
Abstract
Changes in the diversity of plant communities may undermine the economically and environmentally important consumer species they support. The structure of trophic interactions determines the sensitivity of food webs to perturbations, but rigorous assessments of plant diversity effects on network topology are lacking. Here, we use highly resolved networks from a grassland biodiversity experiment to test how plant diversity affects the prevalence of different food web motifs, the smaller recurrent sub-networks that form the building blocks of complex networks. We find that the representation of tri-trophic chain, apparent competition and exploitative competition motifs increases with plant species richness, while the representation of omnivory motifs decreases. Moreover, plant species richness is associated with altered patterns of local interactions among arthropod consumers in which plants are not directly involved. These findings reveal novel structuring forces that plant diversity exerts on food webs with potential implications for the persistence and functioning of multitrophic communities.
Similar content being viewed by others


Introduction
Global change drivers are causing pervasive shifts in the distribution of biodiversity1. Concern that changes in biodiversity will threaten the services that nature provides to humans has sparked numerous experimental manipulations of diversity over the past decades2. These experiments typically manipulate plant communities, which provide food and habitat for myriad consumer species that perform important ecosystem services such as pest control and pollination. Reductions in plant diversity are associated with reduced consumer species richness3,4 and increased temporal variability in consumer community abundance5,6. These effects have been attributed to a propagation of plant diversity effects through multiple trophic levels3,4,7. However, a lack of well-resolved food webs has hindered mechanistic insight into plant diversity effects on the stability and robustness of multitrophic communities. Consequently, we are only just beginning to understand the effects of plant diversity on the structure of trophic interactions7,8,9.
The number and arrangement of feeding links in food webs determine the ability of the network to endure perturbations that result in species loss10,11. One way to fingerprint the structure of food webs is to examine the distribution of the small recurring subgraphs that are the building blocks of larger networks, termed network modules or motifs12,13. When excluding cannibalism, there are 13 directed, connected configurations of three-node motifs, that is, triads (Fig. 1). Examination of the distribution of motifs in a range of biological networks, including empirical food webs, has revealed that four of these motifs account for the vast majority of the triads:14,15,16 tri-trophic chains (s1), omnivory (s2), apparent competition (s4), and exploitative competition (s5). This non-random representation may arise due to two non-exclusive processes14,15,16,17,18. First, there may be constrains on how the network assembles such that some motifs become more common. Second, some motifs may function in a way that makes them beneficial to network stability, increasing their abundance in webs that have persisted so that they can be observed. However, current insight into patterns of network motif distributions in food webs comes predominantly from comparative studies using networks collected with different methods in a diverse range of ecosystems14,19.

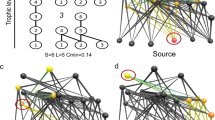
Three-species motifs in food webs. a The 13 possible connected triads, which may contain only single feeding links (labelled with ‘s’) or at least one double feeding link (‘d’). The common motifs are shown in colour: tri-trophic chains (e.g. a plant fed on by a herbivore which is then preyed upon by a predator; s1), omnivory (the species at the top of the food chain feeds on both other species; s2), apparent competition (two resources that are fed on by the same consumer; s4) and exploitative competition (a resource shared by two consumers; s5). b A hypothetical example of the consumer community (grey nodes) observed on a plot with two plant species (white nodes) and a node for detritus (black node). Coloured links show examples of the common triads that reoccur within the larger network. In this case, the omnivory (s2; blue) and exploitative competition (s5; pink) motifs are grounded (i.e. connected to a basal resource), while the tri-trophic chain (s1; green) and apparent competition (s4; orange) motifs are free floating (i.e. not connected to a basal resource)
Here, we examined the effect of plant species richness on the distribution of three-species motifs in replicate aboveground food webs from a long-running grassland biodiversity experiment. We constructed food webs for 80 grassland plots that were sown with a plant diversity gradient ranging from 1 to 60 plant species20. Feeding links were assigned using literature- and trait-based rules to populate an interaction metaweb, which was combined with encounter probability based on species abundances on each plot21,22. Subsequently, we enumerated all three-species motifs in each food web and assessed their representation relative to counts in randomly rewired null-model networks to account for the relationships among plant species richness, network size, and connectance. Further, we sought to elucidate whether any effect of plant species richness on motif distribution was restricted to motifs that are grounded, that is, they contain a basal resource node (plant or other resource such as detritus)17, or whether plant species richness remotely affects the local structure of interactions among consumers at higher trophic levels. Accordingly, we examined changes in the frequency of free-floating motifs (i.e. not connected to a basal resource) in a sub-web for each plot containing only the arthropod consumers.
