
Abstract
Flowering is the most important event in higher plants. Compared to most fruit tree species, Chinese jujube (Ziziphus jujuba Mill.), the most important member of the large, diverse Rhamnaceae family and a leading dry fruit-producing species, has unique characteristics that include a short juvenile phase and extremely fast flower bud differentiation. However, the distinct mechanism of flowering regulation in Chinese jujube is still unclear. The morphological and cytological development period of jujube flowering was first investigated, and the crucial developmental stages were defined. Flower bud differentiation in Chinese jujube took only approximately 11–13 days, which is a distinct characteristic of perennial fruit trees. Afterward, 44 genes related to six flowering pathways were identified in the jujube genome and were found to be randomly distributed among 11 of the 12 chromosomes. Tissue-specific and spatiotemporal expression patterns showed that all these genes were expressed in the flowers. Overall, photoperiod-related genes were highly expressed during flower bud differentiation. These genes were also positively responsive to photoperiod regulation and phase change processes, indicating that photoperiod- related genes play crucial roles in jujube flower bud differentiation. Under protected cultivation, ZjPIF4, a temperature-related gene, was expressed in the early stages of flowering and responded to increasing temperatures. Moreover, STRING analysis and yeast two-hybrid screening indicated that photoperiod-related (ZjCO) and temperature-related (ZjPIF4) proteins could interact with ZjFT, the key protein involved in the determination of flowering time, indicating crosstalk between photoperiod-related pathways and ambient temperature-related pathways in jujube. This study is the first report to comprehensively analyze the flowering pathways in Chinese jujube and revealed that photoperiod-related and ambient temperature-related pathways are the main mechanisms regulating the distinct flowering process and that members of the ZjPHY family (ZjPIF4, ZjFT, and ZjCO5) are the key factors involved in the regulatory network. These results will increase our understanding of the molecular and genetic mechanisms of flowering in Chinese jujube and provide meaningful clues for the flowering regulation of other fruit tree species.
Similar content being viewed by others

Introduction
The flowering process in higher plants is the end result of a central hub controlling the transformation from the vegetative growth stage to the reproductive growth stage. Flowering time has an important impact on the breeding, early fruiting and high yields of perennial woody fruit trees. An intricate network involving various (epi-) genetic regulators controls the timely onset of flowering. A series of genes related to flowering have been isolated and identified from model plant species and woody fruit tree species1,2.
Flowering involves complex regulatory mechanisms and pathways that ensure flower transformation at the appropriate time. In Arabidopsis, there are mainly six regulatory pathways, i.e., the vernalization-related, photoperiod-related, gibberellin-related, autonomous-related, ambient temperature-related, and age-related pathways3. These pathways are independent of each other but exhibit crosstalk to form a flower regulatory network with precise regulatory functions4. According to previous studies, the flowering process is affected by external environmental conditions and internal growth and development. However, the flowering pathways of most fruit tree species are still unclear.
Chinese jujube (Ziziphus jujuba Mill.) is the most important member of the large, diverse Rhamnaceae family and one of the most economically important fruit tree species in China5. In general, most fruit trees have long juvenile vegetative phases. Compared to most other fruit trees, Chinese jujube trees have a very short juvenile phase and the ability to produce fruit within the first year of planting, with flower bud differentiation and fruiting occurring in the same year. In our previous study, several MADS-box genes related to flowering were identified6. However, there have been no reports on the jujube flowering pathway. Therefore, to clarify the main pathways regulating jujube flowering and the critical genes involved in jujube flowering, the morphology and developmental period of jujube flowers were observed. In addition, the genes related to different flowering pathways were identified at the genomic level, and their involvement in the flowering process was further analyzed. These results will increase our understanding of the molecular and genetic mechanisms underlying flowering in Chinese jujube and provide clues for flowering regulation in other fruit tree species.
Results
Morphological and cytological observations of flower development in Chinese jujube
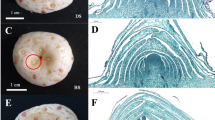
To determine the critical sampling periods, we systematically observed the entire flowering process of Chinese jujube. In general, there are three stages during the plant flowering process: sex determination, flower bud differentiation, and floral organ development. From sex determination to flower bud differentiation, we divided the process into six stages at the histological level: the undifferentiated stage, initial flower bud differentiation, sepal differentiation, petal differentiation, stamen differentiation, and carpel differentiation (Fig. 1a). Correspondingly, the morphology and developmental period of the bearing shoots were also investigated (Table S1).

