
Abstract
In the face of the biodiversity crisis, it is argued that we should prioritize species in order to capture high functional diversity (FD). Because species traits often reflect shared evolutionary history, many researchers have assumed that maximizing phylogenetic diversity (PD) should indirectly capture FD, a hypothesis that we name the “phylogenetic gambit”. Here, we empirically test this gambit using data on ecologically relevant traits from >15,000 vertebrate species. Specifically, we estimate a measure of surrogacy of PD for FD. We find that maximizing PD results in an average gain of 18% of FD relative to random choice. However, this average gain obscures the fact that in over one-third of the comparisons, maximum PD sets contain less FD than randomly chosen sets of species. These results suggest that, while maximizing PD protection can help to protect FD, it represents a risky conservation strategy.
Similar content being viewed by others


Introduction
We are in the midst of a period of heightened biological extinction, with rates several orders of magnitude higher than background rates estimated from the fossil record1,2,3. In addition to having potentially widespread consequences for the functioning of ecosystems and the provisioning of valuable ecosystem services, this situation poses an immense moral challenge4,5,6,7,8. Since the extent that resources for conservation actions remain limited, agonizing choices as to which species most warrant attention become necessary9,10. To keep humanity’s options open, and our common legacy as rich as possible, it is widely argued that we should seek to maximize the biological diversity of form and function in conservation strategies6,7,8,9,10,11,12. The biological diversity of form and function can be measured as functional diversity (FD) (see Methods). However, in practice, it is challenging to prioritize species on the basis of FD: we have imperfect knowledge about which, and how many traits and functions are important in a given context, how these traits and functions vary among species and across space, and how the importance of traits may change in the future13. Many researchers have therefore advocated for a hypothesis that we name the ‘phylogenetic gambit’; that is, if species traits reflect their shared evolutionary history, then the pattern of that evolutionary history—their phylogeny—should serve as a useful stand-in for unmeasured and unmeasurable traits9,14,15. The phylogenetic gambit implies that maximizing phylogenetic diversity (PD), i.e., the breadth of evolutionary history, will ensure that a wide variety of forms and functions are present within a species set14,15,16,17.
Following this logic, phylogenetic diversity has formed the basis of global conservation schemes, notably the EDGE of existence program18 has been used by restoration biologists19 and has been widely embraced by researchers across the biodiversity sciences20,21,22,23. Despite this enthusiasm, the critical question of whether maximizing PD will actually capture more FD than prioritization schemes that ignore phylogeny has, to our knowledge, never been empirically tested16. Some studies have discussed24,25 and documented the relationship between FD and PD, both at regional26 and global scales20,22, and many of these studies have shown that maximizing PD does not maximize FD. However, such studies do not test the fundamental phylogenetic gambit at the heart of all PD-based conservation strategies: maximizing PD captures more FD than randomly choosing species. No one would dispute that the best way to maximize FD is to prioritize FD, but phylogenetic diversity has emerged as prioritization tool because we rarely have sufficient trait data to calculate FD. Here we test whether PD-based conservation passes a much less stringent, but ultimately more fundamental, test: is conserving on the basis of PD better than conserving at random? Worryingly, a recent theoretical study has demonstrated that PD could be a poor surrogate for FD and, in some scenarios, prioritizing species on the basis of PD could actually capture less FD than if species were simply selected at random16.
This recent work points to the need for empirical tests of the phylogenetic gambit, i.e., whether—within a given species pool—sets of species selected to maximize PD actually contain more FD than sets of species selected without regard to evolutionary relatedness. We clarify what our goals are in testing the utility of PD to capture FD. First, we take as given that maximizing PD is not the overarching goal per se of PD-maximization schemes, but rather that a PD maximization strategy is valued for its ability to capture more FD compared to a strategy that ignores phylogeny. Second, it is important to note that we are selecting species sets to maximize PD or FD within a region. While this is a simplification, as conservation actions often aim to select sets of areas (e.g., in reserve design), the only global phylogenetically informed conservation initiative is species-centered18 (EDGE). Critically, the question we raise has been shown to be distinct from asking whether traits have phylogenetic signal (whether closely related species tend to share similar sets of traits), since PD can be a poor surrogate for FD even if traits exhibit phylogenetic signal16.
Here, we test the phylogenetic gambit using data on ecologically-relevant traits from >15,000 vertebrate species. We find that maximizing PD results in an average gain of 18% of FD relative to random choice. However, this average gain obscures the fact that in over one third of the comparisons, maximum PD sets contain less FD than randomly chosen sets of species. These results suggest that, while maximizing PD protection can help to protect FD, it represents a risky conservation strategy.
Results
Approach
We evaluate the PD–FD relationship for different species pools (taxonomic families and geographical assemblages, i.e., sets of species co-occurring at a given scale) using a large global dataset including trait, phylogenetic, and geographic range data for 4616 species of mammals, 9993 species of birds, and 1536 species of tropical fish. Specifically, we measure FD as functional richness (see Methods) and compute, for any given species pool, an estimate of surrogacy27,28 (SPD–FD, Fig. 1). SPD–FD represents the amount of FD sampled by the set of species chosen to maximize PD, relative to the FD sampled by optimal set of species selected to maximize FD directly, with both components controlled for the expected FD from a random species set of the same size. SPD–FD will be positive if the averaged PD-maximized set contains more FD than the averaged random set, and negative if not. SPD–FD will equal 100% if the PD-maximization strategy is optimal (i.e. to maximize FD). We integrate SPD–FD for each species pool across all deciles of species richness but because they are many sets of species that can maximize PD or that can be chosen at random, we computed SPD–FD based on the averaged FD over 1000 PD-maximized sets and 1000 random sets16.

