
Abstract
The term Blue Carbon (BC) was first coined a decade ago to describe the disproportionately large contribution of coastal vegetated ecosystems to global carbon sequestration. The role of BC in climate change mitigation and adaptation has now reached international prominence. To help prioritise future research, we assembled leading experts in the field to agree upon the top-ten pending questions in BC science. Understanding how climate change affects carbon accumulation in mature BC ecosystems and during their restoration was a high priority. Controversial questions included the role of carbonate and macroalgae in BC cycling, and the degree to which greenhouse gases are released following disturbance of BC ecosystems. Scientists seek improved precision of the extent of BC ecosystems; techniques to determine BC provenance; understanding of the factors that influence sequestration in BC ecosystems, with the corresponding value of BC; and the management actions that are effective in enhancing this value. Overall this overview provides a comprehensive road map for the coming decades on future research in BC science.
Similar content being viewed by others
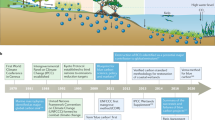


Introduction
Blue Carbon (BC) refers to organic carbon that is captured and stored by the oceans and coastal ecosystems, particularly by vegetated coastal ecosystems: seagrass meadows, tidal marshes, and mangrove forests. Global interest in BC is rooted in its potential to mitigate climate change while achieving co-benefits, such as coastal protection and fisheries enhancement1,2,3. BC has attracted the attention of a diverse group of actors beyond the scientific community, including conservation and private sector organizations, governments, and intergovernmental bodies committed to marine conservation and climate change mitigation and adaptation. The momentum provided by these conservation and policy actors has energized the scientific community by challenging them to address knowledge gaps and uncertainties required to inform policy and management actions.
The BC concept was introduced as a metaphor aimed at highlighting that coastal ecosystems, in addition to terrestrial forests (coined as green carbon), contribute significantly to organic carbon (C) sequestration1. This initial metaphor evolved to encompass strategies to mitigate and adapt to climate change through the conservation and restoration of vegetated coastal ecosystems1,2. As BC science consolidates as a paradigm, some aspects are still controversial; for instance, contrasting perspectives on the role of carbonate production as a component of BC4 and whether seaweed contributes to BC5,6. We propose an open discussion to refocus the current research agenda, reconcile new ideas with criticisms, and integrate those findings into a stronger scientific framework (Box 1). This effort will address the urgent need for refined understanding of the role of vegetated coastal ecosystems in climate change mitigation and adaptation.
There is, therefore, a need to establish a comprehensive research program on BC science that addresses current gaps while continuing to respond to immediate policy and managerial needs. Furthermore, this research program can inform policy directions based on new knowledge, thus playing a role in setting the management agenda and not simply responding to it. Here we identify, based on a broad effort by the leading research academics in BC science, key questions and challenges that need to be addressed to consolidate progress in BC science and inform current debate. We do so through three main steps. First, we briefly summarize the elements of BC science that represent the pillar of this research program. Second, we identify key scientific questions by first surveying the scientific community. Then we clustered these questions into common themes, which develop research goals and agendas. Last, we provide guidance as to how these questions can be best articulated into a new research agenda as a path for progress.
Scientists’ perspectives on the 10 key fundamental questions in BC science
We identified and selected scientists from among the leading and senior authors of the 50 most-cited papers on BC science (ISI Web of Science access date 22 June 2017), together with the participants in a workshop on BC organized at King Abdullah University of Science and Technology, Saudi Arabia, in March 2017. We did not attempt to identify any scientists’ area of specialisation to avoid bias. Among these authors, we surveyed those affiliated with academic or research institutions. A group of 50 scientists were asked to contribute from their perspective the top pending questions (up to 10) in BC science. Specifically, the invitees were asked to “Email your ten most important questions (or fewer) relevant to improving our understanding of blue carbon science and its application to climate change mitigation”. We did not ask scientists to prioritise their questions, or target any particular geographical area, but we did ask them to focus on mangrove, tidal marsh, macroalgal, and seagrass ecosystems. The answers received (35 total respondents, see Supplementary Note 1) and were then clustered into ten themes (by grouping questions that were similar) that were subsequently articulated into individual, overarching research questions:
Q1. How does climate change impact carbon accumulation in mature Blue Carbon ecosystems and during their restoration?
The impacts of climate change on BC ecosystems and their C stocks are dependent on the exposure to climate change factors. This is influenced by both the frequency and intensity of stressors, and the sensitivity and resilience of the ecosystem14. Question 1 reflects uncertainties associated with the rate and magnitude of climate change15,16,17 as well as uncertainties about the impacts of climate change on current and restored BC ecosystems, their rates of C sequestration and the stability of C stocks, which are likely to vary with past sea level history18, over geographic locations, among BC ecosystems, and within ecosystems.
BC ecosystems mainly occupy the intertidal and shallow water environments, where their distribution, productivity and rates of vertical accretion of soils are strongly influenced by sea level19,20 and the space available to accumulate sediment21. Thus, sea level rise ranks among the most important factors that will influence future BC stocks and sequestration. Sea level rise can result in BC gains, with increasing landward areal extent of ecosystems where possible22, and enhanced vertical accretion of sediments and C stocks18,23; and losses, with losses of ecosystem extent24, failure of restoration25, remineralization of stored organic matter26 that result in greenhouse gas emissions to the atmosphere (Table 1). Intense storms17, marine heat waves, 27, elevated CO228, and altered availability of freshwater29 have also all been implicated as important factors affecting the distribution, productivity, community composition and C sequestration of BC ecosystems over a range of locations (Table 1). Geographic variation in exposure to climate change is high. Rates of sea level rise and land subsidence30, which enhances relative rates of sea level rise, vary geographically18. Additionally, rates of temperature change and changes in the frequency of intense storms and rainfall vary regionally15,16,17. Geomorphic models have provided first pass assessments of the global vulnerability of BC ecosystems to sea level rise20,31, and for restoration success32, but local scale descriptors of changes in exposure of BC ecosystems to climate change and impacts on C stocks are often incomplete or missing. For instance, storm associated waves are important for determining the persistence and recruitment of BC ecosystem33, yet local assessments are not widely available.
Responses of adjacent ecosystems to climate change may influence the exposure and sensitivity of BC ecosystems and their C stocks to climate change. For example, degradation of coral reefs could increase wave heights within lagoons which may lead to losses of seagrass or mangroves within lagoons with rising sea levels as waves increase34, or decreases of carbonate sediments due to ocean acidification, may reduce the ability of some BC ecosystems to keep up with sea level rise35. Additionally, the sensitivity of BC ecosystems to climate change is also likely influenced by human activities in the coastal zone. For example, deterioration in water quality may increase the impacts of sea level rise on seagrass36 and decreased sedimentation from damming of rivers, hydrological modifications and presence of seawalls may negatively affect BC stocks in mangroves and tidal marshes20,31.
Q2. How does disturbance affect the burial fate of Blue Carbon?
The effect of disturbance on BC production and storage has become a topic of intense interest because of an increasing desire to protect or enhance this climate-related ecosystem service. There are three key issues, all beginning to be addressed by BC researchers, but requiring further study: (1) the depth in the soil profile to which the disturbance propagates, (2) the proportion of disturbed C that is lost as CO2, and (3) the extent to which issues 1 and 2 are context dependent. The first global estimates of potential losses of BC resulting from anthropogenic disturbance combined changes in the global distribution of BC ecosystems with simple estimates of conversion (remineralisation) of stored BC per unit area37. The estimated annual CO2 emission from the disturbance of BC ecosystems was estimated at 0.45 Petagrams CO2 globally37. The generalised assumptions necessary for such global assessments—e.g., remineralization within only the top 1 m of soil, and 100% loss of BC—provide little guidance at a local management scale and gloss over the variability of effects from different disturbance types38. This deficiency has led to a more nuanced theoretical framework accounting for the intensity of disturbance, especially whether the disturbance affects only the habitat-forming plant (e.g., clearing, eutrophication, light reduction, toxicity) or whether it also disturbs the soil (e.g., erosion, digging, reclamation)39,40. The duration of disturbance is another important predictor of disturbance effects on BC remineralisation because, over time, more soil BC is exposed to an oxic environment41.