The positive effect of plant diversity on consumer species richness is strongest for consumers at low trophic levels and with a low degree of omnivory3,4,23. Herbivore species richness increases more rapidly with plant diversity than predator or omnivore species richness3,4,23. Thus, the herbivore:predator richness ratio increases with plant diversity4. In line with these patterns, we hypothesised that the representation of apparent and exploitative competition motifs (s4 and s5) would increase with plant species richness, with a corresponding decrease in the representation of tri-trophic chain and omnivory motifs (s1 and s2). This is because the competition motifs (s4 and s5) represent a feeding interaction that can occur between only two trophic levels. Consequently, these motifs may become relatively more common in food webs with more plant and herbivore nodes than motifs requiring an omnivore or predator (s1 and s2). Our results confirmed these hypotheses with the exception of trends in the representation of the tri-trophic chain motif (s1), providing novel insights into the assembly of food web substructures across a gradient of diversity. We contend that the observed changes in motif representation with increasing plant species richness may be due to selection against local structures that are unstable according to ecological theory.
Results
Effect of plant diversity on motif counts in full networks
The total number of all motifs in the food webs increased with plant species richness (Fig. 2), as expected due to the increase in network size. The four common configurations (tri-trophic chains, omnivory, apparent competition, and exploitative competition) accounted for the vast majority of the total number of motifs regardless of plant species richness (mean >99%). Apparent competition motifs were particularly numerous, especially at high plant species richness (mean 41–58% of all motifs across richness levels; Fig. 2c). Three-species structures with a loop (motif s3) or double links (motifs labelled with ‘d’) were uncommon (mean <1%), with the majority of food webs containing none (Fig. 2e). At high plant species richness, tri-trophic chains and both apparent and exploitative competition motifs became more highly represented in the observed networks than in rewired null models (Fig. 3a, c, d; Supplementary Table 1). In contrast, the representation of omnivory motifs significantly decreased across the gradient of plant species richness (Fig. 3b; Supplementary Table 1).

Frequency of motifs in food webs. Effect of sown plant species richness on mean counts of tri-trophic chain (a, s1), omnivory (b, s2), apparent competition (c, s4), exploitative competition (d, s5) and other motifs (e, i.e. sum of s3 and all double motif counts). Filled points and solid lines show motif frequency in the full food web, and open points and dashed lines display frequency in the consumer sub-web (i.e. free-floating motifs). P values from linear mixed models (two-tailed test) for the slope of all displayed relationships are <0.001 (for full model results see Supplementary Table 1). Linear models for e (other) could not be validated due to the bimodal distribution of the motif frequency

Representation of motifs relative to null models. Effect of sown plant species richness on mean normalised z-scores for tri-trophic chain (a, s1), omnivory (b, s2), apparent competition (c, s4) and exploitative competition (d, s5). Filled points and solid lines show motif frequency in the full food web, and open points and dashed lines display frequency in the consumer sub-web (i.e. free-floating motifs). P values from linear mixed models (two-tailed test) for the slope of all displayed relationships are <0.001 (for full model results see Supplementary Table 1)
Effect of plant diversity on motifs in consumer sub-webs
For all common configurations, the contribution of grounded motifs to the total number of motifs increased with plant species richness. This was evidenced by the number of free-floating motifs not increasing with plant species richness as quickly as the total number of motifs (Fig. 2; Supplementary Table 1). However, compared to null models, the effects of plant species richness on the representation of the common free-floating motifs were similar to the patterns observed for the full food webs (Fig. 3; Supplementary Table 1).
Discussion
Here, we performed a novel empirical test of how a gradient of plant diversity alters complex food web structure. Specifically, plant diversity shifted the distribution of three-species motifs. In food webs at every plant diversity level, tri-trophic chains (s1) were generally overrepresented compared to the null models (Fig. 3a). This result mirrors an analysis of 50 empirical food webs that used the same null model we use here14. However, the significantly increasing overrepresentation of tri-trophic chains with plant diversity contrasted our initial hypothesis that relatively weak effects of plant species richness on predator species richness would be associated with reductions in the representation of tri-trophic chain motifs. This was perhaps due to a shift in the omnivore community towards predatory species24, or because predators could form many partially overlapping tri-trophic chains by preying on a generalist herbivore, detritivore or omnivore that was involved in numerous apparent competition interactions. We observed a similar increasing trend for both the competition motifs (s4 and s5). On average, these motifs were underrepresented at low plant diversity and overrepresented at high plant diversity (Fig. 3c, d), the latter being typical of many empirical food webs14. This trend matched our expectation based on the increasing contribution of plants and herbivores to total species richness compared to predators in high-diversity communities, which greatly increases the possibilities for interactions between the two lower trophic levels. However, plant diversity effects on motif representation were conserved even when considering only the consumer sub-web, indicating that plant diversity influences the local structure of food webs beyond the immediate interaction partners of the plants.