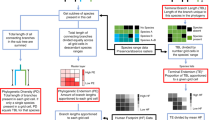
A conceptual approach for evaluating whether PD is a good surrogate for FD. To evaluate if PD is a good surrogate of FD, we measure to what extent a species prioritization strategy that maximize PD captures FD relative to an optimal and a random strategy. To do so, we compare FD accumulation curves (i.e., FD computed for increasing proportion of the species pool considered) across these three different sampling strategies: the random sampling (i.e., rarefaction curve, averaged over 1000 sets), the maxPD sampling, and the maxFD (optimal) sampling (i.e., sets that maximize FD, see legend). Then, we measure the surrogacy of PD for FD (SPD–FD) as the area between the random and the maxPD curve (“A”, see legend) divided by the area between the random and the maxFD curve (“A + B”, see legend). If SPD–FD is positive, PD is a good surrogate for FD (the maximum value being 1 where PD is an optimal surrogate) while when SPD–FD is negative preserving species based on PD is worse than preserving them at random
Mean surrogacy of PD for FD
We find that selecting the most phylogenetically diverse sets of species within a given taxonomic family or within a given geographical location (large grid-cells across the globe) captures, on average, 18% more FD than sets of randomly chosen species (i.e., SPD–FD = 18%, SD ± 6.5% across pools, see Figs. 2 and 3 and Supplementary Figures 1 and 2). Although the surrogacy is generally positive, there was substantial variation across species pools. For example, the surrogacy of PD varies widely from a minimum of −85% to a maximum of 92%, meaning that selecting the most phylogenetically diverse sets of taxa can capture either 85% less (or 92% more) FD than sets of randomly chosen taxa (Figs. 2 and 3 and Supplementary Figures 1 and 2). However, in 88% of the species pools, choosing sets of species according to PD captured more FD than would be expected at random (i.e., surrogacy values > 0 in 88% of the cases, see Figs. 2 and 3). This suggests that, on average, maximizing PD is a sound strategy to capture FD.

PD is a good surrogate for FD across geographical areas. The figure presents the distribution and correlates of SPD–FD for mammals (a–c), birds (d–f), and tropical fishes (g–i) separately across space. For each of the three groups, the SPD–FD frequency distribution is presented in top panels (b, e, h) along with its mean (vertical line). The color scheme is common to all panels, with blue indicating positive SPD-FD (maximizing PD captures more FD than random) and red indicating negative SPD–FD. SPD–FD geographical distribution is presented in middle panels (a, d, g). Relationships between SPD–FD and species pool richness are presented in panels c, f, i. In each grid cell, SPD–FD values are based on the mean over 1000 repetitions of random and PDmax set draw (there is only one maxFD set). The maps in this figure were generated making use of the R-package rasterVis and latticeExtra. The animal silhouette images in this figure were created by the corresponding author in Adobe Acrobat

PD is a good surrogate for FD across clades. The figure presents the distribution and correlates of SPD–FD for mammals (a–c), birds (d–f) and fishes (g–i) across families. For each of the three groups, the SPD–FD frequency distribution is presented (b, e) along with its mean (vertical line). The color scheme is common to all panels. SPD–FD phylogenetic distribution is presented in panels a, d and g. Relationships between SPD–FD and species pool richness are presented in panels c, f, and i. For each taxonomic family, SPD–FD values are based on the mean over 1000 repetitions of random and maxPD set draw (there is only one maxFD set). The animal silhouette images in this figure were created by the corresponding author in Adobe Acrobat
Reliability of the surrogacy of PD for FD
Even though maximizing PD preserves more FD than averaged random selection, this averaged analysis does not capture the reliability of its performance. The PD maximization and the random selection strategies exhibit variation: simply by chance, random selection of species can capture very high (or, conversely, very low) FD, and the same may be true (to a previously unstudied degree) for PD. The extent of this variation is important: if it is less than the average difference, PD maximization is a reliable strategy as it will always yield more FD, but if it is not, then PD maximization could be unreliable for individual conservation interventions. To contrast these two situations, we measured the fraction of times that, within each species pool, the PD-maximization strategy yielded more FD than random selection (see Methods). PD-based selection was the best choice in 64% of cases (SD across species pool = 9%, see Supplementary Table 1 and Supplementary Figures 3 and 4), making it the better strategy but not a perfectly reliable one. Thus, while the PD-maximization strategy has a consistent positive effect (i.e., the average PD-maximization strategy yields more FD than the average random strategy), its effect is weak (i.e., the PD-maximization strategy still yields less FD than the random strategy in 36% of the trials within a species pool).
Drivers of surrogacy
We next explored the drivers of surrogacy values across species pools. Surrogacy of PD appears to weaken as the species pool richness increases (on average, Spearman Rho between absolute surrogacy and species richness = −0.15), most clearly seen in the tropics and in species-rich families such as the Muridae (rats, mice, and allies) and Columbidae (pigeons and allies) (Figs. 2 and 3). This is likely because our measure of FD (see Methods) rapidly saturates as the number of selected species increases and species from these large pools harbor high functional redundancy, such that a random prioritization scheme performs relatively well, or at least no worse than other strategies (Supplementary Figure 5). In contrast, FD can be greatly increased by prioritization of species using PD from species poor assemblages or clades. This is particularly the case in spatial assemblages containing multiple taxonomic orders, which are both phylogenetically and ecologically divergent from one another. Interestingly, the PD–FD relationship was not consistent across taxonomic scale: we found that, in contrast to patterns at the family level, for certain mammalian and avian orders (which are older than the families described above), using PD to select species is much worse for capturing FD than choosing species at random (see, for example, the Afrosoricidae, Chiroptera, and Charadriiformes in Supplementary Figure 6).