We have a nascent understanding of the processes by which natural and human disturbances alter C decomposition. Die-off of below-ground roots and rhizomes in tidal marshes, for example, changes the chemical composition of BC and associated microbial assemblages, subsequently increasing decomposition and decreasing stored C (by up to 90% (ref. 42)). In seagrass ecosystems, exposing deeply buried sediments to oxygen triggered microbial breakdown of ancient BC43. At this stage, there is some evidence that disturbances can diminish BC stocks, for example: oil spills44, seasonal wrack deposition42, aquaculture45, eutrophication46, altered tidal flows46, and harvesting of fisheries resources38,47. Such knowledge is key for the construction of Emissions Factors for modelling. But examples in the literature are often specific for a particular disturbance or ecosystem setting, and do not yet offer the generalised understanding necessary to build a comprehensive framework guiding management projects. Finally, although there is widespread agreement that a changing climate directly affects BC production and storage, we recommend a clearer focus on the interacting effects of climate and direct anthropogenic disturbances.
Q3. What is the global importance of macroalgae, including calcifying algae, as Blue Carbon sinks/donors?
Macroalgae are highly productive (Table 2) and have the largest global area of any vegetated coastal ecosystem48. Yet only in a relatively few cases have macroalgae been included in BC assessments. Unlike angiosperms, which grow on depositional soils2, macroalgae generally grow on hard or sandy substrata that have no or only limited C burial potential6. However, a recent meta-analysis has estimated that macroalgae growing in soft sediments have a global C burial rate of 6.2 Tg C yr−1 (ref. 6), which is comparable to the lower range of estimates for tidal marshes. Furthermore, several studies show that macroalgae act as C donors3,6,49,50,51, where detached macroalgae are transported by currents, and deposited in C sinks beyond macroalgae habitats. Recent first-order estimates have suggested that up to 14 Tg C yr−1 of macroalgae-derived particulate organic C is buried in shelf sediments and an additional 153 Tg C yr−1 is sequestered in the deep ocean6. These calculations suggest that macroalgae may be supporting higher global C burial rates than seagrass, tidal marshes, and mangroves combined. This research highlights that if we are to incorporate macroalgal systems into BC assessments we need a better understanding of the fate of C originating from these systems. Furthermore, if we are to scale up from local measurements of C-sequestration to the global level, more refined estimates of the global surface area of macroalgal-dominated systems are needed.
Most estimates of C-sequestration by marine vegetated ecosystems refer solely to organic C even though calcifying organisms are also important components of such ecosystems52. For calcifying algae, whether they serve as C-sinks or sources is debated4, especially where calcifying organisms form and become buried within seagrass meadows4,5. Carbonate production results in the release of 0.6 mol of CO2 per mol of CaCO3 precipitated53, suggesting that calcifying algae are sources of CO2 that counteract C-sequestration in these ecosystems. However, co-deposition of organic and inorganic C may also have interacting effects on C-sequestration4. Carbonate may help protect and consolidate organic C sediment deposits, and CO2 release from mineralization of organic matter may stimulate carbonate dissolution and hence, CO2 removal48,53,54. Burial of inorganic carbon in seagrass and mangrove ecosystems is also to a large extent supported by inputs from adjacent ecosystems rather than by local calcification. Furthermore, mass balances highlight that such Blue Carbon ecosystems are sites of net CaCO3 dissolution54. More studies are needed to assess the net effect of organic and inorganic C deposition on C sequestration in calcifying systems.
Q4. What is the global extent and temporal distribution of BC ecosystems?
Our attempts to upscale BC estimates and model changes across large spatial and temporal scales is hindered by poor knowledge of their current and recent-past global distributions. The best constrained areal estimates exist for mangroves, which occur in tropical and subtropical regions, generally where winter seawater isotherms exceed 20 °C55. Overall, the global spatial extent of mangroves, and patterns and drivers of their temporal change, are relatively well understood, especially when compared with other BC ecosystems. Still, Giri et al.56 estimated a global area of mangroves of ca. 140,000 km2 in the year 2000 and Hamilton and Casey57 83,495 km2 in 2000 and 81,849 km2 in 2012. Both studies used Landsat data but different methodologies. Mangroves occur in 118 countries worldwide, but ~75% of total coverage is located within just 15 countries, with ~23% found in Indonesia alone56. Total mangrove extent during the second half of the 20th century declined at rates 1–3% yr−1 mainly due to aquaculture, land use change and land reclamation58. There are uncertainties in the area of mangrove that are scrub forms and which are therefore often not considered as forests despite their importance in arid and oligotrophic settings and often their large soil C stocks59,60. Since the beginning of the 21st century, mangrove loss rates are 0.16–0.39% yr−1 (ref. 57), probably reflecting changes in aquaculture and conservation efforts.
Tidal marshes are primarily found in estuaries along coasts of Arctic, temperate and subtropical coastal lagoons, embayments, and low-energy open coasts, although they also occur in some tropical regions61. Woodwell et al.62 estimated global tidal marsh extent of 380,000 km2 using the fraction of global coastline occupied by estuaries and the assumption that ~20% of estuaries supported tidal marshes48. However, tidal marsh area has been mapped in only 43 countries (yielding a total habitat extent of ca. 55,000 km2), which represents just 14% of the potential global area63. Tidal marsh extent is well documented for Canada, Europe, USA, South Africa and Australia63,64,65 but remains unknown to a large extent in regions, including Northern Russia and South America. An historical assessment of 12 estuaries and coastal seas worldwide indicated that >60% of wetland coverage has been lost66 mostly due to changes in land use, coastal transformation and land reclamation61. The minimum global rate of loss of tidal marsh area is estimated at 1–2% yr−1 (ref. 67).
Despite the widespread occurrence of seagrass across both temperate and tropical regions, the global extent of seagrass area is poorly estimated48. The total global area was recently updated to 350,000 km2 (ref. 68), although estimates range from 300,000 (ref.) to 600,000 km2 (ref. 69), with a potential habitable area for seagrass of 4.32 million km2 (ref. 70). Available distribution data are geographically and historically biased, reflecting the imbalance in research effort among regions71, and most data have been collected since the 1980s72. The total global seagrass area has decreased by ~29% since first reported in 1879—with ~7-fold faster rates of decline since 1990 (ref. 72)—due to a combination of natural causes, coastal anthropogenic pressure and climate change73.
Producing accurate estimates of the global extent of BC ecosystems is therefore a prerequisite to assess their contribution in the global carbon cycle. In addition, given the fast rate of decline reported for many BC ecosystems, regular revision of these estimates is needed to track any changes in their global extent and importance. Extensive mapping, with particular focus on understudied areas that may support critical BC ecosystems, that combines acoustic (i.e., side scan sonar and multi-beam eco-sounder) and optical (i.e., aerial photography and satellite images) remote sensing techniques with ground truthing (by scuba diving or video images) should be undertaken to map and monitor their extent and relative change over time74.
Q5. How do organic and inorganic carbon cycles affect net CO 2 flux?
Even though BC ecosystems are significant Corg reservoirs, depending on Corg and Cinorg dynamics they could also be net emitters of CO2 to the atmosphere through air-water CO2 gas exchange75. For instance, in submerged BC ecosystems (i.e., seagrasses), Corg storage is not directly linked with the removal of atmospheric CO2 because the water column separates the atmosphere from benthic systems. BC science gaps exist in complex inorganic and organic biogeochemical processes occurring within the water column and determining CO2 sequestration functioning.
Photosynthesis lowers the CO2 concentration in surface water as dissolved inorganic C (DIC) is incorporated into Corg ((1) in Fig. 1), and respiration and remineralization increases the CO2 concentration ((2) in Fig. 1). Net autotrophic ecosystems would lower surface water CO2 concentration and be a direct sink for atmospheric CO276,77. Lowering of surface water CO2 concentration is facilitated if allochthonous Corg ((3) in Fig. 1) and DIC inputs ((4) in Fig. 1) are low. Reactions of the inorganic C (Cinorg) cycle can also change the CO2 concentration in surface water and therefore influence net exchange of CO2 with the atmosphere4,5,78. Formation of calcium carbonate minerals (calcification) results in an increase of CO2 in the water column ((5) in Fig. 1) while dissolution of carbonate minerals decreases CO2 ((6) in Fig. 1). These processes may critically affect air–water CO2 gas exchange. Although recent studies related to the role of BC in climate change mitigation are beginning to address the abundance and burial rate of Cinorg in soils4,5,54,78,79,80, studies investigating the full suite of key processes for air–water CO2 fluxes, such as carbonate chemistry and Corg dynamics in shallow coastal waters and sediments, are still scarce (but see76,77,81,82). In particular, relevance of carbonate chemistry to the overall spatio-temporal dynamics of Corg and Cinorg pools and fluxes (e.g., origin, fate, abundance, rate, interactions) and air–water CO2 fluxes is largely uncertain for BC ecosystems4.