Omnivory motifs (s2) were overrepresented compared to null models in most cases, but in contrast to the trends for tri-trophic chains (s1) and competition motifs (s4 and s5), this overrepresentation significantly decreased with plant species richness (Fig. 3b). This confirms our initial hypotheses, congruent with a proportional decrease in the number of omnivores present in the networks from high diversity communities (Supplementary Fig. 1). Among other empirical food webs, the omnivory motif shows the most highly variable patterns of over- and under-representation14,18,19. A potential reason for this inconsistency is that the role of omnivory for food web stability is highly dependent on interaction strengths and network context15,25,26. Early theoretical work suggested that omnivory would destabilise complex networks, but a range of theoretical and empirical work has more recently demonstrated the conditions under which omnivory may be stabilising, such as when embedded in larger networks or when omnivores feed adaptively25,27.
Comparative studies have shown that mixed patterns of motif representation among empirical food webs is not limited to omnivory motifs19. However, previous work has rarely considered the species composition of motifs28, which could influence their dynamic behaviour. For instance, plants cannot engage in behaviours such as seeking refugia that can influence motif persistence29. Our experimental system revealed that plant diversity altered the trophic identity of species constituting motifs. For example, when plant nodes are few at low plant diversity, most s2 motifs were free floating (mean 88% in the monocultures; Fig. 2b). This actually represents a case of intraguild predation among three consumers, where a top predator feeds on an intermediate prey with which it also competes with for a lower prey30. As plant diversity increases, there is an increase in the proportion of s2 motifs containing a basal resource, that is, classic omnivory of a consumer feeding from two trophic levels (mean 65% in 60-species communities). This is important because there are only limited conditions under which the intermediate consumer can coexist with the two-fold challenge of predation and competition in intra-guild predation31. In contrast, plants are generally not killed or consumed as a whole by their arthropod consumers. Consequently, future investigations will benefit from considering the traits of species in food web motifs in addition to motif prevalence.
Debate currently surrounds the role and relative importance of the different three-species motifs for food web stability. Multiple recent studies have assessed the stability of three-species motifs by generating replicate Jacobian matrices based on draws of interaction strength, and assessing whether the resulting motif persists (i.e. returns to equilibrium) following a small perturbation at steady state (i.e. local stability)14,15. This exercise revealed that in isolation (i.e. when not embedded in larger networks) tri-trophic chains and competition motifs (s1, s4 and s5) are the most locally stable, while s3 (feed-forward loop) and all motifs with double links were unstable. Omnivory (s2) was unique in that it was moderately stable in isolation, and whether a return to equilibrium occurred depended on the sampled interaction strengths. Further, there is a correlation between the local stability of motifs and their frequency in empirical food webs; motifs s1, s4 and s5 occur more frequently than s2. Consequently, the authors suggest that these motifs are selected for because their stability offers some advantage to network function or persistence14,15. Based on these findings, we could make the intuitive conclusion that food webs assembling on plots with more diverse plant communities may be more robust to disturbances, as they contain relatively higher s1, s4 and s5 motif representation, and relatively lower s2 representation than the low plant diversity communities. However, the assumptions of these assessments may be violated in empirical systems because the motifs do not exist in isolation and exhibit seasonal population fluctuations rather than being at steady state.
Other theoretical investigations suggest that the stability of common motifs in isolation is not directly related to their influence on whole network persistence when they are embedded in complex food webs17. This conclusion was reached by assessing the initial number of s1, s2, s4 and s5 motifs in food webs generated with a common food web model (the niche model32) and tracking how the initial frequencies affected the long-term persistence of the entire networks with dynamic simulations. In this case, omnivory (s2) motifs contributed positively to whole network persistence, while high frequencies of both competition motifs (s4 and s5) decreased the likelihood of whole network persistence. The surprising implication of this result is, that based on network substructure, we would imply that the food webs from communities with high plant diversity are less likely to persist in the long term. We do not test any metric of network stability, so we cannot shed light on the link between our empirical results and the current state of the network stability theory. However, such a condition conflicts the fact that natural diverse communities can and do persist. Reconciliation of this potential mismatch between simulations and empirical findings will benefit from studies of food web assembly. In particular, we note that better understanding the role of dispersal, a feature not considered in the closed systems of previous modelling work, could be important. It may be that a high diversity of resources can support unstable interaction structures by being more attractive to colonists from the regional species pool. In a similar vein, omnivory motifs may be selected for at low plant diversity to enable network persistence despite these communities being less attractive to arthropod colonists.