We then explored whether we can explain this variability within- and between-datasets, and in particular, why for some assemblages/clades, a PD-prioritization strategy fails to capture more FD than random choice. It is often implicitly assumed that phylogenetic signal (i.e., the degree to which closely related species tend to harbor similar sets of traits) can be used to evaluate the effectiveness of PD as a surrogate for FD5,15,16,17. Surprisingly perhaps, the value of PD as a surrogate for FD was only weakly correlated with the phylogenetic signal of the underlying traits (Supplementary Figure 7-8, on average Spearman Rho = 0.17). Similarly, tree imbalance, which is known to affect surrogacy in simulations16, did not explain surrogacy in these empirical data (Supplementary Figures 7 and 8).
For mammals, regions where PD did worse than random were located in the Sahara, south western Patagonia, southern Africa including parts of Madagascar, and New Guinea (Fig. 2). These latter two in particular are of concern since they are global conservation priorities on the basis of species endemism and habitat loss. We suggest two historical reasons for such idiosyncratic poor performance of PD. First, there is a tendency for a large carnivore species, either a top predator (e.g., cheetahs in the Sahara or foxes in Patagonia) or a large scavenger (e.g., the hyena in South Africa) to co-occur with a close relative with distinct traits in these areas (e.g., a desert cat with the cheetah or the aardwolf with the hyena, see Supplementary Figure 9). Only one of these closely related species will tend to be selected under prioritization schemes that maximize PD, thus reducing the volume of the convex hull on average when the functionally distinct one is not selected (the large predator or scavenger). This seems also to drive the low surrogacy of PD in Charadriiformes (especially Larus and Sterna; see Supplementary Figure 10). Second, lineages in which traits evolve very slowly will contribute little to FD, even over long periods of time (branch lengths) that contribute greatly to PD. For example, in New Guinea many co-occurring bats with similar traits diverged long ago, such that they are always selected in the PD maximizing set, but do not add much to the convex hull, resulting in a poor surrogacy of PD for FD. Such strong ecological niche conservatism is common in mammals29, e.g., in the Geomyidae: two basal branches of the Geomyidae tree harbor very similar traits (species descending from these branches are actually grouped in the same genus Thomomys) while being distantly related in the phylogenies we used (Supplementary Figure 9). As such, they will be selected in all PD maximizing sets, but will not contribute greatly to FD.
Discussion
Maximizing PD in conservation decisions is now commonplace in the academic world20,21,22,23,30,31,32 and is also starting to be used in real-world conservation prioritizations, for example with the EDGE of existence program18. To the best of our knowledge, there are no clear direct ecosystem function or health benefits that phylogenetic branch lengths provide. Rather, high PD is perceived as valuable because it is assumed to be a good proxy for high diversity of traits or “features”14 (referred as to high functional diversity in this paper, FD), a hypothesis that we name the “phylogenetic gambit”. High FD might be valuable for a number of reasons, for example ecosystem functioning, ecosystem services, future “options values”14,15 or “evolutionary potential”15,33. The utility of PD for conservation stems from the fact that calculating PD is relatively fast and cheap, often making it an easier way to prioritize species or areas than FD. Indeed, we have imperfect knowledge about which, and how many, traits and functions are important in a given context, how these traits and functions vary among species and across space, and how the importance of traits may change in the future13. Yet, even if convenient, maximizing PD can only be an effective and realistic conservation strategy to conserve FD if the phylogenetic gambit holds and maximizing PD yields more FD than a strategy that ignores phylogeny. If maximizing PD yields less FD than a random strategy (i.e., the gambit fails), then researchers and conservationists should reconsider whether maximizing PD as a useful conservation strategy. A large body of literature has shown that maximizing PD does not maximize FD empirically20,21,22,23 or even in simple theoretical cases16, but such work does not test the phylogenetic gambit of whether PD prioritization captures more FD than random selection16. Here we have shown that the phylogenetic gambit generally holds: PD is an effective conservation metric to capture FD. Yet we also reveal some limitations of this strategy: PD is good “on average”, but there is still some risk associated with taking it.
We found that prioritizing the most phylogenetically diverse set of taxa in a region or clade will result in an average gain of 18% functional diversity relative to applying the same conservation effort without considering phylogeny, but this gain will decrease as species richness increases. In contrast to what has previously been implicitly assumed15,16, we find weak empirical evidence that the presence of phylogenetic signal in traits predicts whether PD-based conservation will prioritize FD. Our result suggests that PD is a reasonable conservation prioritization strategy, especially in species-poor clades or regions, or in the absence of meaningful data on functional traits. However, we note three important caveats to the use of this strategy. First, 18% extra FD may not always be a useful conservation target. It is currently unknown whether this added 18% of FD can actually be of enough conservation value. Second, in cases of either recent trait divergence or, alternatively, very strong trait conservatism, a PD prioritization scheme can capture less FD than a random scheme. Evolutionary biologists commonly focus on “unusual” clades with rapid divergences (e.g., cichlids); we show here that divergence does not have to be that spectacular (e.g., African carnivores) to alter the PD–FD relationship. Third, we found that while this strategy, on average, captures FD well, it is also somewhat unreliable, and 36% of the time will not capture more FD than random choice. This means that while the PD gambit can be a bet worth taking, it is still a bet with associated risk, not a sure thing.