a (Flower bud differentiation). A1: Undifferentiated period; A2: Initial stage; A3: Sepal differentiation; A4: Petal differentiation; A5: Stamen differentiation; A6: Carpel differentiation; Bar: 50 µm. b (Flower development). B1: Bud emergence; B2: Inflorescence emergence; B3: Bud flattening; B4: Bud swelling; B5: Bud yellowing. Bar: 500 µm. c (Flower opening). C1: Bud break; C2: Early blooming; C3: Semiopening; C4: Petal elongation; C5: Petal flattening; C6: Stamen flattening; C7: Wilting; Bar: 1000 µm
At the undifferentiated stage, two bracts appeared on both sides of the leaf axils, and no obvious convex apical meristem was observed (Fig. 1A1, Table S1). When flower bud differentiation began, the top of the bud gradually flattened to a pointed circle, forming a flower primordium (Fig. 1A2). It took approximately 1.5–2 days for the transition from the initial stage to sepal differentiation. Compared to that of other stages, differentiation from sepals to petals was the longest stage (approximately 6–7 days, Table S1). The protuberance became a rudimentary dichotomous stigma (Fig. 1A6), and a flower bud was visible between the leaf axils. In total, the whole process of flower bud differentiation in Chinese jujube took only approximately 11–13 days, and the extremely fast differentiation of flowers is a distinct characteristic of perennial fruit tree species.
Flower development began with the enlargement of the flower bud. Accordingly, we divided flower development into five stages: bud emergence, inflorescence emergence, bud flattening, bud swelling, and bud yellowing (Fig. 1b). With the development of flower buds in bearing shoots, the bud became lighter in color. When the diameter of the bud reached approximately 2600 µm, the bud shape changed from round to flat. The bud color ultimately became yellow-green or light-yellow (Fig. 1b). The bud development period of Chinese jujube lasted 25–34 days.
We also observed the single-flower opening process of Chinese jujube and divided it into seven stages: bud splitting, early opening, sepal flattening, the petal upright stage, petal flattening, stamen flattening, and wilting (Fig. 1c). The single-flower opening process was short and took only approximately 2–3 days.
Identification of candidate genes related to jujube flower bud differentiation
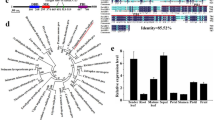
After performing a homology search, we identified 44 candidate genes related to flower bud differentiation for six flowering pathways in Chinese jujube (Table 1). The ORF of these genes ranged from 222 to 3900 bp, and their encoded proteins ranged from 83 to 1299 amino acids (aa), with a predicted molecular mass of 8.16–144.81 kDa and a pI ranging from 4.34 to 10.81. Moreover, these genes were mapped to the 11 pseudochromosomes (Fig. 2). Most members of the CONSTANS family were concentrated on Chromosome (Chr) 4, but ZjCO2 and ZjCO8 were distributed on Chr6 and 10, respectively. We also compared the homologous genes between jujube and other related species, and the results showed that most identified flowering-related genes had high homology with those of other species (Table S2), indicating that conserved evolutionary pathways are shared among these plant species.

(In the circle diagram, the gene density is represented by two circles from inside to outside, and the outer circle shows the gene position on the chromosome.)
Organ-specific expression of the genes related to jujube flower bud differentiation
We investigated the expression patterns of 44 candidate genes in five different organs (Fig. 3), and the results showed that most of them were expressed in both the vegetative and reproductive organs. Compared to that in other organs, the expression of these genes in the roots was lower. In particular, the genes related to the photoperiod pathway were highly expressed in the stems, leaves, and flowers (Fig. 3). The genes showing high expression levels in the stems may be related to flower bud differentiation.

ZjACT was used as an internal control gene. Note: 1-photoperiod, 2-GA and flowering integration factor, 3-autonomy, 4-vernalization, 5-circadian clock and 6-ambient temperature
Photoperiod-related genes may play key roles in jujube flower bud differentiation
To further investigate the role of the above-mentioned genes in flower development, their expression patterns were determined throughout the flowering process using qRT-PCR. Based on the jujube flowering process (Fig. 1), ten typical periods were selected in this study. High expression levels of the genes related to the photoperiod pathway and two crucial genes (ZjFT and ZjSOC1) were observed during the early flower bud differentiation stages (Fig. 4), indicating that these genes may be key genes involved in jujube flowering. The trends of the expression patterns of most genes involved in vernalization and circadian rhythm pathways were opposite those of the genes involved in the photoperiod pathway, meaning that they should regulate the late flowering stages (Fig. 4). Thus, the above results preliminarily suggested that the genes related to the photoperiod pathway might play critical roles in jujube flower bud differentiation.