There are two potential caveats to using a metaweb of plausible interactions to assess variation in network structure across a gradient of plant diversity. First, we do not know whether the interactions identified by the literature and trait-based rules actually occurred at the plot level. However, we determined that our results are robust to the assumptions we made during food web construction by varying the likelihood that plausible consumer–plant and predator–prey interactions actually occur according to their relative abundances (Supplementary Note 1, Supplementary Figs 3-6). Second, the metaweb approach implies that interactions between two species are an intrinsic property of the species based on their traits, and therefore that consumer feeding behaviour does not vary with plant species richness. There is evidence that this assumption may be violated; prey suppression may be lowered in more complex habitats, potentially due to an increase in prey refugia33. We showed that our results are not sensitive to this potential scenario with an additional analysis that lowered the density-dependent probability of predator–prey interactions with increasing plant diversity (Supplementary Note 2, Supplementary Fig. 7). While these measures cannot replace actual observations of every interaction, constructing species-level food webs with other methods (e.g. gut content or stable isotope analysis) across the large number of replicates required to appropriately test our hypotheses is highly infeasible. By combining the best available evidence on observed feeding interactions on the study site8 with established methods for determining interactions from literature and trait-based evidence34,35,36, we offer a novel and robust insight into plant diversity effects on food web structure.
We have shown the first empirical evidence that changes in plant diversity induce shifts in the representation of three-species motifs in food webs. Further, we demonstrate that plant diversity affects the local interaction structure of consumers at higher trophic levels, even though plants are not directly involved in those interactions. This supports suggestions that the bottom-up effects of plant communities on multitrophic community abundance and richness are mediated through trophic interactions3. Additionally, the effects on network structure do not simply reflect the effect of plants on consumer species diversity; in some cases, the trends in network substructure could not be predicted from our current understanding of plant diversity effects on the richness of higher trophic levels. Specifically, tri-trophic chains become more over-represented at high plant diversity even though plant diversity effects on consumers attenuate with increasing trophic level3,4. Given the need for a continued provision of ecosystem services in the face of global changes, a central priority is to increase understanding of how these diversity-induced changes in network structure are related to the stability and functioning of multitrophic communities37,38. Further testing of theoretical predictions of network structure with empirical evidence across experimental gradients will be crucial in this context.
Methods
Experimental design and plant surveys
Data were collected on 80 grassland plots (each 6 × 9 m2) of The Jena Experiment (Germany; 50°55′N, 11°35′E, 130m a.s.l.) that were sown with 1, 2, 4, 8, 16 or 60 grass, small herbs, tall herbs and legume species in 2002 on a previously arable field20. The plant species mixtures were maintained by weeding three times per year and mowing twice annually39. The plots are arranged in four blocks corresponding to variation in soil conditions. The presence (realised richness) and percentage cover of the initially sown plant species remaining on the plots in 2010 and 2012 was assessed visually on a 3 × 3 m2 area of each plot twice per year (May and August). Additional static basal resources, comprising detritus, moss, algae, fungi, dung and carrion, were assumed to be present on each plot based on previous observations.
Arthropod sampling and traits
Sampling of aboveground invertebrates was conducted over two sampling periods in each of 2010 and 2012 (May/June and July/August). Ground-dwelling taxa were sampled with two pitfall traps (each 4.5 cm diameter) that were open for 20 to 29 days in each time period, and herb-layer taxa were sampled by two suction samples of an area 0.75 × 0.75 m2 with a modified industrial vacuum cleaner24. Taxa included in food webs are those with more than two individuals sampled on the entire field site in a given year, and were identified to species level (a total of 403 consumer species). This included taxa from the classes Arachnida (orders Araneae and Opiliones), Chilopoda (orders Geophilomorpha and Lithobiomorpha), Diplopoda (orders Julida and Polydesmida), Insecta (orders Coleoptera, Hemiptera, Orthoptera and Thysanoptera) and Malacostraca (order Isopoda). Achieving a high resolution of taxonomic identification across all plots and time periods precluded the inclusion of every consumer group (e.g. Diptera, Hymenoptera and vertebrates were omitted), so that our replicate networks represent a consistent subset of the aboveground food web. Due to the data-intensive nature of food web construction, most existing food webs do not include every species (e.g. parasites40). Traits for each consumer species were assembled through extensive literature searches24. These included body length, coarse feeding guild (detritivore, herbivore, omnivore or predator) and the vertical habitat stratum primarily used by the consumer (ground layer, herb layer or both).