Our objective in this paper is to test the phylogenetic gambit using empirical datasets. This means that we do not aim to provide a coherent prioritization strategy34, or ready-to-use conservation guidelines. Indeed, we simplistically and implicitly assume that species will either be saved or go extinct, and we have not linked our various scenarios to any particular policy position or conservation objective other than maximizing FD within a phylogenetic clade or region28,30. In reality, conservation decisions reflect the interplay of social, economic, political, and scientific priorities, and do not necessarily result in the saving of target species (and therefore of their associated FD or PD). While our study is thus not directly applicable, the test we are conducting is actually critical to validate (or invalidate) the use of PD in conservation as a whole. While it is not clear whether our results would generalize to other taxa (although we hope that others will extend our work and test the phylogenetic gambit in other systems), we do feel it is important to consider the uncertainty that has been introduced into our analysis as a result of uncertainty associated with the spatial scale of our analysis, our phylogenetic data, and our choice of trait and measurement of FD.
The scale of conservation activities can vary, from the global scale of the hotspots approach to local protected areas within a single country, but, unfortunately, the connection between these scales remains unclear. For example, if the motivation for protecting FD is to maintain community-driven ecosystem functions and services5,6,35, the value of a regional or global focus may be questionable36; studies are increasingly focusing on local scales6. Ecologists are refining and improving our understanding of how local assemblages assemble within a regional context37, and while the concept of the “regional pool” of species is increasingly being viewed as a simplification, it is unlikely that regional- and local-scale patterns are totally disconnected. We emphasize that our results are relatively robust to variation in spatial scale (see Supplementary Fig 3), but we acknowledge that future studies should test the phylogenetic gambit at more local scale as well.
The set of species that maximize PD obviously relies on the phylogenetic hypothesis used. No hypothesis is perfect or without uncertainty, and these phylogenetic uncertainties could in turn impact the composition of the set of species that maximize PD and hence the surrogacy values we compute. In this study, we explicitly took into account these uncertainties by using 100 different trees38,39. The explicit propagation of this phylogenetic uncertainty through into our results may underlie some of the uncertainty (risk) of our result for birds and mammals, and we suggest that future studies explicitly take into account phylogenetic uncertainty when testing the phylogenetic gambit.
The motivator of our test of the surrogacy value of PD for FD is the fact that ecologically relevant trait data is in short supply, especially for rare and data-deficient species. Indeed, if it were not for this relative paucity of data, we could simply prioritize species based on their unique contribution to FD directly. Although there have been massive and well-funded efforts to collect and curate trait data from across the Tree of Life40,41,42, we are still far from having comprehensive coverage. Furthermore, despite recent progress43, it is still not fully understood which traits are most relevant for responses to environmental change, or that contribute most to certain ecosystem functions and services, and how these vary among systems. Our analysis suffers from a similar data limitation. We chose these traits because they are frequently collected in ecological studies, not because we know they are ecologically important. Our assumption is that their phylogenetic distribution is typical of those traits that are most desirable for the purpose of conservation and that our primary results are therefore widely applicable. While we did test the robustness of our results to the variation of trait information retained to compute FD (Supplementary Figure 10), it is true that, overall, we used a rather limited set of traits. We acknowledge that it is possible that many other potential valuable traits are not captured by our measure of FD. One of the ideas behind the use of PD is that phylogeny might account for these for unmeasured and unmeasurable traits9,14,15, however, as this hypothesis is not testable (we do not have these traits), it seems risky to assume it is true. Our objective here is to test the phylogenetic gambit given the limited set of traits that we have: we consider that carrying out our imperfect test is more informative than not carrying any test at all.
In conclusion, we found that maximizing PD results in an average gain of 18% of FD relative to random choice. However, this average gain hides the fact that in over 1/3 of the comparisons, maximum PD sets contain less FD than randomly chosen sets of species. These results suggest that, while maximizing PD can help capture FD, it represents a risky strategy. If maximizing PD is a risky strategy, then, should we abandon the use of PD in conservation? We believe that before such dramatic decision, our test should be repeated across space, traits and taxa, in order to narrow the uncertainties of our results. This is why we now urge others to expand our simple phylogenetic gambit test to other clades and other traits in order to test the generality of our findings. We hope that our study will stimulate the production of numerous tests to finally rigorously assess the usefulness of PD in conservation.
Methods
Type of data
We use two classes of data to address the question of whether choosing sets of species according to PD captures the underlying trait diversity (as measured with FD) well. First, we used taxonomic groups (clades) of species as our unit of analysis (“species pool” hereafter) and, second, we investigated broad assemblages found across the globe. The former is more explicitly evolutionary, ensuring that our results are not driven by well-established relationships across large taxonomic groups (e.g., monotremes are distinct from placental mammals) and the latter is likely more relevant to actual conservation practice. We use distribution data to delineate geographical assemblage species pool and taxonomy to delineate clade-based species pools (namely families and orders).
Distribution data
For mammals, we used the distribution maps provided by the Mammal Red List Assessment (http://www.iucnredlist.org/) for 4616 species. For birds, full (breeding and wintering) and breeding ranges distribution maps were extracted from BirdLife (http://www.birdlife.org/) for 9993 species. The best resolution at which these maps should be used is still under discussion in the literature, so we decided to use the 40,000 km2 resolution (200 × 200 km gird cell at the equator) that is commonly used at global scale44,45. The total number of grid cells was 3646. Domestic and aquatic mammals were excluded from the analysis. In order to make sure our results were not driven by the important trait difference between volant and nonvolant mammals, we repeated our results excluding bats. For birds, we repeated our analysis using the full ranges. Finally, we evaluated the robustness of our result to the spatial resolution considered by repeating our analysis at a resolution of 100 × 100 km (number of cells was 13,330) for birds and mammals; we present these results in the supplementary materials, as they are qualitatively identical to those conducted at 200 × 200 km (Supplementary Figure 1). For fishes, we used a database of 1536 species, for which we had distribution data, phylogenetic and functional data. Distribution data were extracted from a global-scale distribution database46. Species composition was then extracted from grid cells of 5°x5°, corresponding to approximately 555 × 555 km at the equator47. This grain size of the grid was chosen because it represents a good compromise between the desired resolution and the geographical density of information. Fish distribution data are available upon request to DM and FL. Maps were handled and plotted in R using the packages rasterVis48 and latticeExtra49.