Conceptual diagram showing the biogeochemistry of carbon associated with air-water CO2 exchanges. Blue lines indicate the processes that enhance the uptake of atmospheric CO2, and red lines indicate those that enhance the emission of CO2 into the atmosphere. The CO2 concentration in surface water is primarily responsible for determining the direction of the flux. The concentration of surface water CO2 is determined by carbonate equilibrium in dissolved inorganic carbon (DIC) and affected by net ecosystem production (the balance of photosynthesis, respiration, and remineralization), which directly regulate DIC (1 and 2), allochthonous particulate and dissolved organic carbon (Corg), particulate inorganic carbon (Cinorg), and DIC inputs from terrestrial systems and coastal oceans (3 and 4), net ecosystem Cinorg production (the balance of calcification and dissolution), directly regulating both DIC and total alkalinity (TA) (5, 6), and temperature (solubility of CO2). Calcification produces CO2 with a ratio (released CO2/precipitated Cinorg) of approximately 0.6 in normal seawater54
Therefore, in addition to Corg related processes occurring in sediments and vegetation, future BC science should also quantify other key processes, such as air-water CO2 fluxes and Corg and Cinorg dynamics in water, to fully understand the role of BC ecosystems in climate change mitigation83.
Q6. How can organic matter sources be estimated in BC sediments?
Coastal ecosystems, mangroves, seagrasses and tidal marshes, occupy the land-sea interface and are subject to convergent inputs of organic matter from terrestrial and oceanic sources as well as transfers to and from nearby ecosystems84. However, the most basic requirement of quantifying organic matter inputs, and differentiating between allochthonous and autochthonous sources of Corg, remains a challenge. This limitation has particular relevance because of interest in financing the restoration of coastal ecosystems through the sale of BC offset-credits85. Policy frameworks such as the Verified Carbon Standard Methodology VM0033 (ref. 86) stipulate that offset-credits are not allocated under the framework for allochthonous Corg because of the risk of duplicating C sequestration gains that may have been accounted for in adjacent ecosystems. New methods are emerging that have greater potential to quantify the contribution of different primary producers to sedimentary organic carbon in marine ecosystems87.
Natural abundance of stable isotopes, most commonly 13C, 15N and 34S, have been used to trace and quantify allochthonous and autochthonous Corg sources and their relative contributions to carbon burial. The costs are low, the methodology for sample preparation and analysis is relatively easy and the validity of the technique has been widely, and generally successfully tested88. However, the diversity of organic matter inputs can result in complex mixtures of Corg that are not well resolved based on the isotopic separation of the sources. Isotopic values of different species may be similar, or may vary within the same species with microhabitats, seasons, growth cycle or tissue type89,90.
The use of bulk stable isotopes must be improved by additionally analysing individual compounds with a specific taxonomic origin. Biomarkers such as lignin, lipids, alkanes and amino acids, have proven useful for separating multiple-source inputs in coastal sediments88,91. Leading-edge studies, using compound-specific stable isotopes, employ both natural and radiocarbon analyses, providing the added dimension of age to taxonomic specificity92,93. Oxygen and hydrogen stable isotopes could also be used to improve resolving power, but up to now they have been used mainly in foodweb studies and their utility in determining sedimentary sources in coastal systems still needs to be validated87. Studies using both bulk and compound-specific isotopes must consider how decomposition may alter species-specific signatures89,90,94 Other, alternative fingerprinting techniques are emerging. The deliberate stable isotope labelling of organic matter and tracing its fate is a powerful approach that overcomes some of the limitations of natural abundance studies (e.g., source overlap), but has only looked at short-term Corg burial to-date95. The use of environmental DNA (eDNA) has been used to describe community composition in marine systems, but the potential to quantify the taxonomic proportions of plant sources in sediments has rarely been tested87,96.
Overall, projects using 13C and 15N stable isotopes will likely continue to dominate the investigation of organic matter sources, especially in simple two end member systems. While there is a growing suite of organic matter tracers, the ability to distinguish between specific blue carbon sources such as marsh vegetation and seagrass still remains a challenge. Sample size requirement, analytical time and cost implications, will be crucial in the selection of the most appropriate tracers for the characterisation and quantification of the molecular complexity in blue carbon sediments. In general, applications of most compound specific tracers have focused on environments other than those supporting blue carbon ecosystems88,93,97, and more work is needed to apply the same research tools to these systems. We recommend, wherever possible, that complementary methods such as compound-specific isotopes and eDNA that take advantage of methodological advances in distinguishing species contributions, be used in conjunction with bulk isotopes.
Q7. What factors influence BC burial rates?
BC ecosystems have an order of magnitude greater C burial rates than terrestrial ecosystems3. This high BC burial rate is a product of multiple processes that affect: the mass of C produced and its availability for burial; its sedimentation; and its subsequent preservation. A host of interacting biological, biogeochemical and physical factors, as well as natural and anthropogenic disturbance (see Q2), affect these processes. With respect to biological factors, it remains unclear how primary producer diversity and traits (e.g., biochemical composition, productivity, size and biomass allocation) influence BC98,99. However, it is likely that the suite of macrophytes present in BC ecosystems is critical to the mass of C available to be captured and preserved (as suggested for tidal marshes100). Equally, it is uncertain how fauna influence the production, accumulation or preservation of Corg via top-down processes such as herbivory38,101,102,103. Similarly, predators can regulate biomass, persistence and recovery of seagrasses, marshes and mangroves by triggering trophic cascades38. In addition, the functional diversity and activity of the microbial decomposer community, and how they vary with depth and over time, is only just beginning to be examined104 and will need to be linked to BC burial rates. Most likely this microbial community will be more important in defining the fate of Corg entering BC soils than its production and sedimentation.
The general effects of hydrodynamics on carbon sequestration in BC ecosystems are understood, yet there is much we still do not understand which could explain the variability in sequestration we see across BC ecosystems. We know that hydrodynamics, mediated by biological properties of BC ecosystems (e.g., canopy size and structure), affect particle trapping105,106,107 and, presumably, Corg sedimentation rates. For example, increasing density of mangrove stands positively affects affect wave attenuation, enhancing the accumulation of fine grained material108, which promotes Corg accumulation (silts and clays retain more Corg than sands109,110. However, significant variation in soil Corg has been observed within seagrass meadow111, pointing to complex canopy-hydrodynamic interactions which we do not understand but which could affect our ability to develop robust estimates of meadow-scale BC burial. For example, a study of restored seagrass meadow found strong positive correlations between Corg stocks and edge proximity leading to gradients in carbon stocks at scales of >1 km112. Elsewhere, flexible canopies have been shown to interact with wave dynamics, increasing turbulence near the sediment surface113. This could explain the loss of fine sediments, and presumably Corg, in low shoot density meadows compared to high density meadows114, with implications for carbon sequestration over time following restoration of BC ecosystems and the development of canopy density. Because these types of hydrodynamic interaction can affect the spatial and temporal patterns in carbon accumulation they need to be better understood in order to design stock and accumulation assessments and to predict the temporal development of stocks following management actions.
The basic biogeochemical controls on Corg accumulation within soils are understood (e.g., biochemical nature of the Corg inputs which vary among primary producers115,116,117 and the chemistry of their decomposition products)110, but it remains unclear what controls the stability of stored Corg in BC soils and whether these factors vary across ecosystems or under different environmental conditions (incl. disturbance). With the exception of one recent paper43, we know little about the Corg -mineral associations in BC ecosystems, how these affect the recalcitrance of soil Corg or whether specific forms are protected more by this mechanism than others, though this is clearly the case in other ecosystems118,119,120. Undoubtedly the anaerobic character of BC soils places a significant control on in situ rates of Corg decomposition and remineralisation. However, the time organic materials are exposed to oxygen before entering the anaerobic zone of BC soils will impact the quantity and nature of Corg as will the redox potential reached within the soil. The amount of time organic matter is exposed to oxygen explains the observation that Corg concentrations in tidal marshes globally are higher on coastlines where relative sea level rise has been rapid compared to those where sea level has been relatively stable18. Moreover, exposure of BC to oxygen has been recently shown trigger microbial attack, even ancient (5000-year-old) and chemically recalcitrant BC43. Enhancing our understanding of oxygen exposure times and critical redox potentials will help explain variations in Corg accumulation rates and preservation within different BC ecosystems.