Food web construction
We constructed food webs for each plot in each time period (n = 4; May/June and July/August in 2010 and 2012) by first identifying the co-occurring species from plant inventories, pitfall traps and suction samples along with the static resource nodes. However, co-occurrence does not necessarily mean that species are interacting41. Consequently, we assigned trophic interactions based on the mathematical framework for population-level interactions of Poisot et al.22. Under this framework, the probability that species i and j interact in the adjacency matrix A is
where N is a function defining the encounter probability of two species based on their local abundances, and T is a function giving the probability that two species actually interact based on their traits. The term ε describes higher-order effects such as the impact of the environment on interaction probability22.
We defined whether two species will interact (i.e. T) based on evidence from extensive literature searches and trait-based rules, which were used to construct a metaweb of all plausible trophic interactions21. The feeding links in the metaweb were categorised into five link types depending on how they were assigned: (1) specific feeding interactions reported in the literature; (2) generalised feeding interaction reported in the literature; (3) links based on trophic levels; (4) trait-based rules; and (5) combined trait-based rules. Generally, link types 1–3 were applied to interactions between basal resources (i.e. plants or static resource nodes) and herbivore, detritivore and omnivore consumers. An example of a generalised feeding interaction reported in the literature (link type 2) is for feeding by the beetle Chrysolina oricalcia, which is oligophagous and commonly feeds on plants in the family Apiaceae42. Link types 4 and 5 (those assigned from trait-based information) generally describe predatory (i.e. consumer–consumer) interactions. For example, the large web-building spider Coelotes terrestris preys on non-spider prey that forage on the ground43,44. For full details of metaweb construction see ref. 21. For each plot, the relevant rows and columns for co-occurring species were extracted, and when an interaction was identified in the metaweb, the interaction probability Tij was 1 because there is strong evidence these species will interact if they encounter each other21.
Encounter probability N depended on the link type in the metaweb and the trophic guild of the resource (i), so that interaction probability Aij is
where n is relative cover or abundance and α or β are scalar values. We assumed that species would always encounter one another (i.e. N = 1) in the case of species interactions that have been specifically identified in the literature (link type 1; Eq. 2). This is because these links are applied most often to highly specialised feeding interactions (e.g. monophagous herbivores) and the species are likely co-occurring because one has sought out the other as a food resource. The same logic was applied to interactions identified from generalised literature reports or based on trophic guild rules (link types 2 and 3) where the resource was a consumer (Eq. 3). Encounter probability was also certain for links from generalised literature reports where the resource was a static node such as detritus that is ubiquitous (Eq. 3). In all other cases, encounter probability was defined as proportional to the product of the relative cover (for plants) or relative abundance (for consumers, calculated separately for ground- and herb-layer)45. We considered two scalar values (α or β) because it is likely that encounter probability operates differently for plant–consumer interactions (link types 2 and 3; Eq. 4) and predator–prey interactions (types 4 and 5; Eq. 5).
Determining appropriate values for α and β is important because these values define the likelihood that generalist herbivores and omnivores encounter appropriate plants (α; Eq. 4) and predators encounter prey (β ; Eq. 5). Increasing α or β therefore has the effect of increasing the feeding generalism of many populations in the food webs. We considered a range of values for α and β and assigned the values based on available evidence from DNA-based analysis of gut contents for an omnivorous beetle sampled from another plant-diversity experiment located on the same field site8. This study identified an increase in the total number of feeding interactions (plants and animals) from ca. 2 to ca. 7 along a plant diversity gradient of 1 to 8 species8. Consequently, we selected values of α and β that maximised the median generality of omnivores in each food web at ca. 30 species (Supplementary Fig. 2). This corresponds to a mean realised plant species richness of ca. 33 species by 2010 and 2012 on plots that were initially sown with 60 species in 2002. Although this is an extrapolation, the choice of α and β do not qualitatively affect the conclusions of our study (see Supplementary Note 1 and Supplementary Figs. 3–6). Note that our approach does not consider how higher-order interactions or environmental context (i.e. ε in Eq. 1) may affect the probability of species interactions differently across the plant diversity gradient. Consequently, we performed an analysis to show that our results were not sensitive to the feasible scenario where encounter probability is lower at high plant diversity (Supplementary Note 2 and Supplementary Fig. 7) due to an increase in habitat complexity that provides refugia for prey33.