Phylogenetic data
In order to prioritize species to maximize PD, phylogenies of each species pool are needed. We used the first 100 published calibrated ultrametric trees of Jetz et al.39 for birds and of Faurby and Svenning38 for mammals. By repeating our analyses across a posterior distribution of phylogenetic hypotheses, we control and account for phylogenetic uncertainty. For tropical reef fishes, we built a phylogeny for 18 families (i.e., Labridae, Scaridae, Pomacentridae, Chaetodontidae, Acanthuridae, Haemulidae, Balistidae, Carangidae, Serranidae, Lutjanidae, Sparidae, Caesionidae, Holocentridae, Mullidae, Muraenidae, Tetraodontidae, Lethrinidae, and Siganidae) by pruning a dated molecular phylogenetic tree for 7822 extant fish species47. These families were selected as the most representative tropical reef fish families, that is, they are abundant and speciose on tropical reefs. We grafted missing species on the pruned phylogenetic tree (circa 50% among the 1536 studied species) based on published phylogenies for these families, supplemented by taxonomic information from fish identification guides and FishBase47,50. The corresponding tree is available on figshare (https://doi.org/10.6084/m9.figshare.6430982.v1). We recorded, for each of these trees, a measure of imbalance (as measured by beta51) and “tipiness” (as measured by gamma52). For both mammals and birds, we chose to group species in families and orders. We used these groupings when calculating the purely phylogenetic, clade-based analyses, but not within the spatial, assemblage-based analyses. For the taxonomic analysis of mammal families, we removed two families (Dipodidae and Echimyidae) because of their very poor phylogenetic resolution (i.e., polytomies for an important number of species).
Trait data
For birds and mammals, four traits (diet, (log-transformed) body mass, activity cycle, and foraging height) were extracted from Elton Traits1.042. These traits are generally assumed to appropriately represent Eltonian niche dimensions within an assemblage or clade of mammals or birds53,54. For fishes, we used a previously published database12. We used 6 categorical traits: size, mobility, period of activity, schooling, vertical position in the water column, and diet (for a full description of the dataset, see Mouillot et al.12). These traits have already been used to investigate community assembly rules55 and to seek vulnerable fish functions11. Fish trait data is available upon request to DM and FL. For each clade and assemblage, we used the raw trait (only body mass was log-transformed and rescaled by the clade/assemblage range of body masses) values to compute distances between species using Gower distance and use PCoA to summarize the trait space in few dimensions. We retained the numbers of PCoA axes necessary to represent 70% of the total initial variability (using a 80% threshold did not quantitatively change our conclusions, see Supplementary Figure 10). We also recorded phylogenetic signal for each PCoA axis using Blomberg’s K56.
General approach
Our aim was to evaluate, across a wide range of clades and regions, the ability of PD-informed prioritization scheme to capture FD in comparison with two other prioritization schemes: selecting species to directly maximize FD (“maxFD” hereafter) and selecting species randomly (Fig. 1). Our premise was that we often do not know or have not measured the traits that are most relevant for ecosystem function and services such that maximizing FD is not generally feasible. By focusing on a subset of traits and assuming that they are representative of ecologically relevant traits, we were able to get an estimate of how well PD does compared to the best we could possibly do. We used performance relative to choosing on the basis of FD as an upper-limit to the performance of PD as a surrogate for FD and used random species selection as a lower benchmark.
Random prioritization scheme
For each pool (i.e., each clade and each geographical assemblage) and each number of selected species (10, 20, 30, 40, 50, 60, 70, 80, 90, and 100% of the total pool), 1000 random sets of species were produced, from which the average FD was recorded.
Prioritization scheme maximizing PD (maxPD)
While there are many, overlapping metrics for measuring the evolutionary history encompassed by a set of species15,57, the most common is the sum of all branch lengths (often in units of time) connecting a set of species to a common root14, called phylogenetic diversity (PD). This is the metric whose maximization has most commonly been proposed as a conservation prioritization metric14,33,58, and as a measure of phylogenetic “richness” it most naturally maps onto our chosen FD metric57. We used the greedy algorithm proposed by Bordewich et al.59 to find our maxPD set of species S. For a given tree there are likely multiple, and possibly very many, sets of species with the same PD as S. As a consequence, we produced, for each pool, each number of selected species, and each alternative phylogenetic trees, 10 maxPD sets of species. We then averaged the FD of these sets across our 100 phylogenetic trees, so that each value is an average of 1000 sets (10 sets for each of the 100 trees).