ZjACT was used as an internal control gene. The mean expression value was calculated from three independent biological replicates. Color bar: Log2(fold change). Note: A1. Unp (Undifferentiated period); A2. Ins (Initial stage); A3. Sed (Sepal differentiation); A4. Ped (Petal differentiation); A5. Std (Stamen differentiation); A6. Cad (Carpel differentiation); B1. Bue (Bud emergence); B2. Ine (Inflorescence emergence); B5. Ybs (Bud yellowing); C5. Pes (Petal flattening). The scaled log2 expression values are shown from blue to red
Photoperiod-related genes and ZjPIF4 positively respond to jujube flowering under protected cultivation
To verify the roles of the photoperiod pathway in jujube flowering, jujube trees in the greenhouse were used, and the flowering time was advanced to January from May in an open field. There are four types of shoots in jujube trees: primary shoots, secondary shoots, mother-bearing shoots, and bearing shoots. Mother-bearing shoots, which are extremely condensed (expanding by only approximately 1 mm a year), produce bearing shoots every year. Bearing shoots are the only flowering and fruiting shoots in Chinese jujube and drop in the winter. Thus, the expression of the above genes in mother-bearing shoots and their branch parts (bark) were further investigated before flowering.
Most photoperiod-related genes responded differently to the different developmental stages in the mother-bearing shoots and in their base branch bark (Fig. 5). Some of the genes (ZjICE1, ZjCO6, ZjSOC1, and ZjFT) in both tissues were expressed at high levels in the later developmental stages, suggesting that these genes may be involved in the flowering process. This experiment also indicated that the photoperiod pathway plays crucial roles in jujube flowering. Compared with that in the base branch bark, the expression of ZjPIF4 in the mother-bearing shoots was much higher in the early stages, indicating that ZjPIF4 is also an important regulator of floral initiation.

The mean expression value was calculated from three independent biological replicates. Note: 1. The first stage (October 28, 2018); 2. The second stage (January 8, 2019); 3. The third stage (January 18, 2019); 4. The fourth stage (January 28, 2019); 5. The fifth stage (February 28, 2019). The scaled log2 expression values are shown from blue to red
Photoperiod-related genes involved in the jujube phase change process
The seedlings of hybrid progeny of JMS2×Xing 16 with different flowering times were observed in two consecutive years. The flowering rates in the first and second years were 20% and 100%, respectively (Table S3). The individual seedlings were divided into juvenile, transition, and adult areas according to the secondary shoot and flowering positions (Fig. S1). The first flowering position of the hybrid progeny was concentrated in transitional secondary shoots, which is expected to become a marker trait of stage transformation.
We investigated the growth dynamics of the seedlings and the expression patterns of photoperiod-related genes during the phase transition (Fig. 6a). Most genes were highly expressed in the juvenile period, five genes (ZjICE1, ZjCO6, ZjCO4, ZjPHYC, and ZjCOP1) were highly expressed in the transition period, and four genes (ZjNFYC2, ZjNFYB5, ZjNFYB3, and ZjFT) were highly expressed in the adult period. Overall, the expression patterns of most photoperiod-related genes during the phase transition of seedlings were consistent with those in the perennial tree species (Fig. 4), suggesting that photoperiod-related genes are also involved in the phase transition.

a Note: 1. Juv (juvenile period); 2. Trp (transition period); 3. Adp (adult period). The scaled log2 expression values are shown from blue to red. b Relative expression of the photoperiod genes under long-day and short-day treatments. Note: 1. Control (12 h light/12 h dark); 2. The long-day treatment: 14 h light/10 h dark; 3. The long-day treatment: 16 h light/8 h dark; 3. The long-day treatment: 18 h light/6 h dark; 5. The short-day treatment: 10 h light/14 h dark; 6. The short-day treatment: 8 h light/16 h dark; 7. The short-day treatment: 6 h light/18 h dark. The scaled log2 expression values are shown from blue to red. The mean expression values were calculated from three independent biological replicates
Photoperiod-related genes positively respond to photoperiod regulation
To further verify the photoperiod-related genes involved in jujube flowering, the jujube seedlings were subjected to long-day and short-day photoperiod treatments under the same growth conditions. We found that 68% and 58% of the genes responded to the long-day and short-day treatments, respectively (Fig. 6b). In the long-day treatment, the expression of these genes (ZjNFYB5, ZjNFYC3, ZjNFYB3, ZjCO7, ZjCO4, ZjCO5, ZjCO6, ZjNFYC1, ZjCRY1, ZjAS1, ZjPHYA, ZjCO2, ZjCO3, ZjNFYC2, and ZjCOP1) increased significantly with increasing light duration, and the highest expression was found under an 18 h/6 h light/dark photoperiod. In the short-day treatment, the expression of these genes (ZjNFYB5, ZjCRY1, ZjAS1, ZjPHYA, ZjCO3, ZjNFYC2, ZjCO1, ZjNFYA1, ZjNFYC9, and ZjGI) increased significantly and peaked under a 6 h/18 h light/dark photoperiod. The photoperiod regulation significantly altered the expression of photoperiod-related genes, and the long-day treatment also increased the early flowering of jujube seedlings (Table S4).