Analysis of motifs in food webs
After obtaining the matrix of interaction probabilities (Aij) for each plot and time period, we sampled these probabilities 50 times and calculated network properties for the resulting binary food web at each iteration. Any nodes that were disconnected from a network (i.e. degree = 0) after sampling the probabilities were omitted from the calculation of network properties. The resulting plot-level trophic networks contained a mean of 20.4 to 99.0 species and mean of 36.2 to 1060.0 trophic links. The frequency of each of the 13 three-species motifs in all networks was enumerated using the triad.census function from the igraph package46 in R47 as implemented in custom functions by reference14. Each set of three species was assigned to only one motif13,14,19, the one with the highest link density. For example, three species interacting in an omnivory arrangement did not also interact as the tri-trophic chain, apparent competition and exploitative competition motifs contained within. All reported motif frequencies are the mean number from the 50 iterations of each food web. To count the number of free-floating motifs (those not connected to a basal resource17), we omitted plant and static resource nodes from the network (leaving a consumer sub-web) and repeated this process. The motif images in the figures were produced using previously published code14 and colours48.
Randomisation of networks and null-web benchmarking
Network properties such as size and connectance covary with plant species richness. Therefore, we compared the number of motifs in the plot-level food webs with the number in a null model web produced by randomly rewiring the feeding links. This was achieved using the Curveball algorithm49, which takes existing links, for example, A→B and C→D, and rewires them, resulting in A→D and C→B. This is performed in an unbiased way for the entire network, maintaining the number of species, links (and thus connectivity) and degree in and out of each node (i.e. row and column sums). Thus, this process creates a null expectation by redistributing links without modifying the fundamental ecological properties of the network (e.g. plants do not eat consumers), and has previously been used in studies of food web motifs14. This null model allows us to gain insight into how the food web assembles non-randomly across the gradient of plant species richness.
For each iteration of the probability-based food webs, we generated 250 rewired networks and calculated the representation of each of the 13 motifs i assessed by calculating a z-score:13
where Xi is the frequency of motif i in the sampled web, and \(\overline {X_i}\) and σi are the mean and standard deviation of the frequency of motif i in the rewired null-model networks. The z-score vector was subsequently normalised for each food web because larger networks show more extreme patterns of over- or under-representation:50
where j is an index over the 13 motifs. The normalised value provides information on the relative importance rather than the absolute importance of a motif to a network, which allows comparison of networks of different sizes. A positive value within the normalised profile vector (ni) indicates that the motif i was overrepresented in the observed trophic network. Z-scores are reported as the mean from the 50 iterations of each matrix of interaction probabilities.
Statistical analyses
All statistical analyses were conducted in R47. We analysed the effect of sown plant species richness (two-tailed test) on motif counts and normalised z-scores with linear mixed models using the lmer function of the lme4 package51. Random effects were time period (n = 4) and plot (n = 80) within experimental block (n = 4) (total n for all analyses = 320). Motif frequencies were log transformed for normality. Lines of fit in Figs. 2, 3 were predicted from the fixed model parameters reported in Supplementary Table 1.
Reporting summary
Further information on experimental design is available in the Nature Research Reporting Summary linked to this article.
Code availability
Custom R scripts used to generate and analyse the data are available on GitHub (https://doi.org/10.6084/m9.figshare.7605173.v2).
Data availability
The datasets generated and analysed during the current study (including raw data for Figs. 2, 3 and Supplementary Figs.) are available in the figshare repository (https://doi.org/10.6084/m9.figshare.7605242.v2). The metaweb of feeding interactions that supports the findings of this study is available in ref. 21.
References
Butchart, S. H. M. et al. Global biodiversity: indicators of recent declines. Science 328, 1164–1168 (2010).
Cardinale, B. J. et al. Biodiversity loss and its impact on humanity. Nature 486, 59–67 (2012).
Scherber, C. et al. Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature 468, 553–556 (2010).
Haddad, N. M. et al. Plant species loss decreases arthropod diversity and shifts trophic structure. Ecol. Lett. 12, 1029–1039 (2009).
Proulx, R. et al. Diversity promotes temporal stability across levels of ecosystem organization in experimental grasslands. PLoS ONE 5, e13382 (2010).