Prioritization scheme maximizing FD (maxFD)
Functional diversity was estimated using a functional richness index (FRic)60,61,62. The FRic index relies on a multidimensional Euclidean space, where the axes are traits (or factorial axes from a principal coordinates analysis (PCoA) computed using these traits) along which species are placed according to their trait values. This index measures the volume of trait space occupied by a given species assemblage by calculating the convex hull volume62, defined by the species at the vertices of the functional space, that encompasses the entire trait space filled by all species in this assemblage. In a single dimension, this simply equals the range of values62. This broadly used metric in ecology is set monotonic with species richness, a property generally assumed desirable in conservation whereby the addition of a new species can never decrease the metric’s value63. FD measures the total amount of variation in trait values, making it conceptually comparable to PD57. We used the FRic index instead of the FD index based on a functional dendrogram since recent studies showed that the FD index may lead to biased assessments of functional diversity and inaccurate ecological conclusions64. The most straightforward way to obtain the maximal FD for n species is to compute FD for all possible combinations of n species and simply record the greatest value (the brute force approach). However, this is not feasible in practice as the numbers of combinations of selected species was too high (e.g., 1071 possible sets for all mammal assemblages). To rapidly and efficiently find the set of species that aim to maximize FD, we developed a novel (at least in ecology) greedy algorithm. In brief, our approach iteratively (starting with two species) select the species that is the furthest from the centroid of the already selected set. To avoid selecting two species that are far from the centroid but close to each other, we penalized the distance to the centroid by the distance to the closest neighbor in the already selected set. Here we present in details the greedy algorithm we used to find the set of species that maximize FD:
Step 1. Select the two species with the highest trait distance.
Step 2. Compute the centroid of these two selected species.
Step 3. Compute distances between species not in the set and this “set centroid”.
Step 4. Penalize these distances by adding the following factor f (Eq. 1)
with K and L being penalizing factors and minD the distance between a given candidate species and the nearest species already in the selected set.
Step 5. Select the species that maximized the penalized distance.
Step 6. Go back to step one with this new set of species until the desired number of species is reached.
To avoid arbitrarily setting the penalizing parameters, we tested 1000 pairs of parameters drawn from a truncated normal distribution (mean = 1, SD = 0.5) and retained the parameter pairs that yielded the maximal FD.
In tests of subsets of the data for which finding the true maxFD was feasible, we found our approach to adequately approximate the true maxFD and to produce a very good approximation of the true degree of PD’s surrogacy for FD (Supplementary Figure 11).
Surrogacy estimates
We use a common approach27,28 to quantify the extent to which a given surrogate (here, the maxPD choice) reaches a certain objective (here, maximize FD). Species from a given pool (i.e., for each dataset (clade and assemblages) independently,) were prioritized and selected according to (1) the objective, i.e., maximize FD, producing the “optimal curve” (maxFD curve in Fig. 1) the surrogate, i.e., maximize PD, producing the “surrogate curve” (maxPD curve in Figs. 1 and 3) at random (random curve in Fig. 1), i.e., producing the “random curve” (Fig. 1). To compute a “surrogacy” estimate of PD (SPD–FD), we compare the position of the surrogate curve (1) to the random curve (2) relative to the optimal curve (2) (Fig. 1 and Eq. 2) across the deciles of species richness of the pool (given as an interval 0–1):
This surrogacy metric is at 100% when the surrogate perfectly meets the objective (i.e., the maxFD and maxPD curves are identical and the max PD set is the maxFD set), 0% when the surrogate is not better than randomly chosen sets of species (i.e., the random and maxPD curves are identical) and is negative if the surrogate choice is worse than random (i.e., the maxPD curve is below the random curve). Correlates of SPD–FD were evaluated using Spearman correlations.
Apart from focusing on average tendencies, we quantified the variability of the FD yielded by the PD—maximized selection strategy and the random selection strategy within each species pools. To do so, we compute, for each species pool and for each % of selected species independently, the number of cases where FDrandom > FDmaxPD across the 1000 random *1000 maxPD sets combinations (i.e., 106 comparisons). We then averaged theses number across % of selected species and report statistics across datasets (Supplementary Table 1, Supplementary Figs. 3 and 4).
Code availability
R functions developed in this paper are available at https://github.com/FloMazel/FD_PD_Max
Data availability
Mammal and bird datasets are publicly available (see methods). The Fish phylogeny is available on figshare (https://doi.org/10.6084/m9.figshare.6430982.v1). The distribution and trait fish datasets are available upon request to D.M. and F.L.
Change history
04 February 2019
The original version of this Article contained a plotting error in Figure 3g. The Serranidae and Siganidae families were misplaced in the plotted phylogeny. This error has now been corrected in the PDF and HTML versions of the Article. For comparison, the original, incorrect version of Figure 3g is presented in the associated Author Correction.
References
Pimm, S. L. et al. The biodiversity of species and their rates of extinction, distribution, and protection. Science (80-). 344, 1246752 (2014).
Hull, P. M., Darroch, S. A. F. & Erwin, D. H. Rarity in mass extinctions and the future of ecosystems. Nature 528, 345–351 (2015).
Ceballos, G., Ehrlich, P. R. & Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl Acad. Sci. USA 114, E6089–E6096 (2017).
Tilman, D., Wedin, D. & Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379, 718–720 (1996).
Srivastava, D. S. D., Cadotte, M. M. W., MacDonald, A. A. M., Marushia, R. G. & Mirotchnick, N. Phylogenetic diversity and the functioning of ecosystems. Ecol. Lett. 15, 637–648 (2012).
Cadotte, M. W., Carscadden, K. & Mirotchnick, N. Beyond species: functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 48, 1079–1087 (2011).
Isbell, F. et al. Linking the influence and dependence of people on biodiversity across scales. Nature 546, 65–72 (2017).
Pascual, U. et al. Valuing nature’s contributions to people: the IPBES approach. Curr. Opin. Environ. Sustain 26–27, 7–16 (2017).