Protein–protein interactions among several key flower-related proteins
Based on the orthologs of Arabidopsis, the interactions of photoperiod-related proteins of Chinese jujube were predicted by STRING analysis (Fig. 7a). The results predicted that many candidate key proteins interact with each other, i.e., PHYB (homolog of ZjPHYB) could interact with PIF4 (homolog of ZjPIF4), CRY1 (homolog of ZjCRY1) could interact with COP1 (homolog of ZjCOP1), COP1 (homolog of ZjCOP1) could also interact with GI (homolog of ZjGI) and CO (homolog of ZjCO1), BBX19 (homolog of ZjCO5) could interact with CO (homolog of ZjCO1) and COP1 (homolog of ZjCOP1), and FT (homolog of ZjFT) could interact with CO (homolog of ZjCO1) and AGL20 (homolog of ZjSOC1). A yeast two-hybrid assay (Y2H) then verified that there were interactions between ZjCO5 and ZjFT and between ZjCO5 and ZjAS1 (Fig. 7b). Furthermore, ZjPIF4, an important regulator related to photoperiod and temperature, interacted directly with ZjCO5 and ZjFT (Fig. 7b). Take together, these results indicated that the photoperiod- and ambient temperature-related pathways are the key pathways involved in jujube flowering and that the ZjPHY family members ZjPIF4, ZjFT and ZjCO5 are the key factors involved in the regulatory network.

a Protein–protein interaction analysis of ZjPHYB, ZjGI, ZjCO and ZjFT by STRING. b Yeast two-hybrid screening of select proteins involved in the photoperiod-related and ambient temperature-related pathways
Discussion
The distinct flowering process of Chinese jujube
Flowering is an intricate developmental process involving several stages, including floral induction, floral initiation, flower development, and blooming7. Here, we found that flower bud initiation and differentiation lasted less than 1 month in Dongzao, the most popular jujube cultivar for fresh markets5, and showed a fast-slow-fast trend. However, in other fruit tree species, such as apple, pear, and peach, flower bud initiation starts with the cessation of shoot growth7,8, and floral differentiation occurs in the next year9,10. Thus, flower bud initiation and differentiation occur in two consecutive years in most fruit trees11. As a perennial woody fruit tree species, Chinese jujube has a very fast differentiation rate during flowering, which is a distinct trait of fruit trees, but its molecular mechanisms need to be further studied. Additionally, apart from the 44 flowering-related genes identified, additional genes may be involved in the complex flowering process in the jujube genome.
The photoperiod pathway is the dominant pathway in Chinese jujube flowering
Many plants sense seasonal cues, day length or photoperiod changes to align the timing of the developmental transition to flowering with the changing seasons for reproductive success12. In this study, the expression levels of most photoperiod-related genes in jujube were responsive to flower bud differentiation, which was confirmed using samples with phase transitions and those under protected cultivation. The transition from the juvenile vegetative phase to the adult vegetative phase is referred to as the juvenile-to-adult transition or vegetative phase change13. This transition is an important shift for the acquisition of adult vegetative characteristics and subsequent reproductive competence14. Most photoperiod-related genes were highly expressed in the juvenile and transition phases, indicating that their function contributed to the juvenile-to-adult transition.
In general, the circadian rhythm pathway is associated with the photoperiod pathway, and endogenous circadian rhythms are sensitive to light/dark stimuli15,16. The expression patterns of most genes involved in these two pathways showed sequential expression in this study (Fig. 4). Overall, the expression of the genes related to the photoperiod pathway occurred earlier than did that of the genes related to the circadian rhythm pathway, indicating that the photoperiod pathway might play a pivotal role during flower bud differentiation and that the circadian rhythm pathway may play an important role in flower development. Compared to annual herbs, the growth process of perennial fruit trees is more complex, and the environmental factors are more uncontrollable; thus, their molecular mechanisms of flowering might have distinct characteristics and need to be further studied.
ZjPIF4 is an orthologous gene of PIF4, which is a core component of the photoperiod- and ambient temperature-related pathways17,18,19. Under protected cultivation, the expression of ZjPIF4 was also high in early developmental stages, and Y2H assays proved that ZjPIF4 could also interact with the photoperiod-related proteins ZjCO5 and ZjFT (Fig. 7b), indicating that crosstalk occurred between the photoperiod- and ambient temperature-related pathways in jujube. Moreover, the expression of an increasing number of photoperiod-related genes increased significantly under long-day conditions, and the jujube seedlings showed early flowering (Fig. 6b, Table S4). Overall, all the above results indicated that the photoperiod pathway is the dominant pathway in Chinese jujube (Fig. S2).