From the above, there is increasing evidence that we do not understand the complex interactions among influencing environmental factors well enough to predict likely Corg stocks in soils, including temperature, hydrodynamic, geomorphic and hydrologic factors that can affect biogeochemical processes or mediate biological processes, and this leads to apparent contradictions. For example, the influence of nutrient availability on Corg stocks is unclear with one study reporting an increase in soil Corg stocks along a gradient of increasing phosphate availability121, another reporting no effect122, and yet others121,123 finding that increasing nutrient availability led to lower soil Corg. Some empirical studies have examined interactive effects or evoked them to explain difference in Corg stock101,124,125. However, these studies are rare and limited by the complexity or the interactions being examined. We conclude that gaining insights into these interactive effects is more likely to be advanced through modelling approaches.
Q8. What is the net flux of greenhouse gases between Blue Carbon ecosystems and the atmosphere?
BC ecosystems are generally substantial sources or sinks of greenhouse gases (GHGs) (CO2, CH4, N2O), though we cannot construct accurate global BC budgets due to uncertainties in net fluxes. The C budget is best constrained for mangroves, with mangroves globally taking up 700 Tg C yr−1 through Gross Primary Production, and respiring 525 Tg C yr−1 (75%) back to the atmosphere as CO2126. However, large uncertainty exists in budgets due to poorly constrained mineralization pathways linked to CO2 efflux119.
We lack robust global C budgets for other BC ecosystems due to insufficient empirical evidence127. For example, while we have estimated global soil Corg stocks128 and accumulation rates for seagrasses, this is insufficient to create a budget129 because we lack representative data on community metabolism and GHG fluxes, particularly for CH4 and N2O emissions. Thus, we need to better quantify sink/source balances, e.g., the net balance between primary production vs. emissions from ecosystem degradation and pelagic, benthic, forest floor and canopy respiration126. We also need to understand how source/sink dynamics change budgets over time and how environmental parameters affect GHG fluxes129,130, allowing us to estimate thresholds that flip BC ecosystems from GHG sinks to sources.
Budgets generally focus on CO2 fluxes, though we must better understand fluxes of other GHGs such as CH4 and N2O, and their contribution to the global BC budget131. Global estimates show that CH4 emissions can offset C burial in mangroves by 20% because CH4 has a higher global warming potential than CO2 on a per molecule basis132. CH4 emissions may also offset C burial in seagrasses, though these estimates have not been made. In contrast, some mangroves are N2O sinks133 which would enhance the value of the C burial as a means to mitigate climate change. Overall, CH4 and N2O biogeochemistry is understudied in BC ecosystems.
Finally, we must understand how GHG fluxes change as BC ecosystems replace each other, such as when mangroves expand onto marshes at their latitudinal limits134, or are planted on seagrass meadows in Southeast Asia. We also need to understand how emissions may change with loss of BC ecosystems. For example, it has been coarsely estimated that a 50% loss of seagrass would result in a global reduction in N2O emissions of 0.012 Tg N2O-N yr−1 and a 50% loss of mangroves would result in a global reduction in emissions of 0.017 Tg N2O-N yr−1 (ref. 130).
Q9. How can we reduce uncertainties in the valuations of Blue Carbon?
Studies into BC increasingly include a valuation aspect, focussed on coastal sites135 but more recently also including offshore sites136, showing a range of values for different ecosystems as depicted in Fig. 2. Differences in values are driven by differences in BC sequestration and storage capacity and/or potential avoided emissions through conservation and restoration of ecosystems. There is also variation in BC values due to uncertainties in the calculation of C sequestration and permanence of C storage, as is required for valuation. The wide range of C valuation methods, including social costs of C111, marginal abatement costs112, and C market prices, also enhances the uncertainty and variation in valuation estimates.

Estimates of the economic value of blue carbon ecosystems per hectare. Data from ref. 1 and references therein. Symbols and images are courtesy of the Integration and Application Network, University of Maryland Center for Environmental Science (ian.umces.edu/symbols/)
Valuation of BC enables its inclusion in policy and management narratives113, facilitating the comparison of future socio-economic scenarios, including mitigation and adaptation interventions137, and raises conservation interests as an approach to mitigate climate change and offset CO2 emissions2. For example, BC budgets can be incorporated into national greenhouse gas inventories138. Alternatively, demonstrable gains in C sequestration and/or avoided emissions through conservation and restoration activities can be credited within voluntary C markets or through the Clean Development Mechanism of the United Nations Framework Convention on Climate Change (UNFCCC)86. Voluntary market methodologies for BC ecosystems have been released within the American Carbon Registry139 and within the Verified Carbon Standard86, while some countries are developing BC-focussed climate change mitigation schemes that provide economic incentives. However, on the international scale, BC ecosystems have previously not been consistently incorporated into frameworks for climate change mitigation that offer economic reward for the conservation of C sinks, such as the REDD + program140, possibly as there was insufficient information for its inclusion. Avoiding degradation of mangroves, tidal marshes and seagrasses could globally offer up to 1.02 Pg CO2-e yr−1 in avoided emissions37. Developing countries with BC resources have the opportunity to use BC for the NDC, for example Indonesia, where BC contribution to reduce emissions could be as much as 0.2 Pg CO2-e yr−1 or 30% of national land-based emission while mangrove deforestation only contributes to 6% of national deforestation141.
To reduce uncertainty in BC values and encourage use of values in future policy and management, we recommend improved interdisciplinary research, combining ecological and economic disciplines to develop standardised approaches to improve confidence in the valuation of BC. Ideally this should be undertaken alongside studies which recognise the additional values of conserving BC ecosystems, for example the benefits generated from fisheries enhancement, nutrient cycling, support to coastal communities and their livelihoods2 and coastal protection, which is considered a cost-effective method compared to hard engineering solutions142.
Q10. What management actions best maintain and promote Blue Carbon sequestration?
Research over the past decade has improved estimates of C dynamics at a range of spatial scales. This has enabled modelling of potential emissions from the conversion of seagrass, mangrove and tidal marsh to other uses41, and estimates of rates of and hotspots for CO2 emissions resulting from ecosystem loss. The development of policy, implementation of management actions and the demonstration of BC benefits (including payments), however, are still in their infancy.
There are three broad management approaches to enhance C mitigation by BC ecosystems: preservation, restoration and creation. Preserving ecosystem extent and quality—for example, through legislative protection and/or supporting alternative livelihoods—has the two-fold benefit of avoiding the remineralisation of historically sequestered C, while also protecting future sequestration capacity. Preservation may include direct or indirect approaches to maintain or enhance biogeochemical processes, such as sedimentation and water supply46. Restoration pertains to a range of activities seeking to improve biophysical and geochemical processes—and therefore sequestration capacity—in BC ecosystems. Examples include passive and/or active reforestation of logged and degraded mangrove forests143; earthwork interventions to return aquaculture ponds to mangrove ecosystems141; and the restoration of hydrology to drained coastal floodplains144. Managed realignment is a particular option for creating or restoring tidal marshes as part of a strategy to achieve sustainable coastal flood defence together with the provision of other services, including C benefits145; other similar options include: regulated tidal exchange131 and beneficial use of dredged material146. Although restoration may re-establish C sequestration processes, it is important to note that it may not prevent large amounts of fossil C being lost following future disturbance or intervention. ‘No net loss’ policies have been now developed and applied to wetland ecosystems in many countries (e.g., USA and EU). These generally imply the creation of BC ecosystems to replace those lost through development. Such approaches should be treated with caution, however, since there is confusion about terminology141, lack of enforcement and limited capacity to recreate the qualities of pristine sites.
Tools for the accounting and crediting of C payments now exist for coastal wetland conservation, restoration and creation under the voluntary C market86,147. Several small-scale projects (e.g., Mikoko Pamoja in Kenya) are now using these frameworks to generate C credits with others projects in development148. Few jurisdictions have adopted their own mechanisms for the accounting and/or trading of BC, though some have undertaken preliminary research to identify BC policy opportunities149.
