a cell density of 2.5 × 106 cell/mL in fresh media F17 serum-free media with 2% Glutamax and 0.1% P188. For each 150 mL of culture, 450 μg of the plasmid (1 μg/μL) was diluted to 135 µL with sterilized 1.5 M NaCl. This mixture was added to each 150 mL cell culture flask and incubated for 5 min in the incubator. After that, 1.35 mg of PEI-MAX (1 mg/mL) was mixed to 135 µL with sterilized 1.5 M NaCl and added to the cell culture flask. Cells were diluted 1:1 with pre-warmed media supplemented with valproic acid 24 h post-transfection to a final concentration of 2.2 mM. Cells were harvested 6 days post-transfection by spinning down at 300 × g for 5 min, after which the supernatants were collected and centrifuged at 4,000 × g for 15 min.
Supernatant was dialyzed against buffer A (25 mM TRIS pH 7.5, 300 mM NaCl) and loaded into a His-Trap Column (GE Healthcare). Protein was eluted with an imidazol gradient in buffer A from 10 mM up to 500 mM. Buffer exchange to 25 mM TRIS pH 7.5, 150 mM NaCl (buffer B) was carried out using a HiPrep 26/10 Desalting Column (GE Healthcare). TEV protease was then added in a ratio 1:50 (TEV:protein) to the fusion construct in order to cleavage the His-GFP. After 20 h of reaction at 18 °C, the cleavage was satisfactorily verified through SDS-PAGE.
TEV protease and GFP were later removed from the solution using a His-Trap Column (GE Healthcare), and isolated HsFUT8 was then loaded into a HiLoad 26/60 Superdex 75 Colum (GE Healthcare), previously equilibrated with buffer B. Quantification of protein was carried out by absorbance at 280 nm using his theoretical extinction coefficient (ε280 nmHsFut8 = 90,760 M−1 cm−1).
Crystallization and data collection
Crystals of the HsFUT8 were grown by sitting drop experiments at 18 °C by mixing 0.5 μL of protein solution (4.5 mg/mL HsFUT8, 5 mM GDP and 5 mM G0 in buffer B) with an equal volume of a reservoir solution (0.1 M carboxilic acids, 0.1 M buffer system 3 pH 8.5 and 30% precipitant mix 1 (Molecular Dimensions). The crystals were cryoprotected in mother liquor containing 30% glycerol and flash frozen in liquid nitrogen.
Structure determination and refinement
Diffraction data were collected on the synchrotron beamline I24 of the Diamond Light Source (Harwell Science and Innovation Campus, Oxfordshire, UK) at a wavelength of 0.97 Å and a temperature of 100 K. Data were processed and scaled using XDS36 and CCP4 (refs. 37,38) software packages. Relevant statistics are given in Table 1. The crystal structure was solved by molecular replacement with Phaser37,38 using the PDB entry 2DE0 as the template. Initial phases were further improved by cycles of manual model building in Coot39 and refinement with REFMAC5 (ref. 40). Further rounds of Coot and refinement with REFMAC5 were performed to obtain the final structure. The final model was validated with PROCHECK; model statistics are given in Table 1. The AU of the C2 crystal contained two molecules of HsFUT8. The Ramachandran plots for the HsFUT8-GDP-G0 show that 92.1%, 7.6%, 0.2% and 0% of the amino acids are in most favored, allowed, generously allowed and disallowed regions, respectively.
Molecular mechanics minimizations
GDP-Fuc was obtained from CePoFUT1 in complex with GDP-Fuc (PDB entry: 3ZY6 (ref. 25)) and then it was superimposed on our crystal structure. The calculations were carried out on the complexes using AMBER 18 package, which was implemented with ff14SB, GLYCAM06 and GAFF force fields. The complex was immersed in a water box with a 10 Å buffer of TIP3P water molecules and neutralized by adding explicit Na+. A two-stage geometry optimization approach was performed with a total of 5000 minimization steps and using the default settings of AMBER 18. The first stage minimizes only the positions of solvent molecules and ions, and the second stage is an unrestrained minimization of all the atoms in the system.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
References
Wang, X. et al. Dysregulation of TGF-beta1 receptor activation leads to abnormal lung development and emphysema-like phenotype in core fucose-deficient mice. Proc. Natl. Acad. Sci. USA 102, 15791–15796 (2005).
Schneider, M., Al-Shareffi, E. & Haltiwanger, R. S. Biological functions of fucose in mammals. Glycobiology 27, 601–618 (2017).
Wang, X. et al. Core fucosylation regulates epidermal growth factor receptor-mediated intracellular signaling. J. Biol. Chem. 281, 2572–2577 (2006).
