
Abstract
Lakes and impoundments are an important source of methane (CH4), a potent greenhouse gas, to the atmosphere. A recent analysis shows aquatic productivity (i.e., eutrophication) is an important driver of CH4 emissions from lentic waters. Considering that aquatic productivity will increase over the next century due to climate change and a growing human population, a concomitant increase in aquatic CH4 emissions may occur. We simulate the eutrophication of lentic waters under scenarios of future nutrient loading to inland waters and show that enhanced eutrophication of lakes and impoundments will substantially increase CH4 emissions from these systems (+30–90%) over the next century. This increased CH4 emission has an atmospheric impact of 1.7–2.6 Pg C-CO2-eq y−1, which is equivalent to 18–33% of annual CO2 emissions from burning fossil fuels. Thus, it is not only important to limit eutrophication to preserve fragile water supplies, but also to avoid acceleration of climate change.
Similar content being viewed by others
Introduction
The importance of lakes and rivers in the global carbon cycle is well established1,2,3. Local, regional, and global emission estimates of the carbonic greenhouse gases (GHGs), methane (CH4) and carbon dioxide (CO2), are reported regularly1,2,4,5 with estimates of nitrous oxide (N2O), another potent GHG produced in aquatic systems, also occasionally reported6. One of the most recent estimates of global GHG emissions from lakes and impoundments7 found that while absolute emission of CO2 is 5–10 times more than that of CH4 and N2O (in Tg of carbon (C) or nitrogen (N) per year), about 72% of the climatic impact of GHG emissions (in CO2-equivalents) from lakes and impounded waters is due to CH4. This is because CH4 is up to 34-times more potent as a GHG than CO2 and is responsible for approximately 20% of the overall additional atmospheric radiative forcing observed since 17508.
This recent finding regarding the importance of aquatic CH4 emissions7 contradicts earlier reports that CH4 and CO2 contributed equally to the global warming potential of GHG emissions from lakes and impoundments2,4,6. These previous estimates, however, were based on a simplistic upscaling method of extrapolating average observed emission rates to global lake and impoundment surface area without regard for driving mechanisms. The use of this type of upscaling, rather than a process-based approach, may be the cause for the large uncertainties surrounding global aquatic CH4 emissions9.
While CH4 emission rates are known to be controlled by a wide range of factors including lake depth10 and sedimentation rates11, to name a few, incorporation of drivers into approaches for estimating CH4 emissions has been limited by the lack of world-wide data on these factors. Remote sensing approaches are beginning to fill data gaps, however, and global datasets are now available for lake size and productivity, two important drivers of CH4 emissions12. DelSontro et al.7 used these global datasets, along with >8000 GHG flux measurements, and modeled emission rates as a function of system productivity and lake size. They found, as did a smaller study of impoundments13, that not only was CH4 the most important GHG emitted from aquatic systems in terms of climate impact, but that it rises exponentially with lake and impoundment chlorophyll a (chla) concentration, a proxy for productivity7. This is consistent with multiple lake studies showing that CH4 emissions positively correlate with productivity variables such as total phosphorus (TP) and chla5,10,13,14,15,16. Ultimately, these relationships reflect the link between an increase in organic substrate and enhanced rates of methanogenesis in productive aquatic systems14,17,18. This is an important finding because the productivity of inland waters is projected to increase in the coming decades.
Three distinct mechanisms are expected to induce increases in aquatic productivity over the next century (Table 1, Supplementary Table 1). First, increased human populations (+37% by 2050, +50% by 210019) will augment the release of sewage and agricultural fertilizers to inland waters by an estimated 1.23×–1.97× (e.g., increase by up to a factor of 1.97, expressed as 1.97× hereafter) by 2050 and 1.41×–3.19× by 2100 20,21,22. On a global scale, the spatial distribution of agricultural nutrient use correlates with that of surface water23, presumably because sustained crop growth requires both nutrients and ample water; therefore, increased global nutrient use is likely to affect inland surface waters. Second, increased storms and runoff will enhance nutrient losses from land by a mean 1.14× over the same period24,25, further increasing nutrient delivery to inland waters. Third, warming surface waters will increase global aquatic primary production by ~1.30× by 2100 26. These increases in eutrophication and consequent CH4 production will be augmented by the 1.10× net global expansion in lakes and impoundment spatial extent2,26,27 expected over the 21st century, resulting in more CH4-emitting surface waters. A further increase we cannot assess is enhanced nutrient concentrations in warm regions resulting from increased evaporation, which may be regionally important26,28, but has not been estimated globally. Projected increases in lake eutrophication are supported by a recent report that the fraction of lakes in the US that are oligotrophic decreased from 25% of all surveyed lakes to 7% over just a 5-year period (2007–2012)29. Depending on the trajectory of human population growth and changes in climate and weather, multiplying nutrient effects by exacerbating factors (Table 1, Supplementary Table 1) shows that productivity of lakes and impoundments will likely increase 1.37×–3.10× by 2050 and 2.17×–4.91× by 2100.