Technical, financial and policy barriers remain before local initiatives can be scaled-up to make large impacts—such as through national REDD + initiatives. Significant barriers include: biases in the geographic coverage of data; approaches for robust, site-specific assessment and prediction of some C pools (e.g., below-ground C and atmospheric emissions); high transaction costs; and ensuring that equity and justice are achieved. In addition, most demonstrated efforts are recent actions with little quantification of C mitigation benefits (or societal outcomes) beyond the scale of a few years.
Despite such barriers, we now have the fundamental knowledge to justify the inclusion of BC protection, restoration and creation in C mitigation mechanisms. While there remain knowledge gaps—both in science, policy and governance—these will partly be addressed through the effective demonstration, monitoring and reporting of existing and new BC projects.
Toward a research agenda on the role of vegetated coastal ecosystems on climate change mitigation and adaptation
The questions above are not short of challenges and therefore, provide ample scope for decisive experiments to be designed and conducted, current hypotheses to be rejected or consolidated and new ideas and concepts to unfold. Emerging questions that are not yet supported by robust observations and experiments, include, for example: the estimation of allochthonous C (organic and inorganic) contributions to BC, which remains challenging due to availability of markers able to quantitatively discriminate among the different carbon sources; and the net balance of GHG emissions, which remains challenging as it requires concurrent measurements across relevant time and spatial scales of all major GHGs (CO2, CH4, NO2), for which not a single estimate is available to-date. The core questions that capture much of current research efforts in BC science include the role of climate change on C accumulation, efforts to improve the precision of global estimates of the extent of BC ecosystems, factors that influence sequestration in BC ecosystems, with the corresponding value of BC, and the management actions that are effective in enhancing this value. The preceding text provides a summary of current research efforts and future opportunities in addressing these key questions.
Three questions are long-standing, controversial, and need resolution in order to properly constrain the BC paradigm. The first is the effect of disturbance on GHG emissions from BC ecosystems, where the initial assumption, that the top meter of the soil C stock is likely to be emitted as GHG following disturbance37,128, continues to be carried across papers without being challenged or verified. The second is whether macroalgae-C can be considered BC. The term BC refers to C sequestered in the oceans1, and the focus on seagrass, mangroves and tidal marshes is justified by the intensity of local C sequestration these ecosystems support. If macroalgae provide intense C sequestration, whether in the ecosystem or beyond, they need to be dealt with in this context. And the third controversy is whether carbonate accumulation in BC ecosystems render them potential sinks of CO2 following disturbance. It is clear that there are far too many key uncertainties4 to resolve this at the conceptual level, since empirical evidence to provide a critical test is as yet lacking. We propose that a research program including key observational and experimental tests designed to resolve the mass balance of carbonate (e.g., balance between allochthonous and autochthonous production and dissolution)—and then the coupling between BC ecosystems and the atmosphere—is needed. In the case of all three controversies, we believe that the positive approach to address these questions, is to pause the current discussion, which are largely rooted in the lack of solid, direct empirical evidence, and recognize that further science is required before any conclusion can be reached.
In summary, the overview of questions provided above portrays BC science as a vibrant field that is still far away from reaching maturity. Apparent controversies are a consequence of this lack of maturity and need to be resolved through high quality, scalable and reproducible observations and experiments. We believe the questions above inspire a multifarious research agenda that will require continued broadening the community of practice of BC science to engage scientists from different disciplines working within a wide range of ecosystems and nations.
Change history
08 November 2019
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Nellemann C., et al. (eds) Blue Carbon. A Rapid Response Assessment. United Nations Environment Programme (GRID-Arendal, 2009). This report was the first to use the term ‘blue carbon’
Duarte, C. M., Losada, I. J., Hendriks, I. E., Mazarrasa, I. & Marba, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Change 3, 961–968 (2013). Reviewed data on blue carbon burial, stocks, accretion rates and potential losses.
McLeod, E. et al. A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 9, 552–560 (2011). Identified key areas of uncertainly and specific actions needed to understand the role of vegetated coastal ecosystems as carbon sinks.
Macreadie Peter, I., Serrano, O., Maher Damien, T., Duarte Carlos, M. & Beardall, J. Addressing calcium carbonate cycling in blue carbon accounting. Limnol. Oceanogr. Lett. 2, 195–201 (2017). Argued that calcium carbonate cycling has been ignored in blue carbon offset schemes, but warrants serious attention.
Howard, J. et al. Clarifying the role of coastal and marine systems in climate mitigation. Front. Ecol. Environ. 15, 42–50 (2017).
Krause-Jensen D., Duarte C. M. Substantial role of macroalgae in marine carbon sequestration. Nat. Geosci. 9, 737 (2016). Demonstrated that macroalgal export can make an important contribution to deep sea carbon sequestration, where it can be sequestered from the atmosphere.
Smith, S. V. Marine macrophytes as a global carbon sink. Science 211, 838–840 (1981). Seminal paper on the role of marine macrophytes in sequestering CO2; suggested that the role of seagrasses were overlooked.
Duarte, C. M., Middelburg, J. J. & Caraco, N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences 2, 1–8 (2005). This paper showed that vegetated marine habitats are responsible for a previously unaccounted, 50% of C burial in ocean sediments.
Kennedy H., et al. in Supplement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories: Wetlands (eds Hiraishi, T., et al.) (IPCC, 2014).
Arias-Ortiz, A. et al. A marine heatwave drives massive losses from the world’s largest seagrass carbon stocks. Nat. Clim. Change 8, 338 (2018).
Marbà, N. et al. Impact of seagrass loss and subsequent revegetation on carbon sequestration and stocks. J. Ecol. 103, 296–302 (2015).
Macreadie, P. I. et al. Losses and recovery of organic carbon from a seagrass ecosystem following disturbance. Proc. Biol. Sci. 282, 1–6 (2015).
Atwood, T. B. et al. Global patterns in mangrove soil carbon stocks and losses. Nat. Clim. Change 7, 523 (2017).
IPCC. Climate Change. 2007. Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change (Intergovernmental Panel on Climate Change, Geneva, Switzerland, 2007).
Cabanes, C., Cazenave, A. & Le Provost, C. Sea level rise during past 40 years determined from satellite and in situ observations. Science 294, 840–842 (2001).
Hansen, J. et al. Global temperature change. Proc. Natl Acad. Sci. USA 103, 14288–14293 (2006).
Knutson, T. R. et al. Tropical cyclones andclimate change. Nat. Geosci. 3, 157–163 (2010).
Rogers, K. et al. Wetland carbon storage controlled by millennial-scale variation in relative sea-level rise. Nature 567, 91–95 (2019).
Kirwan, M. L. & Megonigal, J. P. Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504, 53–60 (2013).
Lovelock, C. E. et al. The vulnerability of Indo-Pacific mangrove forests to sea-level rise. Nature 526, 559–563 (2015).
Woodroffe, C. D., et al. Mangrove sedimentation and response to relative sea-level rise. Annu. Rev. Mar. Sci. 8, 243–266 (2016).
Schuerch, M. et al. Future response of global coastal wetlands to sea-level rise. Nature 561, 231–234 (2018).
Kelleway, J. J. et al. Seventy years of continuous encroachment substantially increases ‘blue carbon’ capacity as mangroves replace intertidal salt marshes. Glob. Change Biol. 22, 1097–1109 (2016).
Albert, S., et al. Winners and losers as mangrove, coral and seagrass ecosystems respond to sea-level rise in Solomon Islands. Environ. Res. Lett. 12, 094009 (2017).
Lee, S. Y., Hamilton, S., Barbier, E. B., Primavera, J. & Lewis, R. R. Better restoration policies are needed to conserve mangrove ecosystems. Nat. Ecol. Evol. 3, 870–872 (2019).
Ellison, J. C. Mangrove retreat with rising sea-level, Bermuda. Estuar. Coast Shelf Sci. 37, 75–87 (1993).
Wernberg, T. et al. An extreme climatic event alters marine ecosystem structure in a global biodiversity hotspot. Nat. Clim. Change 3, 78–82 (2013).
Reef, R. et al. The effects of elevated CO2 and eutrophication on surface elevation gain in a European salt marsh. Glob. Change Biol. 23, 881–890 (2017).
Asbridge, E., Lucas, R., Ticehurst, C. & Bunting, P. Mangrove response to environmental change in Australia’s Gulf of Carpentaria. Ecol. Evol. 6, 3523–3539 (2016).
Syvitski, J. P. M. et al. Sinking deltas due to human activities. Nat. Geosci. 2, 681–686 (2009).