Roles of the photoreceptors ZjPHYs and ZjCRY1 in Chinese jujube
Light is necessary for plant development, and plants deploy sensory photoreceptors such as phytochromes to capture light energy20. After capturing an incoming photon, activated phytochrome molecules must relay the information to nuclear genes that are poised to respond by directing appropriate adjustments in plant development21. Red/far-red/blue light sensing in higher plants is mediated by a diverse but structurally conserved group of soluble photoreceptors. In Arabidopsis, the phytochrome family consists of five members designated phyA to phyE22. In the present study, three phytochrome genes, ZjPHYA, ZjPHYB, and ZjPHYC, were identified in the jujube genome, and the expression of ZjPHYA, which mediates far-red-light stimulation, was higher during the vegetative growth stage than during the reproductive stage but gradually decreased during reproductive growth (Figs. 5 and 6a). ZjPHYA and ZjPHYC were also responsive to long- and short-day conditions (Fig. 6b). ZjCRY1, which encodes a blue light receptor, was also highly expressed in the juvenile period and in the early stage under protected cultivation (Figs. 5 and 6a).
Roles of ZjPIF4 and ZjFT in Chinese jujube
PHYTOCHROME INTERACTING FACTOR 4 (PIF4), a core component of high-temperature signaling18,19,23,24,25, was identified as a molecular link connecting high-temperature signaling and stomatal development and revealed a direct mechanism by which the production of a specific cell lineage can be controlled by a broadly applicable environmental signaling factor23. The expression patterns of ZjPIF4 were investigated under different conditions in an open field and under protected cultivation. In the open field, the expression of ZjPIF4 increased at the later stage of flower development, and the temperature also increased during this period (in May). Under protected cultivation, ZjPIF4 was highly expressed in early stages, in which the temperature ranged from 20 to 23 °C. Thus, the results under different conditions were consistent with each other, suggesting that ZjPIF4 is responsive to increasing temperature. Previous studies have shown that high temperature induces the expression of PIF4 at both the transcriptional and posttranscriptional levels18,24,25, and many PIF family transcription factors are well-established repressors of light signaling19,26,27,28 and interact with light-activated phytochromes23,29,30,31. In jujube, ZjPIF4 interacted with the photoperiod-related protein ZjCO5 (Fig. 7b), which is consistent with the results of previous studies.
An unregulated photoperiod induces the expression of FLOWERING LOCUS T (FT) in the phloem companion cells (PCC) of leaves. The FT protein then acts as a florigen that is translocated to the shoot apical meristem (SAM)32,33,34,35,36. Based on the jujube genome annotation, there is only one FT gene in this species. ZjFT shared high identity with homologous genes in Rosacea species, e.g., 96.84% identity with MdFT (Malus × domestica, ACV92037.1), 93.68% identity with PbFT (Pyrus × bretschneideri, NP_001289254.1) and 91.95% identity with PpFT (Prunus persica, XP_007206002.1). Conserved domain analysis showed that ZjFT contains a phosphatidyl ethanolamine-binding protein (PEBP) domain, which is the conserved domain of FT. Taken together, all of these results showed that ZjFT is a homologous gene of FT.
As a key transcription factor in the bHLH family, PIF4 directly binds to the promoter region of FT and may also regulate FT at the protein level in Arabidopsis37. In the present study, ZjPIF4 was proven to interact with ZjFT at the protein level, which is consistent with the results of a previous study. Moreover, in Arabidopsis, PIF4 also interacts with DELLA proteins to integrate photoperiod–GA–circadian clock signals38,39. Thus, PIF4 likely has multiple functions at the transcriptional and translational levels, and its homologous genes in jujube and other fruit tree species are worthy of further research.
As shown in Figs. 4 and 5, the expression patterns of ZjPIF4 and ZjFT in the field and under protected cultivation were not different. In actuality, the growth period of the samples collected in protected cultivation (Fig. 5) occurred earlier than did that in the field (Figs. 1 and 4), and the later growth stages under protected cultivation were occurring just when the initial stages were occurring in the field. Thus, the expression patterns of ZjPIF4 and ZjFT at later stages in protected cultivation should be consistent with those at initial stages in the field, which was confirmed in this study.