Here, we use the relationship between CH4 emission rates and chla reported in DelSontro et al.7 (Fig. 1) to predict the effect of increased eutrophication of the earth’s lakes and impoundments on CH4 emissions. We simulated four levels of increased TP concentration (1.5× , 2× , 2.5× , and 3× that of current levels) that are conservatively within the magnitude of increase predicted by diverse authors and models through 2100 (Table 1). We further simulated a future where improved nutrient management results in TP concentrations 0.75× that of current levels. We calculated the corresponding changes in chla (0.8× , 1.3× , 1.7× , 2.0× , and 2.2× that of current levels) using a non-linear TP—chla relationship derived from published data30 (Supplementary Figure 1). These relationships were applied to the global lake and impoundment surface area reported in Downing et al.31. The current global distribution of chla in lakes and impoundments was estimated using satellite-based measurements of chla in 80,000 lakes around the world32. Our simulation results indicate that enhanced eutrophication will increase CH4 emissions from lakes and impoundments by 30–90% over the next century.

Relationships between CH4 flux rates and productivity measured as the concentration of chlorophyll a (chla) in the water column of lakes and impoundments. Solid lines are regression relationships. a Diffusive flux is the flux rate from the water to the atmosphere driven by super-saturation in the water column. b Ebullitive flux is that emitted by the formation of bubbles transported to the surface. c Total flux is the sum of both diffusive and ebullitive flux. Note: panel a shows only the relationship between diffusive CH4 flux and chla concentration, whereas the predictive model for diffusive flux includes both a lake size and chla effect (Table 3)
Results
Enhanced CH4 emissions due to future productivity increases
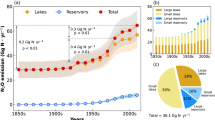
Our model indicates that future eutrophication of the world’s lakes and impoundments will increase diffusive, ebullitive, and total CH4 emissions (Fig. 2). Assuming a 3× increase in TP concentrations, diffusive emissions may increase from current levels (28 Tg C-CH4 y−1) up to 45 Tg CH4-C y−1, representing a 60% increase in emissions. Ebullition and total (diffusive + ebullitive) CH4 emission rates are more sensitive to enhanced eutrophication and will nearly double (mean increase of 1.86×) in a future environment with 3× greater nutrient loading to lakes and impoundments (Supplementary Table 2).

Simulated future global emissions of CH4 calculated at multiple levels of increased eutrophication driven by accelerated TP loading to lakes and impoundments (see Methods). The scenarios are based on a review of the published literature and reflect future TP concentrations that are 0.75, 1.5, 2, 2.5, and 3× greater (i.e., 0.75×, 1.5×, 2×, 2.5×, 3×) than current levels (1×), as indicated on x-axis. Simulated TP concentration was converted to chlorophyll a (chla) using published data (see Methods and Supplementary Figure 1) resulting in future chla concentrations that are 0.8, 1.3, 1.7, 2.0, and 2.2× that of current levels. CH4 emissions were predicted from lake surface area and/or chla
At current productivity levels, we find that total CH4 emissions (112 Tg C-CH4 y−1) are approximately 25% higher than the latest global estimates for lakes and impoundments (88.7 Tg C-CH4 y−1)9, which was based on a synthesis of measurements from mostly northern systems (>50°N) that used the traditional upscaling approach. Under a scenario of a tripling of nutrient loading, we could find the aggregate of global lakes and impoundments emitting more than double the current global estimate. More concerning yet is that even at the smallest increase in TP suggested by the literature (1.5×) we would see total global lake CH4 emissions rise to 141 Tg C-CH4 y−1, equivalent to global emissions from wetlands, which is the current single largest source of atmospheric CH4 (139 Tg C-CH4 y−1)9. A tripling of nutrient loading could result in total lake CH4 emissions being 1.5× higher than the current wetland emission estimate. It is possible, though unlikely, that improved nutrient management practices could reverse the eutrophication of lakes and impoundments, thereby reducing CH4 emissions. For example, a 25% reduction in global TP loading (0.75×) could prevent the emissions of 17 Tg CH4-C y−1 (Fig. 2, Supplementary Table 2). While a net reduction in nutrient loading is unlikely to occur on a global scale due to increasing population, it may be realized regionally or for individual water bodies and could be included in the cost/benefit analysis of nutrient management programs33.