Osumi, D. et al. Core fucosylation of E-cadherin enhances cell-cell adhesion in human colon carcinoma WiDr cells. Cancer Sci. 100, 888–895 (2009).
Zhao, Y. et al. Deletion of core fucosylation on alpha3beta1 integrin down-regulates its functions. J. Biol. Chem. 281, 38343–38350 (2006).
Nishima, W., Miyashita, N., Yamaguchi, Y., Sugita, Y. & Re, S. Effect of bisecting GlcNAc and core fucosylation on conformational properties of biantennary complex-type N-glycans in solution. J. Phys. Chem. B 116, 8504–8512 (2012).
Andre, S. et al. Substitutions in the N-glycan core as regulators of biorecognition: the case of core-fucose and bisecting GlcNAc moieties. Biochemistry 46, 6984–6995 (2007).
Pereira, N. A., Chan, K. F., Lin, P. C. & Song, Z. The “less-is-more” in therapeutic antibodies: afucosylated anti-cancer antibodies with enhanced antibody-dependent cellular cytotoxicity. MAbs 10, 693–711 (2018).
Keeley, T. S., Yang, S. & Lau, E. The diverse contributions of fucose linkages in cancer. Cancers (Basel) 11, 1241 (2019).
Calderon, A. D. et al. Substrate specificity of FUT8 and chemoenzymatic synthesis of core-fucosylated asymmetric N-glycans. Org. Biomol. Chem. 14, 4027–4031 (2016).
Yang, Q., Zhang, R., Cai, H. & Wang, L. X. Revisiting the substrate specificity of mammalian alpha1,6-fucosyltransferase reveals that it catalyzes core fucosylation of N-glycans lacking alpha1,3-arm GlcNAc. J. Biol. Chem. 292, 14796–14803 (2017).
Ihara, H. et al. Crystal structure of mammalian alpha1,6-fucosyltransferase, FUT8. Glycobiology 17, 455–466 (2007).
Serna, S., Yan, S., Martin-Lomas, M., Wilson, I. B. & Reichardt, N. C. Fucosyltransferases as synthetic tools: glycan array based substrate selection and core fucosylation of synthetic N-glycans. J. Am. Chem. Soc. 133, 16495–16502 (2011).
Echeverria, B. et al. Chemoenzymatic synthesis of N-glycan positional isomers and evidence for branch selective binding by monoclonal antibodies and human C-type lectin receptors. ACS Chem. Biol. 13, 2269–2279 (2018).
Holm, L. & Laakso, L. M. Dali server update. Nucleic Acids Res. 44, W351–W355 (2016).
Urbanowicz, B. R. et al. Structural, mutagenic and in silico studies of xyloglucan fucosylation in Arabidopsis thaliana suggest a water-mediated mechanism. Plant J. 91, 931–949 (2017).
Rocha, J. et al. Structure of Arabidopsis thaliana FUT1 reveals a variant of the GT-B class fold and provides insight into Xyloglucan fucosylation. Plant Cell 28, 2352–2364 (2016).
Valero-Gonzalez, J. et al. A proactive role of water molecules in acceptor recognition by protein O-fucosyltransferase 2. Nat. Chem. Biol. 12, 240–246 (2016).
Lira-Navarrete, E. & Hurtado-Guerrero, R. A perspective on structural and mechanistic aspects of protein O-fucosylation. Acta Crystallogr. F Struct. Biol. Commun. 74, 443–450 (2018).
Chen, C. I. et al. Structure of human POFUT2: insights into thrombospondin type 1 repeat fold and O-fucosylation. EMBO J. 31, 3183–3197 (2012).
Corzana, F. et al. The pattern of distribution of amino groups modulates the structure and dynamics of natural aminoglycosides: implications for RNA recognition. J. Am. Chem. Soc. 129, 2849–2865 (2007).
Canales, A. et al. Breaking pseudo-symmetry in multiantennary complex N-glycans using lanthanide-binding tags and NMR pseudo-contact shifts. Angew. Chem. Int. Ed. Engl. 52, 13789–13793 (2013).
Kadirvelraj, R. et al. Human N-acetylglucosaminyltransferase II substrate recognition uses a modular architecture that includes a convergent exosite. Proc. Natl Acad. Sci. USA 115, 4637–4642 (2018).
Moremen, K. W. & Haltiwanger, R. S. Emerging structural insights into glycosyltransferase-mediated synthesis of glycans. Nat. Chem. Biol. 15, 853–864 (2019).
Lira-Navarrete, E. et al. Structural insights into the mechanism of protein O-fucosylation. PLoS ONE 6, e25365 (2011).