Spencer, T. et al. Global coastal wetland change under sea-level rise and related stresses: the DIVA Wetland Change Model. Glob. Planet. Change 139, 15–30 (2016).
Balke, T. & Friess, D. A. Geomorphic knowledge for mangrove restoration: a pan-tropical categorization. Earth Surf. Process. Landf. 41, 231–239 (2016).
Leonardi, N., Ganju, N. K. & Fagherazzi, S. A linear relationship between wave power and erosion determines salt-marsh resilience to violent storms and hurricanes. Proc. Natl Acad. Sci. USA 113, 64–68 (2016).
Saunders, M. I. et al. Interdependency of tropical marine ecosystems in response to climate change. Nat. Clim. Change 4, 724–729 (2014).
Saderne, V. et al. Accumulation of carbonates contributes to coastal vegetated ecosystems keeping pace with sea level rise in an Arid Region (Arabian Peninsula). J. Geophys. Res. 123, 1498–1510 (2018).
Saunders, M. I. et al. Coastal retreat and improved water quality mitigate losses of seagrass from sea level rise. Glob. Change Biol. 19, 2569–2583 (2013).
Pendleton, L. et al. Estimating global “blue carbon” emissions from conversion and degradation of vegetated coastal ecosystems. PLoS ONE 7, e43542–e43542 (2012). Estimated that up to 1 billion tonnes of CO 2 is emitted each year to the atmosphere following destruction of blue carbon ecosystems.
Atwood, T. B. et al. Predators help protect carbon stocks in blue carbon ecosystems. Nat. Clim. Change 5, 1038–1045 (2015).
Bouma, T. J. et al. Identifying knowledge gaps hampering application of intertidal habitats in coastal protection: opportunities & steps to take. Coast. Eng. 87, 147–157 (2014).
Lovelock, C. E. et al. Assessing the risk of carbon dioxide emissions from blue carbon ecosystems. Front. Ecol. Environ. 15, 257–265 (2017).
Lovelock, C. E., Fourqurean, J. W. & Morris, J. T. Modeled CO2 emissions from coastal wetland transitions to other land uses: tidal marshes, mangrove forests, and seagrass beds. Front. Mar. Sci. 4, 1–11 (2017).
Macreadie, P. I., Hughes, A. R. & Kimbro, D. L. Loss of ‘blue carbon’ from coastal salt marshes following habitat disturbance. PLoS One 8, 1–8 (2013).
Macreadie, P. I., et al. Vulnerability of seagrass blue carbon to microbial attack following exposure to warming and oxygen. 686, 264–275 (2019).
Silliman, B. R. et al. Degradation and resilience in Louisiana salt marshes after the BP-deepwater horizon oil spill. Proc. Natl Acad. Sci. USA 109, 11234–11239 (2012).
Sidik, F. & Lovelock, C. E. CO2 efflux from shrimp ponds in Indonesia. PLoS One 8, e66329 (2013).
Macreadie, P. I. et al. Can we manage coastal ecosystems to sequester more blue carbon? Front. Ecol. Environ. 15, 206–213 (2017). Proposed three key management actions for maximising blue carbon sequestration within existing coastal vegetated ecosystems.
Coverdale, T. C. et al. Indirect human impacts reverse centuries of carbon sequestration and salt marsh accretion. PLoS One 9, e9396 (2014).
Duarte, C. M. Reviews and syntheses: hidden forests, the role of vegetated coastal habitats in the ocean carbon budget. Biogeosciences 14, 301–310 (2017).
Raven, J. A. The possible roles of algae in restricting the increase in atmospheric CO2 and global temperature. Eur. J. Phycol. 52, 506–522 (2017).
Trevathan-Tackett, S. M. et al. Comparison of marine macrophytes for their contributions to blue carbon sequestration. Ecology 96, 3043–3057 (2015).
Hill, R. et al. Can macroalgae contribute to blue carbon? An Australian perspective. Limnol. Oceanogr. 60, 1689–1706 (2015).
van der Heijden, L. H. & Kamenos, N. A. Reviews and syntheses: calculating the global contribution of coralline algae to total carbon burial. Biogeosciences 12, 6429–6441 (2015).
Smith, S. V. Parsing the Oceanic Calcium Carbonate Cycle: a Net Atmospheric Carbon Dioxide Source, or a Sink? (Association for the Sciences of Limnology and Oceanography (ASLO) L&O e-Books, 2013).
Saderne, V. et al. Role of carbonate burial in Blue Carbon budgets. Nat. Commun. 10, 1106 (2019).
Along, D. M. The Energetics of Mangrove Forests (Springer Science and Business Media BV, 2009).
Giri, C. et al. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 20, 154–159 (2011).
Hamilton, S. E. & Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 25, 729–738 (2016).
Valiela, I., Bowen, J. L. & York, J. K. Mangrove forests: one of the world’s threatened major tropical environments. Bioscience 51, 807–815 (2001).
Adame, M. F. et al. Carbon stocks of tropical coastal wetlands within the Karstic landscape of the Mexican Caribbean. PLoS One 8, e56569 (2013).
Lugo, A. E. Old-growth mangrove forests in the United States. Conserv Biol. 11, 11–20 (1997).
Adam, P. Saltmarshes in a time of change. Environ. Conserv 29, 39–61 (2002).
Woodwell, G.M., Rich, P.H., Mall, C.S.A. Carbon in the Biosphere. In: Woodwell, G.M., Pecan, E.V. (eds.) Proceedings of the 24th Brookhaven Symposium in Biology pp. 221–240 (USAEC, Springfield, Virginian, 1973).
McOwen, C. J. et al. A global map of saltmarshes. Biodiversity Data J. 55, e11764 (2017).
Chmura, G. L., Anisfeld, S. C., Cahoon, D. R. & Lynch, J. C. Global carbon sequestration in tidal, saline wetland soils. Glob. Biogeochem. Cycles 17, 1111 (2003).
Macreadie, P. I., et al. Carbon sequestration by Australian tidal marshes. Sci. Rep. www.nature.com/articles/srep44071 (2017).
Lotze, H. K. et al. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312, 1806–1809 (2006).
Duarte, C. M., Dennison, W. C., Orth, R. J. W. & Carruthers, T. J. B. The charisma of coastal ecosystems: addressing the imbalance. Estuaries Coasts 31, 233–238 (2008).
UNEP-WCMC. Global distribution of seagrasses (version 4.0). Fourth update to the data layer used in Green and Short (2003) (UNEP-WCMC, Cambridge, 2016).
Charpy-Roubaud, C. & Sournia, A. The comparative estimation of phytoplanktonic, microphytobenthic and macrophytobenthic primary production in the oceans. Mar. Microb. Food Webs 4, 31–57 (1990).
Gattuso, J. P. et al. Light availability in the coastal ocean: impact on the distribution of benthic photosynthetic organisms and their contribution to primary production. Biogeosciences 3, 489–513 (2006).
Short, F. T. et al. Extinction risk assessment of the world’s seagrass species. Biol. Conserv 144, 1961–1971 (2011).
Waycott, M. et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl Acad. Sci. USA 106, 12377–12381 (2009).
Orth, R. J. et al. A global crisis for seagrass ecosystems. Bioscience 56, 987–996 (2006).
Pham, D. T. et al. A review of remote sensing approaches for monitoring Blue Carbon ecosystems: mangroves, seagrasses and salt marshes during 2010–2018. Sensors 19, E1933 (2019).
Regnier, P. et al. Anthropogenic perturbation of the carbon fluxes from land to ocean. Nat. Geosci. 6, 597–607 (2013).
Maher, D. T. & Eyre, B. D. Carbon budgets for three autotrophic Australian estuaries: implications for global estimates of the coastal air-water CO2 flux. Glob. Biogeochem. Cycles 26, GB1032 (2012).
Tokoro, T. et al. Net uptake of atmospheric CO2 by coastal submerged aquatic vegetation. Glob. Change Biol. 20, 1873–1884 (2014).
Howard Jason, L., Creed Joel, C., Aguiar Mariana, V. P. & Fourqurean James, W. CO2 released by carbonate sediment production in some coastal areas may offset the benefits of seagrass “Blue Carbon” storage. Limnol. Oceanogr. 63, 160–172 (2017).
Mazarrasa, I. et al. Seagrass meadows as a globally significant carbonate reservoir. Biogeosciences 12, 4993–5003 (2015).
Fodrie, F. J., et al. Oyster reefs as carbon sources and sinks. Proc. Biol. Sci. 284, 20170891 (2017).