Haddad, N. M., Crutsinger, G. M., Gross, K., Haarstad, J. & Tilman, D. Plant diversity and the stability of foodwebs. Ecol. Lett. 14, 42–46 (2011).
Rzanny, M. & Voigt, W. Complexity of multitrophic interactions in a grassland ecosystem depends on plant species diversity. J. Anim. Ecol. 81, 614–627 (2012).
Tiede, J. et al. Trophic and non-trophic interactions in a biodiversity experiment assessed by next-generation sequencing. PLoS ONE 11, e0148781 (2016).
Welti, E., Helzer, C. & Joern, A. Impacts of plant diversity on arthropod communities and plant–herbivore network architecture. Ecosphere 8, e01983 (2017).
McCann, K. S. The diversity–stability debate. Nature 405, 228–233 (2000).
Dunne, J. A., Williams, R. J. & Martinez, N. D. Network structure and biodiversity loss in food webs: robustness increases with connectance. Ecol. Lett. 5, 558–567 (2002).
Bascompte, J. & Melián, C. J. Simple trophic modules for complex food webs. Ecology 86, 2868–2873 (2005).
Milo, R. et al. Network motifs: simple building blocks of complex networks. Science 298, 824–827 (2002).
Borrelli, J. J. Selection against instability: stable subgraphs are most frequent in empirical food webs. Oikos 124, 1583–1588 (2015).
Monteiro, A. B. & Faria, L. D. B. The interplay between population stability and food-web topology predicts the occurrence of motifs in complex food-webs. J. Theor. Biol. 409, 165–171 (2016).
Prill, R. J., Iglesias, P. A. & Levchenko, A. Dynamic properties of network motifs contribute to biological network organization. PLoS Biol. 3, e343 (2005).
Stouffer, D. B. & Bascompte, J. Understanding food-web persistence from local to global scales. Ecol. Lett. 13, 154–161 (2010).
Camacho, J., Stouffer, D. B. & Amaral, L. A. N. Quantitative analysis of the local structure of food webs. J. Theor. Biol. 246, 260–268 (2007).
Stouffer, D. B., Camacho, J., Jiang, W. & Nunes Amaral, L. A. Evidence for the existence of a robust pattern of prey selection in food webs. Proc. R. Soc. Ser. B 274, 1931–1940 (2007).
Roscher, C. et al. The role of biodiversity for element cycling and trophic interactions: an experimental approach in a grassland community. Basic Appl. Ecol. 5, 107–121 (2004).
Hines, J. et al. A meta-food web for invertebrate species collected in an experimental grassland. Ecology (2018). https://doi.org/10.1002/ecy.2679.
Poisot, T., Stouffer, D. B. & Gravel, D. Beyond species: why ecological interaction networks vary through space and time. Oikos 124, 243–251 (2015).
Ebeling, A. et al. Plant diversity effects on arthropods and arthropod-dependent ecosystem functions in a biodiversity experiment. Basic Appl. Ecol. 26, 50–63 (2018).
Ebeling, A. et al. Plant diversity induces shifts in the functional structure and diversity across trophic levels. Oikos 127, 208–219 (2018).
Kratina, P., LeCraw, R. M., Ingram, T. & Anholt, B. R. Stability and persistence of food webs with omnivory: Is there a general pattern? Ecosphere 3, 1–18 (2012).
Wootton, K. L. Omnivory and stability in freshwater habitats: does theory match reality? Freshwat. Biol. 62, 821–832 (2017).
Fagan, W. F. Omnivory as a stabilizing feature of natural communities. Am. Nat. 150, 554–567 (1997).
Cirtwill, A. R. & Eklöf, A. Feeding environment and other traits shape species’ roles in marine food webs. Ecol. Lett. 21, 875–884 (2018).
HilleRisLambers, R., Van De Koppel, J. & Herman, P. M. J. Persistence despite omnivory: benthic communities and the discrepancy between theory and observation. Oikos 113, 23–32 (2006).
Polis, G. A., Myers, C. A. & Holt, R. D. The ecology and evolution of intraguild predation: potential competitors that eat each other. Annu. Rev. Ecol. Syst. 20, 297–330 (1989).
Diehl, S. & Feissel, M. Intraguild prey suffer from enrichment of their resources: microcosm experiment with ciliates. Ecology 82, 2977–2983 (2001).
Williams, R. J. & Martinez, N. D. Simple rules yield complex food webs. Nature 404, 180–183 (2000).
Finke, D. L. & Denno, R. F. Intraguild predation diminished in complex‐structured vegetation: implications for prey suppression. Ecology 83, 643–652 (2002).