ZjFT was highly expressed in bearing shoots before flowering in jujube (Fig. 5), and the expression pattern was similar to that of Arabidopsis3; thus this gene likely plays an important role in the transformation from vegetative growth to reproductive growth in Chinese jujube. However, the expression of ZjFT and ZjPIF4 was higher on short days than on long days, and the jujube seedlings showed relatively early flowering under long-day conditions (Fig. 6b). Indeed, some related studies in Pharbitis40 and Vitis vinifera L.41 also showed similar results. In Pharbitis, the expression of PnFT increased when the night was longer than 11 h40. In Vitis vinifera L., shading treatment was beneficial for enhancing the expression of VvPIF441. Thus, the expression of ZjFT and ZjPIF4 in jujube might also be induced by night treatment under unique conditions or at specific developmental stages, and the detailed regulatory mechanism needs to be further elucidated.
Material and methods
Plant materials
Ten-year-old jujube (Ziziphus jujuba Mill. “Dongzao”) trees were grown at the experimental station of Hebei Agricultural University (HAU), Baoding, China. Roots, stems, leaves, flowers and young fruits were collected from three trees at 9:00 a.m. to 10:00 a.m. Bearing shoots from 0.5 mm to 20 mm were collected during flower bud differentiation, flower buds were harvested at the flower development stage, and flowers were collected while they were blooming.
Mother-bearing shoots and their phloem were harvested from ten-year-old Dongzao jujube trees under protected cultivation. The jujube trees were cultivated in a greenhouse, in which the indoor temperature was controlled at approximately 7–10 °C during the dormant period (approximately 20 days) and then was increased by 2–3 °C every week until the jujube trees sprouted. The temperature was maintained at 25–27 °C during the growth, flowering and fruiting periods, and the humidity remained at 60%–70%.
Jujube seedlings were sown in a laboratory with constant temperature and humidity, which were maintained at 23 °C and 70%, respectively. The plants were adapted to a 12 h light/12 h dark photoperiod for two weeks before treatment, with lights-on defined as zeitgeber time (ZT0) and lights-off defined as ZT12; ZT2 represents 2 h after the start of the light period. The seedlings were treated with long-day (14 h/10 h, 16 h/8 h, 18 h/6 h) and short-day (10 h/14 h, 8 h/16 h, 6 h/18 h) photoperiod treatments. A 12 h/12 h photoperiod treatment was used as the control. At 60 days after treatment, 15 seedlings were sampled in each treatment, and three biological replicates (5 seedlings mixed in one replicate) were collected at ZT2. Their flowering process was observed, and samples were stored at −80 °C for RNA extraction and expression analysis.
Observation of the flowering process
When the perennial jujube trees began to sprout, we used Vernier calipers to measure dynamically and collect samples from the mother-bearing shoots at 9:00 a.m. (from April 5 to 18, 2019). The bearing shoots were subsequently cut into 0.5 mm, 1.0 mm, and 1.2 mm to 20 mm pieces (increments of 0.2 mm units) and then fixed with FAA solution. We then used paraffin to make permanent slices of the shoots42. After sealing solidly, the permanent air-dried slices were placed under a Zeiss light microscope to observe the morphological changes during flower bud development and were imaged by a ZEN universal imaging system.
To observe the process of flower development, we collected flower buds from the bud emergence stage to the blooming stage (from May 17 to June 22, 2019). According to the characteristics of flower development, we collected samples from 9:00 a.m to 10:00 a.m. The flower buds ranged from 500 to 2500 μm in size. We recorded the time and observed the morphological changes associated with flower opening under an XTJ-4400 stereomicroscope. CAM-MA imaging software was used to image the samples.
Jujube seedlings used for phase transition
The tested materials were sown at the experimental station of HAU in 2018. Seedlings of JMS2×Xing16 offspring were used; there was a total of 68 plants under conventional management. Plant height and width were measured by a ruler, the base diameter and the initial diameter were measured by a vernier caliper, and the first flowering node and position were recorded. Bearing shoots were collected from jujube seedlings at 9:00 a.m. (on August 2018).
Identification of flowering pathway genes
Based on the Arabidopsis flower network, the NCBI (http://www.ncbi.nlm.nih.gov/) and PFAM databases (http://pfam.sanger.ac.uk) were first used to identify the genes of various flowering pathways in the jujube genome. Homologous alignment was performed using the BLAST tool of NCBI to eliminate genes with inaccurate annotations. Moreover, a model (HMMER 3.0) was used to further search for all the flowering genes in the protein library43. The online tool ProtParam (http://web.expasy.org/protparam/) was used to predict the molecular weight (MW) and isoelectric point (pI) of each protein.
Chromosomal location and gene duplication
Each identified gene was mapped to pseudochromosomes according to its coordinates on the jujube genome. Tandem duplications were investigated according to previous methods44. A circle graph was ultimately generated by TBtools (http://cj-chen.github.io/tbtools/).