Watanabe, K. & Kuwae, T. How organic carbon derived from multiple sources contributes to carbon sequestration processes in a shallow coastal system? Glob. Change Biol. 21, 2612–2623 (2015).
Bauer, J. E. et al. The changing carbon cycle of the coastal ocean. Nature 504, 61–70 (2013).
Kuwae, T. et al. Blue carbon in human-dominated estuarine and shallow coastal systems. Ambio 45, 290–301 (2016).
Hyndes, G. A. et al. Mechanisms and ecological role of carbon transfer within coastal seascapes. Biol. Rev. 89, 232–254 (2013).
Murray, B., Pendleton, L., Jenkins, W. & Sifleet, S. Green Payments for Blue Carbon: Economic Incentives for Protecting Threatened Coastal Habitats (Nicholas Institute for Environmental Policy Solutions, Duke University, Durham, 2011).
Emmer, I., et al. Methodology for Tidal Wetland and Seagrass Restoration. in Verified Carbon Standard.VM0033 (2015) The first voluntary market methodology for blue carbon ecosystems
Geraldi, N. R. et al. Fingerprinting Blue Carbon: rationale and tools to determine the source of organic carbon in marine depositional environments. Front. Mar. Sci. 6, 263 (2019).
Bianchi, T. S., et al. Redox effects on organic matter storage in coastal sediments during the holocene: a biomarker/proxy perspective. Annu. Rev. Earth Planet. Sci. 44, 295–319 (2016).
Kramer, M. G., Lajtha, K. & Aufdenkampe, A. K. Depth trends of soil organic matter C:N and 15N natural abundance controlled by association with minerals. Biogeochemistry 136, 237–248 (2017).
Canuel, E. A. & Hardison, A. K. Sources, ages, and alteration of organic matter in estuaries. Annu. Rev. Mar. Sci. 8, 409–434 (2016).
Upadhayay, H. R. et al. Methodological perspectives on the application of compound-specific stable isotope fingerprinting for sediment source apportionment. J. Soils Sediment. 17, 1537–1553 (2017).
Wakeham, S. G. & McNichol, A. P. Transfer of organic carbon through marine water columns to sediments – insights from stable and radiocarbon isotopes of lipid biomarkers. Biogeosciences 11, 6895–6914 (2014).
Canuel, E. A. & Hardison, A. K. Sources, ages, and alteration of organic matter in estuaries. Annu. Rev. Mar. Sci. 8, 409–434 (2016).
Oreska Matthew, P. J., Wilkinson Grace, M., McGlathery Karen, J., Bost, M. & McKee Brent, A. Non‐seagrass carbon contributions to seagrass sediment blue carbon. Limnol. Oceanogr. 63, S3–S18 (2018).
Oakes, J. M. & Eyre, B. D. Transformation and fate of microphytobenthos carbon in subtropical, intertidal sediments: potential for long-term carbon retention revealed by 13C-labeling. Biogeosciences 11, 1927–1940 (2014).
Reef, R. et al. Using eDNA to determine the source of organic carbon in seagrass meadows. Limnol. Oceanogr. 62, 1254–1265 (2017).
Close, H. G. Compound-specific isotope geochemistry in the ocean. Annu. Rev. Mar. Sci. 11, 27–56 (2019).
Handa, I. T. et al. Consequences of biodiversity loss for litter decomposition across biomes. Nature 509, 218 (2014).
Chapin, F. S. Effects of plant traits on ecosystem and regional processes: a conceptual framework for predicting the consequences of global change. Ann. Bot. 91, 455–463 (2003).
Kelleway, J. J., Saintilan, N., Macreadie, P. I., Baldock, J. A. & Ralph, P. J. Sediment and carbon deposition vary among vegetation assemblages in a coastal salt marsh. Biogeosciences 14, 3763–3779 (2017).
Thomas, C. R. & Blum, L. K. Importance of the fiddler crab Uca pugnax to salt marsh soil organic matter accumulation. Mar. Ecol. Prog. Ser. 414, 167–177 (2010).
Johnson, R. A., Gulick, A. G., Bolten, A. B. & Bjorndal, K. A. Blue carbon stores in tropical seagrass meadows maintained under green turtle grazing. Sci. Rep. 7, 13545 (2017).
He, Q. & Silliman, B. R. Consumer control as a common driver of coastal vegetation worldwide. Ecol. Monogr. 86, 278–294 (2016).
Liu, S. L. et al. Sediment microbes mediate the impact of nutrient loading on blue carbon sequestration by mixed seagrass meadows. Sci. Total Environ. 599, 1479–1484 (2017).
Gacia, E. & Duarte, C. M. Sediment retention by a mediterranean Posidonia oceanica meadow: the balance between deposition and resuspension. Estuar. Coast Shelf Sci. 52, 505–514 (2001).
Hansen, J. C. R. & Reidenbach, M. A. Wave and tidally driven flows in eelgrass beds and their effect on sediment suspension. Mar. Ecol. Prog. Ser. 448, 271–287 (2012).
Wilkie, L., O’Hare, M. T., Davidson, I., Dudley, B. & Paterson, D. M. Particle trapping and retention by Zostera noltii: a flume and field study. Aquat. Bot. 102, 15–22 (2012).
Horstman, E. M., Dohmen-Janssen, C. M., Narra, P. M. F., van den Berg, N. J. F. & Siemerink, M. Hulscher SJMH. Wave attenuation in mangroves: a quantitative approach to field observations. Coast. Eng. 94, 47–62 (2014).
Keil, R. G. & Hedges, J. I. Sorption of organic matter to mineral surfaces and the preservation of organic matter in coastal marine sediments. Chem. Geol. 107, 385–388 (1993).
Burdige, D. J. Preservation of organic matter in marine sediments: controls, mechanisms, and an imbalance in sediment organic carbon budgets? Chem. Rev. 107, 467–485 (2007).
Ricart, A. M. et al. Variability of sedimentary organic carbon in patchy seagrass landscapes. Mar. Pollut. Bull. 100, 476–482 (2015).
Oreska, M. P. J., McGlathery, K. J. & Porter, J. H. Seagrass blue carbon spatial patterns at the meadow-scale. PLoS One 12, e0176630 (2017).
Abdolahpour, M., Ghisalberti, M., McMahon, K. & Lavery, P. S. The impact of flexibility on flow, turbulence, and vertical mixing in coastal canopies. Limnol. Oceanogr. 63, 2777–2792 (2018).
van Katwijk, M. M., Bos, A. R., Hermus, D. C. R. & Suykerbuyk, W. Sediment modification by seagrass beds: muddification and sandification induced by plant cover and environmental conditions. Estuar. Coast. Shelf Sci. 89, 175–181 (2010).
Trevathan-Tackett, S. M. et al. A global assessment of the chemical recalcitrance of seagrass tissues: Implications for long-term carbon sequestration. Front. Plant Sci. 8, 925 (2017).
Torbatinejad, N. M., Annison, G., Rutherfurd-Markwick, K. & Sabine, J. R. Structural constituents of the seagrass Posidonia australis. J. Agric. Food Chem. 55, 4021–4026 (2007).
Kogel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem 34, 139–162 (2002).
Yeasmin, S., Singh, B., Johnston, C. T. & Sparks, D. L. Organic carbon characteristics in density fractions of soils with contrasting mineralogies. Geochim. Cosmochim. Acta 218, 215–236 (2017).
Lehmann, J. & Kleber, M. The contentious nature of soil organic matter. Nature 528, 60–68 (2015).
Baldock, J. A. & Skjemstad, J. O. Role of the soil matrix and minerals in protecting natural organic materials against biological attack. Org. Geochem. 31, 697–710 (2000).
Armitage, A. R. & Fourqurean, J. W. Carbon storage in seagrass soils: long-term nutrient history exceeds the effects of near-term nutrient enrichment. Biogeosciences 13, 313–321 (2016).
Howard, J. L., Perez, A., Lopes, C. C. & Fourqurean, J. W. Fertilization changes seagrass community structure but not blue carbon storage: results from a 30-year field experiment. Estuaries Coasts 39, 1422 (2016).
Martinez-Crego, B., Olive, I. & Santos, R. CO2 and nutrient-driven changes across multiple levels of organization in Zostera noltii ecosystems. Biogeosciences 11, 7237–7249 (2014).