Morales-Castilla, I., Matias, M. G., Gravel, D. & Araújo, M. B. Inferring biotic interactions from proxies. Trends Ecol. Evol. 30, 347–356 (2015).
Mendonça, V. et al. What's in a tide pool? Just as much food web network complexity as in large open ecosystems. PLoS ONE 13, e0200066 (2018).
Pomeranz, J. P. F., Thompson, R. M., Poisot, T. & Harding, J. S. Inferring predator–prey interactions in food webs. Methods Ecol. Evol. (2018). https://doi.org/10.1111/2041-210X.13125.
Hines, J. et al. Towards an integration of biodiversity-ecosystem functioning and food web theory to evaluate relationships between multiple ecosystem services. Adv. Ecol. Res. 53, 161–199 (2015).
Thompson, R. M. et al. Food webs: reconciling the structure and function of biodiversity. Trends Ecol. Evol. 27, 689–697 (2012).
Weisser, W. W. et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: patterns, mechanisms, and open questions. Basic Appl. Ecol. 23, 1–73 (2017).
Lafferty, K. D. et al. Parasites in food webs: the ultimate missing links. Ecol. Lett. 11, 533–546 (2008).
Barner, A. K., Coblentz, K. E., Hacker, S. D. & Menge, B. A. Fundamental contradictions among observational and experimental estimates of non-trophic species interactions. Ecology 99, 557–566 (2018).
Cox, M. L. Atlas of the Seed and Leaf-Beetles of Britain and Ireland (Information Press, Oxford, 2007).
Nyffeler, M. & Benz, G. Foraging ecology and predatory importance of a guild of orb-weaving spiders in a grassland habitat. J. Appl. Entomol. 107, 166–184 (1989).
Nyffeler, M., Sterling, W. L. & Dean, D. A. How spiders make a living. Environ. Entomol. 23, 1357–1367 (1994).
Canard, E. F. et al. Empirical evaluation of neutral interactions in host–parasite networks. Am. Nat. 183, 468–479 (2014).
Csardi, G. & Nepusz, T. The igraph software package for complex network research. InterJ. Complex Syst. 1695, 1–9 (2006).
R Development Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, 2017). http://www.R-project.org.
Wong, B. Points of view: color blindness. Nat. Methods 8, 441 (2011).
Strona, G., Nappo, D., Boccacci, F., Fattorini, S. & San-Miguel-Ayanz, J. A fast and unbiased procedure to randomize ecological binary matrices with fixed row and column totals. Nat. Commun. 5, 4114 (2014).
Milo, R. et al. Superfamilies of evolved and designed networks. Science 303, 1538–1542 (2004).
Bates, D., Maechler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 (2015).
Acknowledgements
This study was funded by the German Research Foundation (DFG; FOR 1451). We acknowledge support for the Jena Experiment from Friedrich Schiller University (FSU) Jena and the Max Planck Institute for Biogeochemistry. We are grateful to The Jena Experiment scientific coordination team, the gardening staff for the maintenance of the field site and the many student helpers that weeded the experimental plots. We acknowledge Jessy Loranger and Christian Ristok for collecting data used in this analysis, Ulrich Brose and Andrew Barnes for advice and Simone Cesarz and Manfred Türke for creating the images in Fig. 1b. Further support came from the German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, funded by the German Research Foundation (FZT 118).
Author information
Authors and Affiliations
Contributions
J.H. and A.E. conceived the project; N.E., S.T.M., C.R., M.R,. W.V., W.W.W. contributed data; J.H., A.E. and D.P.G. compiled the data; D.P.G. performed the analyses and wrote the manuscript; all authors discussed the results and contributed to the manuscript text.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Journal peer review information: Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Giling, D.P., Ebeling, A., Eisenhauer, N. et al. Plant diversity alters the representation of motifs in food webs. Nat Commun 10, 1226 (2019). https://doi.org/10.1038/s41467-019-08856-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-019-08856-0
This article is cited by
-
Imprints of indirect interactions on a resource-mediated ant–plant network across different levels of network organization
Oecologia (2024)
-
Fear or food? Prey availability is more important than predation risk in determining aerial insectivorous bat responses across a disturbed tropical forest landscape
Biodiversity and Conservation (2023)
-
Vector-borne plant pathogens modify top-down and bottom-up effects on insect herbivores
Oecologia (2021)
-
Biodiversity increases multitrophic energy use efficiency, flow and storage in grasslands
Nature Ecology & Evolution (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