Kotzler, M. P. et al. Donor assists acceptor binding and catalysis of human alpha1,6-fucosyltransferase. ACS Chem. Biol. 8, 1830–1840 (2013).
Tseng, T. H. et al. Substrate preference and interplay of fucosyltransferase 8 and N-acetylglucosaminyltransferases. J. Am. Chem. Soc. 139, 9431–9434 (2017).
Lira-Navarrete, E. et al. Dynamic interplay between catalytic and lectin domains of GalNAc-transferases modulates protein O-glycosylation. Nat. Commun. 6, 6937 (2015).
Albesa-Jove, D. & Guerin, M. E. The conformational plasticity of glycosyltransferases. Curr. Opin. Struct. Biol. 40, 23–32 (2016).
Kurochkina, N. & Guha, U. SH3 domains: modules of protein-protein interactions. Biophys. Rev. 5, 29–39 (2013).
Sun, B. et al. A simplified procedure for gram-scale production of sialylglycopeptide (SGP) from egg yolks and subsequent semi-synthesis of Man3GlcNAc oxazoline. Carbohydr. Res. 396, 62–69 (2014).
Varki, A. & Diaz, S. The release and purification of sialic acids from glycoconjugates: methods to minimize the loss and migration of O-acetyl groups. Anal. Biochem. 137, 236–247 (1984).
Seko, A. et al. Occurence of a sialylglycopeptide and free sialylglycans in hen’s egg yolk. Biochim. Biophys. Acta 1335, 23–32 (1997).
Serna, S., Etxebarria, J., Ruiz, N., Martin-Lomas, M. & Reichardt, N. C. Construction of N-glycan microarrays by using modular synthesis and on-chip nanoscale enzymatic glycosylation. Chemistry 16, 13163–13175 (2010).
Moremen, K. W. et al. Expression system for structural and functional studies of human glycosylation enzymes. Nat. Chem. Biol. 14, 156–162 (2018).
Kabsch, W. Xds. Acta Crystallogr. Sect. D Biol. Crystallogr. 66, 125–132 (2010).
Winn, M. D. et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. Sect. D Biol. Crystallogr. 67, 235–242 (2011).
Collaborative Computational Project, Number 4. The CCP4 suite: programsfor protein crystallography. Acta Crystallogr. D Biol. Crystallogr. 50, 760–763 (1994).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004).
Murshudov, G. N. et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D Biol. Crystallogr. 67, 355–367 (2011).
Acknowledgements
We thank the Diamond Light Source (Oxford, UK) synchrotron beamline I24 (experiment number mx20229-19). We thank ARAID, MEC (CTQ2013-44367-C2-2-P, BFU2016-75633-P, CTQ2017-90039-R, RTC-2017-6126-1 and RTI2018-099592-B-C21), Maria de Maeztu Units of Excellence Program from the Spanish State Research Agency (MDM-2017-0720) and Gobierno de Aragón (E34_R17 and LMP58_18) with FEDER (2014-2020) funds for ‘Building Europe from Aragón’ for financial support. The research leading to these results has also received funding from the FP7 (2007-2013) under BioStruct-X (grant agreement No. 283570 and BIOSTRUCTX_5186).
Author information
Authors and Affiliations
Contributions
R.H.-G. designed the crystallization construct and solved the crystal structure. A.G.-G. and L.C.-L. purified the enzyme and crystallized the complex. R.H.-G. refined the crystal structure. F.C. performed the molecular mechanics calculations. S.S., R.A. and N.C.R. performed the synthesis of G0 and the mass spectrometry experiments together with the microarray of synthetic N-glycans experiment. R.H.-G. wrote the article with mainly the contribution of N.C.R. and F.C. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source Data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
García-García, A., Ceballos-Laita, L., Serna, S. et al. Structural basis for substrate specificity and catalysis of α1,6-fucosyltransferase. Nat Commun 11, 973 (2020). https://doi.org/10.1038/s41467-020-14794-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-020-14794-z
This article is cited by
-
Structure and dynamics of the Arabidopsis O-fucosyltransferase SPINDLY
Nature Communications (2023)
-
FDW028, a novel FUT8 inhibitor, impels lysosomal proteolysis of B7-H3 via chaperone-mediated autophagy pathway and exhibits potent efficacy against metastatic colorectal cancer
Cell Death & Disease (2023)
-
Structural insights into mechanism and specificity of the plant protein O-fucosyltransferase SPINDLY
Nature Communications (2022)
-
Structural basis for the synthesis of the core 1 structure by C1GalT1
Nature Communications (2022)
-
FUT8-mediated aberrant N-glycosylation of B7H3 suppresses the immune response in triple-negative breast cancer
Nature Communications (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