Janousek, C. N. et al. Inundation, vegetation, and sediment effects on litter decomposition in Pacific Coast tidal marshes. Ecosystems 20, 1296–1310 (2017).
Weiss, C. et al. Soil organic carbon stocks in estuarine and marine mangrove ecosystems are driven by nutrient colimitation of P and N. Ecol. Evol. 6, 5043–5056 (2016).
Alongi, D. M. Carbon cycling and storage in mangrove forests. Annu. Rev. Marine Sci. 6, 195–219 (2014).
Duarte, C. M., et al. Seagrass community metabolism: assessing the carbon sink capacity of seagrass meadows. Global Biogeochem. Cycles 24, GB4032 (2010).
Fourqurean, J. W. et al. Seagrass ecosystems as a globally significant carbon stock. Nat. Geosci. 5, 505–509 (2012).
Macreadie, P. I., Baird, M. E., Trevathan-Tackett, S. M., Larkum, A. W. D. & Ralph, P. J. Quantifying and modelling the carbon sequestration capacity of seagrass meadows—a critical assessment. Mar. Pollut. Bull. 83, 430–439 (2014).
Murray, R. H., Erler, D. V. & Eyre, B. D. Nitrous oxide fluxes in estuarine environments: response to global change. Glob. Change Biol. 21, 3219–3245 (2015).
Kroeger, K. D., Crooks, S., Moseman-Valtierra, S. & Tang, J. W. Restoring tides to reduce methane emissions in impounded wetlands: a new and potent Blue Carbon climate change intervention. Sci. Rep. 7, 11914 (2017).
Rosentreter, J. A., Maher, D. T., Erler, D. V., Murray, R. H. & Eyre, B. D. CH4 emissions partially offset ‘Blue Carbon’ burial in mangroves. Sci. Adv. 4, eaao4985 (2018).
Erler, D. V. et al. Applying cavity ring-down spectroscopy for the measurement of dissolved nitrous oxide concentrations and bulk nitrogen isotopic composition in aquatic systems: correcting for interferences and field application. Limnol. Oceanogr. Methods 13, 391–401 (2015).
Bianchi, T. S. et al. Historical reconstruction of mangrove expansion in the Gulf of Mexico: Linking climate change with carbon sequestration in coastal wetlands. Estuar. Coast Shelf Sci. 119, 7–16 (2013).
Barbier, E. B. et al. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 81, 169–193 (2011).
Barange, M. et al. The cost of reducing the North Atlantic Ocean biological carbon pump. Front. Mar. Sci. 3, 290 (2017).
van den Bergh, J. & Botzen, W. J. W. Monetary valuation of the social cost of CO2 emissions: a critical survey. Ecol. Econ. 114, 33–46 (2015).
Hiraishi, T. et al. 2013 Supplement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories: Wetlands. (IPCC, Switzerland, 2014).
Deverel, S., et al. Restoration of California Deltaic and Coastal Wetlands — American Carbon Registry. 1–186 (2017).
Scholz, I. & Schmidt, L. Reducing Emissions from Deforestation and Forest Degradation in Developing Countries: Meeting the Main Challenges Ahead. Briefing Paper 6. (Deutsches Institut für Entwicklungspolitik, German Development Institute, Bonn 2008).
Murdiyarso, D. et al. The potential of Indonesian mangrove forests for global climate change mitigation. Nat. Clim. Change 5, 1089–1092 (2015).
Narayan, S. et al. The effectiveness, costs and coastal protection benefits of natural and nature-based defences. PLoS One 11, e0154735 (2016).
Tamooh, F. et al. Below-ground root yield and distribution in natural and replanted mangrove forests at Gazi bay, Kenya. Ecol. Manag. 256, 1290–1297 (2008).
Howe, A. J., Rodrigues, J. F. & Saco, P. M. Surface evolution and carbon sequestration in disturbed and undisturbed wetland soils of the Hunter estuary, southeast Australia. Estuar. Coast. Shelf Sci. 84, 75–83 (2009).
Luisetti, T. et al. Coastal and marine ecosystem services valuation for policy and management: Managed realignment case studies in England. Ocean Coast. Manag. 54, 212–224 (2011).
Bolam, S. G. & Whomersley, P. Development of macrofaunal communities on dredged material used for mudflat enhancement: a comparison of three beneficial use schemes after one year. Mar. Pollut. Bull. 50, 40–47 (2005).
Plan Vivo. The Plan Vivo standard for community payments for ecosystem services programmes. Available at http://www.planvivo.org/wp-content/uploads/Plan-Vivo-Stan dard-2013.pdf.(2013).
Wylie, L., Sutton-Grier, A. E. & Moore, A. Keys to successful blue carbon projects: lessons learned from global case studies. Mar. Policy 65, 76–84 (2016).
Kelleway, J. et al. Technical Review of Opportunities for Including Blue Carbon in the Australian Government’s Emissions Reduction Fund. (Department of the Environment and Energy, Canberra, 2017).
Ware J. R., Smith S.V., Reaka-Kudla M.L. Coral reefs: sources or sinks of atmospheric CO2? Coral Reefs 11, 127–130 (1992)
Smith S.V., Mackenzie F.T. The Role of CaCO3 Reactions in the Contemporary Oceanic CO2 Cycle. Aquatic Geochemistry 22, 153–175 (2016)
Acknowledgements
P.I.M. and C.E.L. were supported by an Australian Research Council Linkage Project (LP160100242). C.M.D. was supported by baseline funding from King Abdullah University of Science and Technology. T.K. and K.W. were supported by JSPS KAKENHI (18H04156) and the Environment Research and Technology Development Fund (S-14) of the Ministry of the Environment, Japan. B.D.E. was supported by Australian Research Council grants DP160100248 and LP150100519. D.A.S. was supported by the UK Natural Environment Research Council (NE/K008439/1), and D.K.J. was supported by the CARMA project (8021-00222B), funded by the Independent Research Fund Denmark. Funding was provided to P.M. by the Generalitat de Catalunya (MERS, 2017SGR 1588) and an Australian Research Council LIEF Project (LE170100219). This work is contributing to the ICTA ‘Unit of Excellence’ (MinECo, MDM2015-0552). O.S. was supported by an ARC DECRA (DE170101524). N.M. was supported by the Spanish Ministry of Economy, Industry and Competitiveness (MedShift project). N.B. was supported by the UK Research Councils under Natural Environment Research Council award NE/N013573/1. J.W.F. was supported by the US National Science Foundation through the Florida Coastal Everglades Long-Term Ecological Research program under Grant No. DEB-1237517. R.S. had the support of FCT, project FCT UID/MAR/00350/2018. I.E.H. was supported by Ramon y Cajal Fellowship RYC2014-14970, co-funded by the Conselleria d’Innovació, Recerca i Turisme of the Balearic Government and the Spanish Ministry of Economy, Industry and Competitiveness. The University of Dundee is a registered Scottish charity, no. 015096. J.P.M. was supported by the Smithsonian Institution and the National Science Foundation Long-Term Research in Environmental Biology Program (DEB-0950080, DEB-1457100, DEB-1557009).
Author information
Authors and Affiliations
Contributions
P.I.M., A.A., J.A.R. and C.M.D. designed the study. P.I.M., A.A., J.A.R., N.B., R.M.C., D.A.F., J.J.K., H.K., T.K., P.S.L., C.E.L., D.A.S., E.T.A., T.B.A., J.B., T.S.B., G.L.C., B.D.E., J.W.F., J.M.H.-S., M.H., I.E.H., D.K.-J., D.L., T.L., N.M., P.M., K.J.M., P.J.M., D.M., B.D.R., R.S., O.S., B.R.S., K.W. and C.M.D. contributed to the writing and editing of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Macreadie, P.I., Anton, A., Raven, J.A. et al. The future of Blue Carbon science. Nat Commun 10, 3998 (2019). https://doi.org/10.1038/s41467-019-11693-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-019-11693-w
This article is cited by
-
Improving soil carbon estimates of Philippine mangroves using localized organic matter to organic carbon equations
Carbon Balance and Management (2024)
-
Antarctic krill sequester similar amounts of carbon to key coastal blue carbon habitats
Nature Communications (2024)
-
Anthropogenic impacts on mud and organic carbon cycling
Nature Geoscience (2024)
-
Blue carbon storage in a sub-Antarctic marine protected area
Scientific Reports (2024)
-
The sugar kelp Saccharina latissima II: Recent advances in farming and applications
Journal of Applied Phycology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