Our literature review indicates that global TP loading of inland waters will increase over the 21st century, stimulating aquatic productivity and CH4 emissions. We find that these changes could result in total lake CH4 emissions having an atmospheric impact equivalent to 18–33% of that from current fossil fuel CO2 emissions (Table 2). In fact, the net increase in CH4 emissions due to projected nutrient loading alone would have an atmospheric effect more than increased CO2 emissions from land use change (i.e., conversion of forests to agriculture) or up to half of the global oceanic and land carbon sinks (Table 2 3,34). The projected CH4 emission increases reported here should be considered minimal because they do not account for the synergistic interaction between nutrients and water temperature on CH4 production rates39, future increases in impounded area27, or the effect of eutrophication on CH4 emissions from streams, rivers, small ponds, and wetlands. Eutrophication is considered to be one of the world’s most pressing environmental issues35 and, if it continues, will exacerbate global climate change.
Methods
Experimental design
We simulated future TP loadings to lakes and impoundments that are 0.75×, 1.5×, 2×, 2.5×, or 3× that of current loading, a conservative set of scenarios relative to our literature review which indicated that TP may increase nearly 5× relative to current levels by 2100 (Table 1, Supplementary Table 1 20,22,24,]36). Changes in TP concentration were translated to changes in chlorophyll a (chla) concentration using the non-linear TP—chla relationship derived from the data in McCauley et al.30 (Supplementary Figure 1) and an estimate of the current global chla distribution in lakes and impoundments. The global distribution of chla in lakes and impoundments was derived from satellite-based chla measurements of 80,000 lakes around the world32. We combined these data with an estimate of the size distribution of lakes and impoundments31 to generate a joint lake-size by productivity distribution for the worlds lakes and impoundments. Although several global lake-size distributions are available in the literature, all of them offer very similar conclusions. The data were aggregated into twenty 5 μg L−1 chla bins and nine lake-size bins ranging from 0.001 to >100,000 km2.
These TP loading scenarios (0.75×, 1.5×, 2×, 2.5×, or 3×) resulted in future chla concentrations that are, on average, 0.8×, 1.3×, 1.7×, 2.0×, and 2.2× that of the current global chla distribution. We propagated each of these chla distributions across the joint lake-size by chla distribution, resulting in five new distributions reflecting differing levels of eutrophication (Supplementary Data 1).
We calculated global CH4 emissions for each scenario using statistical models relating diffusive, ebullitive, and total CH4 emission rates to lake size and/or water column chla content (Table 3). First, we calculated the mean areal emission rate (mg CH4-C m−2 d−1) for each lake-size by chla bin. Next, we multiplied the predicted areal emission rate for each bin by the total water body surface area corresponding to the bin. Finally, we summed the emissions across bins and scaled the result to an annual emission estimate for each emission mechanism (i.e., diffusive, ebullitive, and total emissions).
The range of TP and chla concentrations in the most extreme 3× simulation (11–1669 µP L−1 and 5.7–216 µg L−1, respectively) were well within the range of values included in the literature used to parameterize the statistical models7 (Fig. 1). Statistical uncertainty in the model predictions was propagated through the calculations and is presented as 95% confidence intervals in Supplementary Table 2 and Supplementary Data 1.
The statistical models used to predict CH4 emission rates do not include an effect for water body origin (i.e., natural vs constructed) because of insufficient data coverage across the range of covariates for both system types. There is little evidence in the literature that suggests CH4 emission rates differ between natural lakes and impoundments, after system size and productivity are accounted for. Furthermore, impoundments constitute a small fraction of total inland water surface area31, therefore any systematic difference in the response of CH4 emission rate to size or productivity between natural lakes and impoundments will likely have little effect on cumulative global CH4 emissions from lentic waters.
Statistical analysis
The models used to upscale lake and impoundment methane (CH4) emissions are presented in DelSontro et al.7 and are reproduced in Table 3. The models predict CH4 emission rates (mg CH4-C m−2 d−1) from lake size (km2) and/or chla (µg L−1).