RNA isolation
According to the manufacturer’s instructions, total RNA was isolated from 200 mg of various tissues of jujube using an RNA Prep Pure Plant Kit (Tiangen, China). Using a TIAN Script First Strand cDNA Synthesis Kit (Tiangen, China), first-strand cDNA was synthesized from 2 μg of total RNA. The cDNA obtained was then diluted and stored at −20 °C for subsequent expression analyses.
Gene expression analyses
Primers for semiquantitative RT-PCR and qRT-PCR were designed using Primer Premier 5.0, and expression patterns were assayed by semiquantitative RT-PCR. PCR was performed using the following program: initial denaturation at 95 °C for 3 min; 30 cycles at 95 °C for 15 s, 55 °C for 15 s, and 72 °C for 30 s; and a final extension cycle at 72 °C for 10 min6. The primers used in this study are listed in Table S5.
The expression of flowering-related genes at different developmental periods was examined using qRT-PCR (Bio-Rad iQ5). According to the instructions (Trans Start Top Green qPCR SuperMix), all reactions were performed in a 20 μL system consisting of 1 μL of cDNA, 10 μL of SYBR Green mix and each primer at 400 nM. The procedures were as follows: 94 °C for 30 s followed by 40 cycles of 94 °C for 5 s, 55 °C for 15 s and 72 °C for 15 s. All experiments were performed for three biological replicates, and each replicate was measured in triplicate. The specificity of the amplicon for each primer pair was verified by using melting curve analysis. ZjACT was used as a reference gene, and the relative expression levels of the tested genes were calculated by using the 2−ΔΔCt method6,45,46.
Prediction of protein–protein interactions
The interactions among photoperiod-related proteins in jujube were predicted by STRING (https://string-db.org/cgi/input.pl). Their orthologs in Arabidopsis were used as references.
Yeast two-hybrid (Y2H) assays
Tested proteins were fused to the Gal4 DNA-binding domain (BD), and the screening proteins were fused to the Gal4 activation domain (AD). The AD-fused proteins and BD-fused proteins were amplified using the primers listed in Table S6 and then cloned into the pGADT7 and pGBKT7 vectors, respectively. The vectors were subsequently cotransformed into yeast strain AH109. NdeI-SmaI was used to digest the tested protein, and EcoRI was used to digest the candidate screening protein. The freshly transformed yeast colonies were resuspended in 10 μL of sterile deionized water, and 0.5 μL aliquots were spotted onto four selective media: synthetically defined (SD) media lacking leucine and tryptophan (−LW); media lacking leucine, tryptophan and histidine (−LWH); −LWH media supplemented with 7 mM 3-amino-1,2,4-triazole (3-AT, Sigma Aldrich) (−LWH+3AT); and media lacking leucine, tryptophan, adenine, and histidine (−LWAH). The growth conditions of the above yeast colonies were monitored after 3 days of incubation at 28 °C47.
References
Hayama, R. & Coupland, G. Shedding light on the circadian clock and the photoperiodic control of flowering. Curr. Opin. Plant Biol.6, 13–19 (2003).
Andrés, F. & Coupland, G. The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet.13, 627–639 (2012).
Blümel, M., Dally, N. & Jung, C. Flowering time regulation in crops-what did we learn from Arabidopsis? Curr. Opin. Biotechnol.32, 121–129 (2015).
Zhai, S. B., Su, L. C., Fang, Y. N., Li, L. & Guo, D. L. Effects of KClO3 on the expression of CONSTANS family genes in the control of flower bud formation of longan. Plant Physiol. J.52, 1429–1437 (2016).
Liu, M. J. et al. The complex jujube genome provides insights into fruit tree biology. Nat. Commun.5, 5315 (2014).
Zhang, L. et al. Genome-wide identification, characterization of the MADS-box gene family in Chinese jujube and their involvement in flower development. Sci. Rep.7, 1025 (2017).
Kotoda, N. et al. Expression pattern of homologues of floral meristem identity genes LFY and AP1 during flower development in apple. J. Am. Soc. Hortic. Sci.125, 398–403 (2000).
Hanke, M. H., Flachowsky, H., Peil, A. & Hättasch, C. No flower no fruit-genetic potentials to trigger flowering in fruit trees. Glob. Sci. Books Genes Genomes Genomics.1, 1–20 (2007).
Zhu, L. H., Borsboom, O. & Tromp, J. The effect of temperature on flower-bud formation in apple including some morphological aspects. Scientia Horticul.70, 1–8 (1997).
Li, Y. et al. Transcriptomic analysis reveals the regulatory module of apple (Malus×domestica) floral transition in response to 6-BA. BMC Plant Biol.19, 1 (2019).
Ito, A., Hayama, H., Kashimura, Y. & Yoshioka, H. Effect of maleic hydrazide on endogenous cytokinin contents in lateral buds, and its possible role in flower bud formation on the Japanese pear shoot. Sci. Horticul.87, 199–205 (2001).