Vane-Wright, R. I., Humphries, C. J. & Williams, P. H. What to protect?—Systematics and the agony of choice. Biol. Conserv. 55, 235–254 (1991).
Bottrill, M. C. et al. Is conservation triage just smart decision making? Trends Ecol. Evol. 23, 649–654 (2008).
Mouillot, D. et al. Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol. 11, e1001569 (2013).
Mouillot, D. et al. Functional over-redundancy and high functional vulnerability in global fish faunas on tropical reefs. Proc. Natl. Acad. Sci. USA 111, 13757–13762 (2014).
Cadotte, M. W., Davies, J. T. & Peres-Neto, P. R. Why phylogenies do not always predict ecological differences. Ecol. Monogr. https://doi.org/10.1002/ecm.1267 (2017).
Faith, D. P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 61, 1–10 (1992).
Winter, M., Devictor, V. & Schweiger, O. Phylogenetic diversity and nature conservation: where are we? Trends Ecol. Evol. 28, 199–204 (2013).
Mazel, F., Mooers, A. O., Riva, G. V. D. & Pennell, M. W. Conserving phylogenetic diversity can be a poor strategy for conserving functional diversity. Syst. Biol. 66, 1019–1027 (2017).
Kelly, S. & Grenyer, R. & Scotland, R. W. Phylogenetic trees do not reliably predict feature diversity. Divers. Distrib. 20, 600–612 (2014).
Isaac, N. J. B., Turvey, S. T., Collen, B., Waterman, C. & Baillie, J. E. M. Mammals on the EDGE: conservation priorities based on threat and phylogeny. PLoS ONE 2, e296 (2007).
Barak, R. S. et al. Taking the long view: integrating recorded, archeological, paleoecological, and evolutionary data into ecological restoration. Int. J. Plant Sci. 177, 90–102 (2016).
Pollock, L. J., Thuiller, W. & Jetz, W. Large conservation gains possible for global biodiversity facets. Nature 546, 141–144 (2017).
Brum, F. T. et al. Global priorities for conservation across multiple dimensions of mammalian diversity. Proc. Natl Acad. Sci. USA 114, 7641–7646 (2017).
Mazel et al. Multifaceted diversity–area relationships reveal global hotspots of mammalian species, trait and lineage diversity. Glob. Ecol. Biogeogr. 23, 836–847 (2014).
Thuiller, W. et al. Conserving the functional and phylogenetic trees of life of European tetrapods. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370, 20140005 (2015).
Mason, N. W. H. & Pavoine, S. Does trait conservatism guarantee that indicators of phylogenetic community structure will reveal niche-based assembly processes along stress gradients? J. Veg. Sci. 24, 820–833 (2013).
Gerhold, P., Cahill, J. F., Winter, M., Bartish, I. V. & Prinzing, A. Phylogenetic patterns are not proxies of community assembly mechanisms (they are far better). Funct. Ecol. 29, 600–614 (2015).
Devictor, V. et al. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world. Ecol. Lett. 13, 1030–1040 (2010).
Ferrier, S. & Watson, G. An evaluation of the effectiveness of environmental surrogates and modelling techniques in predicting the distribution of biological diversity. (NSW National Parks and Wildlife Service for the Department of Environment, Sport and Territories, Canberra, 1997).
Rodrigues, A. S. L. & Brooks, T. M. Shortcuts for biodiversity conservation planning: the effectiveness of surrogates. Annu. Rev. Ecol. Evol. Syst. 38, 713–737 (2007).
Mazel, F. et al. The geography of ecological niche evolution in mammals. Curr. Biol. 27, 1369–1374 (2017).
Carvalho, S. B. et al. Spatial conservation prioritization of biodiversity spanning the evolutionary continuum. Nat. Ecol. Evol. 1, 0151 (2017).
Jetz, W. et al. Global distribution and conservation of evolutionary distinctness in birds. Curr. Biol. 24, 919–930 (2014).
Thévenin, C., Mouchet, M., Robert, A., Kerbiriou, C. & Sarrazin, F. Reintroductions of birds and mammals involve evolutionarily distinct species at the regional scale. Proc. Natl Acad. Sci. USA 115, 3404–3409 (2018).
Forest, F. et al. Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 445, 757–760 (2007).
Brooks, T. M. et al. Global biodiversity conservation priorities. Science 313, 58–61 (2006).
Díaz, S. et al. Incorporating plant functional diversity effects in ecosystem service assessments. Proc. Natl Acad. Sci. USA 104, 20684–20689 (2007).
Srivastava, D. S. & Vellend, M. Biodiversity-ecosytem function research: is it relevant to conservation? Annu. Rev. Ecol. Evol. Syst. 36, 267–294 (2005).
HilleRisLambers, J., Adler, P. B., Harpole, W. S., Levine, J. M. & Mayfield, M. M. Rethinking community assembly through the lens of coexistence theory. Annu. Rev. Ecol. Evol. Syst. 43, 227–248 (2012).
Faurby, S. & Svenning, J.-C. A species-level phylogeny of all extant and late quaternary extinct mammals using a novel heuristic-hierarchical Bayesian approach. Mol. Phylogenet. Evol. 84, 14–26 (2015).
Jetz, W., Thomas, G., Joy, J. & Mooers, A. Ø. The global diversity of birds in space and time. Nature 491, 444–448 (2012).
Kattge, J. et al. TRY--a global database of plant traits. Glob. Chang. Biol. 17, 2905–2935 (2011).
Jones, K. E. et al. PanTHERIA: a species-level database of life history, ecology, and geography of extant and recently extinct mammals. Ecology 90, 2648 (2009).