Data availability
All data used to generate Figure 1 and the models presented in Table 3 are available through figshare, an open-source data repository (https://doi.org/10.6084/m9.figshare.5220001). Joint lake-size and chla distribution tables for all modeled scenarios are available as Supplementary Data 1.
References
Cole, J. J. et al. Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems 10, 171–184 (2007).
Tranvik, L. J. et al. Lakes and reservoirs as regulators of carbon cycling and climate. Limnol. Oceanogr. 54, 2298–2314 (2009).
Ciais, P. & Sabine, C. in Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds Stocker, T. F. et al.) Ch. 6 (Cambridge University Press, Cambridge, UK, 2013).
Bastviken, D., Tranvik, L. J., Downing, J. A., Crill, P. M. & Enrich-Prast, A. Freshwater methane emissions offset the continental carbon sink. Science 331, 50 (2011).
Bastviken, D., Cole, J., Pace, M. & Tranvik, L. Methane emissions from lakes: dependence of lake characteristics, two regional assessments, and a global estimate. Glob. Biogeochem. Cycles 18, GB4009 (2004).
Soued, C., del Giorgio, P. A. & Maranger, R. Nitrous oxide sinks and emissions in boreal aquatic networks in Quebec. Nat. Geosci. 9, 116 (2016).
DelSontro, T., Beaulieu, J. J. & Downing, J. A. Greenhouse gas emissions from lakes and impoundments: upscaling in the face of global change. Limnol. Oceanogr. Lett. 3, 64–75 (2018).
IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds Stocker, T. F. et al.) (Cambridge University Press, New York, NY, USA, 2013).
Saunois, M. et al. The global methane budget 2000–2012. Earth Syst. Sci. Data 8, 697–751 (2016).
West, W. E., Creamer, K. P. & Jones, S. E. Productivity and depth regulate lake contributions to atmospheric methane. Limnol. Oceanogr. 61, S51–S61 (2016).
Maeck, A. et al. Sediment trapping by dams creates methane emission hot spots. Environ. Sci. Technol. 47, 8130–8137 (2013).
Rasilo, T., Prairie, Y. T. & Del Giorgio, P. A. Large-scale patterns in summer diffusive CH4 fluxes across boreal lakes, and contribution to diffusive C emissions. Glob. Change Biol. 21, 1124–1139 (2015).
Deemer, B. R. et al. Greenhouse gas emissions from reservoir water surfaces: a new global synthesis. Bioscience 66, 949–964 (2016).
Yang, H. et al. Greenhouse gas metabolism in Nordic boreal lakes. Biogeochemistry 126, 211–225 (2015).
Juutinen, S. et al. Methane dynamics in different boreal lake types. Biogeosciences 6, 209–223 (2009).
DelSontro, T., Boutet, L., St-Pierre, A., del Giorgio, P. A. & Prairie, Y. T. Methane ebullition and diffusion from northern ponds and lakes regulated by the interaction between temperature and system productivity. Limnol. Oceanogr. 61, S62–S77 (2016).
Whiting, G. J. & Chanton, J. P. Primary production control of methane emission from wetlands. Nature 364, 794–795 (1993).
West, W. E., McCarthy, S. M. & Jones, S. E. Phytoplankton lipid content influences freshwater lake methanogenesis. Freshw. Biol. 60, 2261–2269 (2015).
Samir, K. C. & Lutz, W. The human core of the shared socioeconomic pathways: population scenarios by age, sex and level of education for all countries to 2100. Glob. Environ. Change 42, 181–192 (2017).
Cole, J. J., Peierls, B. L., Caraco, N. F. & Pace, M. L. in Humans as Components of Ecosystems (eds McDonnell, M. J. & Pickett, S. T. A.) 141–157 (Springer, New York, NY, USA, 1993).
Caraco, N. F. & Cole, J. J. Human impact on nitrate export: an analysis using major world rivers. Ambio 28, 167–170 (1999).
Cordell, D., Drangert, J. O. & White, S. The story of phosphorus: global food security and food for thought. Glob. Environ. Change 19, 292–305 (2009).
Pacheco, F. S., Roland, F. & Downing, J. A. Eutrophication reverses whole-lake carbon budgets. Inland Waters 4, 41–48 (2014).
Sinha, E., Michalak, A. M. & Balaji, V. Eutrophication will increase during the 21st century as a result of precipitation changes. Science 357, 405–408 (2017).
Jeppesen, E. et al. Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J. Environ. Qual. 38, 1930–1941 (2009).
Downing, J. A. in Global Environmental Change (ed. Freedman, B.) 221–229 (Springer, Netherlands, 2014).
Zarfl, C., Lumsdon, A. E., Berlekamp, J., Tydecks, L. & Tockner, K. A global boom in hydropower dam construction. Aquat. Sci. 77, 161–170 (2015).
Jeppesen, E. et al. Climate change effects on nitrogen loading from cultivated catchments in Europe: implications for nitrogen retention, ecological state of lakes and adaptation. Hydrobiologia 663, 1–21 (2011).
Stoddard, J. L. et al. Continental-scale increase in lake and stream phosphorus: are oligotrophic systems disappearing in the United States? Environ. Sci. Technol. 50, 3409–3415 (2016).
McCauley, E., Downing, J. A. & Watson, S. Sigmoid relationships between nutrients and chlorophyll among lakes. Can. J. Fish. Aquat. Sci. 46, 1171–1175 (1989).
Downing, J. A. et al. The global abundance and size distribution of lakes, ponds, and impoundments. Limnol. Oceanogr. 51, 2388–2397 (2006).
Sayers, M. J. et al. A new method to generate a high-resolution global distribution map of lake chlorophyll. Int. J. Remote Sens. 36, 1942–1964 (2015).
Gasper, R. R., Selman, M. & Ruth, M. Climate co-benefits of water quality trading in the Chesapeake Bay watershed. Water Policy 14, 758–765 (2012).
Le Quere, C. et al. Global carbon budget 2016. Earth Syst. Sci. Data 8, 605–649 (2016).
Sepulveda-Jauregui, A. et al. Eutrophication exacerbates the impact of climate warming on lake methane emission. Sci. Total Environ. 636, 411–419 (2018).
Davidson, T. A. et al. Synergy between nutrients and warming enhances methane ebullition from experimental lakes. Nat. Clim. Change 8, 156–160 (2018).
Smith, V. H. Eutrophication of freshwater and coastal marine ecosystems: a global problem. Environ. Sci. Pollut. Res. Int. 10, 126–139 (2003).
Bouwman, L. et al. Exploring global changes in nitrogen and phosphorus cycles in agriculture induced by livestock production over the 1900–2050 period. Proc. Natl. Acad. Sci. USA 110, 20882–20887 (2013).
Tilman, D. et al. Forecasting agriculturally driven global environmental change. Science 292, 281–284 (2001).
Acknowledgements
This work was supported by the Minnesota Sea Grant College Program and by the Industrial Research Chair in Carbon Biogeochemistry in Boreal Aquatic Systems (CarBBAS), co-funded by the Natural Sciences and Engineering Research Council of Canada (NSERC) and Hydro-Québec. The joint lake-size by chla distribution, CH4 emission rates, and corresponding chla values are available at the figshare data repository (https://figshare.com/s/0a39281088d644a1d925). This document has been reviewed in accordance with the U.S. Environmental Protection Agency policy and approved for publication. The views expressed in this paper are those of the authors and do not necessarily reflect the views or policies of the U.S. Environmental Protection Agency.
Author information
Authors and Affiliations
Contributions
T.D., J.J.B., and J.A.D. identified the research questions, designed the study approach, and wrote the manuscript. J.A.D. identified the upscaling approach and compiled data on future nutrient loading to lakes and impoundments. J.J.B. wrote the code to implement the scenarios.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Journal peer review information: Nature Communications thanks Sarian Kosten and the other anonymous reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Beaulieu, J.J., DelSontro, T. & Downing, J.A. Eutrophication will increase methane emissions from lakes and impoundments during the 21st century. Nat Commun 10, 1375 (2019). https://doi.org/10.1038/s41467-019-09100-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-019-09100-5
This article is cited by
-
The EU Communication on ensuring availability and affordability of fertilisers—a milestone for sustainable nutrient management or a missed opportunity?
Environmental Sciences Europe (2024)
-
Salinity causes widespread restriction of methane emissions from small inland waters
Nature Communications (2024)
-
Nitrous oxide respiration in acidophilic methanotrophs
Nature Communications (2024)
-
Increased methane emission associated with anthropogenic activities in a highly urbanized tropical reservoir
International Journal of Environmental Science and Technology (2024)
-
Integrating Reservoirs into the Dissolved Organic Matter Versus Primary Production Paradigm: How Does Chlorophyll-a Change Across Dissolved Organic Carbon Concentrations in Reservoirs?
Ecosystems (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