Wilman, H. et al. EltonTraits 1.0: species-level foraging attributes of the world’s birds and mammals. Ecology 95, 2027 (2014).
Lavorel, S. & Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Funct. Ecol. 16, 545–556 (2002).
Hurlbert, A. H. & Jetz, W. Species richness, hotspots, and the scale dependence of range maps in ecology and conservation. Proc. Natl Acad. Sci. USA 104, 13384–13389 (2007).
Holt, B. G. et al. An update of Wallace’s zoogeographic regions of the world. Science (80-) 339, 74–78 (2013).
Parravicini, V. et al. Global patterns and predictors of tropical reef fish species richness. Ecography (Cop) 36, 1254–1262 (2013).
Leprieur, F. et al. Plate tectonics drive tropical reef biodiversity dynamics. Nat. Commun. 7, 11461 (2016).
Lamigueiro, O. P. & Hijmans, R. meteoForecast. R package version 0.41. (2016).
Sarkar, D. & Andrews, F. latticeExtra: Extra Graphical Utilities Based on Lattice. R package version 0.6-28.
Ottimofiore, E. et al. Responses of coral reef fishes to past climate changes are related to life-history traits. Ecol. Evol. 7, 1996–2005 (2017).
Blum, M., François, O. & Steel, M. Which random processes describe the tree of life? A large-scale study of phylogenetic tree imbalance. Syst. Biol. 55, 685–691 (2006).
Pybus, O. & Harvey, P. Testing macro–evolutionary models using incomplete molecular phylogenies. Proc. R. Soc. Lond., Ser. B 267, 2267–2272 (2000).
Safi, K. et al. Understanding global patterns of mammalian functional and phylogenetic diversity. Philos. Trans. R. Soc. Lond. Ser. B 366, 2536–2544 (2011).
Belmaker, J. & Jetz, W. Relative roles of ecological and energetic constraints, diversification rates and region history on global species richness gradients. Ecol. Lett. 18, 563–571 (2015).
Guillemot, N. et al. Functional redundancy patterns reveal non-random assembly rules in a species-rich marine assemblage. PLoS ONE 6, e26735 (2011).
Blomberg, S. P., Garland, T. J. R. & Ives, A. Testing for phylogenetic signal in compartive data: behavioral traits are more labile. Evolution 57, 717–745 (2003).
Tucker, C. M. et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. Camb. Philos. Soc. https://doi.org/10.1111/brv.12252 (2017).
Rodrigues, A. S. L. & Gaston, K. J. Maximising phylogenetic diversity in the selection of networks of conservation areas. Biol. Conserv. 105, 103–111 (2002).
Bordewich, M., Rodrigo, A., Semple, C. & Collins, T. Selecting taxa to save or sequence: desirable criteria and a greedy solution. Syst. Biol. 57, 825–834 (2008).
Villéger, S., Mason, N. & Mouillot, D. New multidimensional functional diversity indices for a multifaceted framwork in functional ecology. Ecology 89, 2290–2301 (2008).
Pavoine, S. & Bonsall, M. B. Measuring biodiversity to explain community assembly: a unified approach. Biol. Rev. 86, 792–812 (2011).
Cornwell, W. K., Schwilk, L. D. & Ackerly, D. D. A trait-based test for habitat filtering: convex hull volume. Ecology 87, 1465–1471 (2006).
Ricotta, C. A note on functional diversity measures. Basic Appl. Ecol. 6, 479–486 (2005).
Villéger, S., Maire, E. & Leprieur, F. On the risks of using dendrograms to measure functional diversity and multidimensional spaces to measure phylogenetic diversity: a comment on Sobral et al. (2016). Ecol. Lett. 20, 554–557 (2017).
Acknowledgements
This paper is a joint effort of the sCAP working group funded by sDIV, the Synthesis Centre of the German Centre for Integrative Biodiversity Research Halle-Jena-Leipzig and the Canadian Institute for Ecology and Evolution. F.M. was supported by an NSERC Accelerator Grant to A.O.M. and a Banting Post-doctoral fellowship. G.V.D.R. thanks Mike Steel. We thank Risa Sergeant at the University of Ottawa for hospitality, the Crawford Lab for Evolutionary Studies at SFU for discussion. M.W.C., D.M., and C.M.T. are members of the FREE working group at the CESAB-FRB.
Author information
Authors and Affiliations
Contributions
F.M., M.W.P., M.W.C., S.D., G.V.D.R., R.G., A.O.M., C.M.T., and W.D.P. conceived the design of the study. F.M. and G.V.D.R. conducted the analysis. F.M., R.G., M.P., and W.D.P. interpreted the results and wrote the first draft of the manuscript. F.M., F.L., D.M., M.W.P., M.W.C., S.D., G.V.D.R., R.G., A.O.M., C.M.T., and W.D.P. edited the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mazel, F., Pennell, M.W., Cadotte, M.W. et al. Prioritizing phylogenetic diversity captures functional diversity unreliably. Nat Commun 9, 2888 (2018). https://doi.org/10.1038/s41467-018-05126-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-018-05126-3
This article is cited by
-
Elevated alpha diversity in disturbed sites obscures regional decline and homogenization of amphibian taxonomic, functional and phylogenetic diversity
Scientific Reports (2023)
-
Global hotspots of traded phylogenetic and functional diversity
Nature (2023)
-
Number of forest fragments and understory plants exert opposite effects on multiple facets of bird diversity in eucalypt plantations
Landscape Ecology (2023)
-
The expected loss of feature diversity (versus phylogenetic diversity) following rapid extinction at the present
Journal of Mathematical Biology (2023)
-
Native diversity buffers against severity of non-native tree invasions
Nature (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