Wellmer, F. & Riechmann, J. L. Gene networks controlling the initiation of flower development. Trends Genet.26, 519–527 (2010).
Guo, C. et al. Repression of miR156 by miR159 regulates the timing of the juvenile-to-adult transition in Arabidopsis. Plant Cell.29, 1293–1304 (2017).
Hayama, R. & Coupland, G. The molecular basis of diversity in the photoperiodic flowering responses of Arabidopsis and rice. Plant Physiol.135, 677–684 (2004).
Alabadí, D. et al. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science293, 880–883 (2001).
Wang, Y. et al. Light-and temperature-entrainable circadian clock in soybean development. Plant Cell Environ.43, 637–648 (2020).
Koini, M. A. et al. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol.19, 408–413 (2009).
Kumar, S. V., Lucyshyn, D., Jaeger, K. E., Alós, E. & Alvey, E. Transcription factor PIF4 controls the thermosensory activation of flowering. Nature484, 242–245 (2012).
Sun, J. Q. et al. PIF4 and PIF5 transcription factors link blue light and auxin to regulate the phototropic response in Arabidopsis. Plant Cell.25, 2102–2114 (2013).
de Wit, M., Galvão, V. C. & Fankhauser, C. Light-mediated hormonal regulation of plant growth and development. Annu. Rev. Plant Biol.29, 513–537 (2016).
Martina, L., Yetkin, C. L. & Christian, F. Molecular mechanisms underlying phytochrome controlled morphogenesis in plants. Nat. Commun.10, 5219 (2019).
Mathews, S. & Sharrock, R. A. Phytochrome gene diversity. Plant Cell Environ.20, 666–671 (1997).
Lau, O. S. et al. Direct control of SPEECHLESS by PIF4 in the high-temperature response of stomatal development. Curr. Biol.: Cb.28, 1273 (2018).
Leivar, P. et al. Dynamic antagonism between phytochromes and PIF family basic helix-loop-helix factors induces selective reciprocal responses to light and shade in a rapidly responsive transcriptional network in Arabidopsis. Plant Cell24, 1398–1419 (2012).
Foreman, J. et al. Light receptor action is critical for maintaining plant biomass at warm ambient temperatures: Light pathways buffer the harmful effects of heat. Plant J.65, 441–452 (2011).
Hayes, S. PIF4 plays a conserved role in Solanum lycopersicum. Plant Physiol.181, 838–839 (2019).
Galvão, V. C. et al. PIF transcription factors link a neighbor threat cue to accelerated reproduction in Arabidopsis. Nat. Commun.10, 4005 (2018).
Sakuraba, Y. et al. Phytochrome-interacting transcription factors PIF4 and PIF5 induce leaf senescence in Arabidopsis. Nat. Commun.5, 4636 (2014).
Leivar, P. & Quail, P. H. PIFs: pivotal components in a cellular signaling hub. Trends Plant Sci.16, 19–28 (2011).
Al-Sady, B. et al. Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Mol. Cell.23, 439–446 (2006).
Rosado, D. et al. Downregulation of PHYTOCHROME- INTERACTING FACTOR 4 influences plant development and fruit production. Plant Physiol.181, 1360–1370 (2019).
Collani, S., Neumann, M., Yant, L. & Schmid, M. FT Modulates genome-wide DNA-binding of the bZIP transcription factor FD. Plant Physiol.180, 367–380 (2019).
Serrano-Bueno, G. et al. CONSTANS-FKBP12 interaction contributes to modulate photoperiodic flowering in Arabidopsis. Plant J. https://doi.org/10.1111/tpj.14590 (2019).
Song, G. Q. et al. VcFT-induced mobile florigenic signals in transgenic and transgrafted blueberries. Horticulture Res.6, 105 (2019).
Putterill, J. & Varkonyi-Gasic, E. FT and florigen long-distance flowering control in plants. Curr. Opin. Plant Biol.33, 77–82 (2016).
Notaguchi, M. et al. Long-distance, graft-transmissible action of Arabidopsis FLOWERING LOCUS T protein to promote flowering. Plant Cell Physiol.49, 1645–1658 (2008).
Halliday, K. J., Salter, M. G., Thingnaes, E. & Whitelam, G. C. Phytochrome control of flowering is temperature sensitive and correlates with expression of the floral integrator FT. Plant J.484, 875–885 (2003).
Jing, Y., Guo, Q., Zha, P. & Lin, R. The chromatin-remodelling factor PICKLE interacts with CONSTANS to promote flowering in Arabidopsis. Plant, Cell Environ.42, 2495–2507 (2019).
Feng, S. et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature451, 475–479 (2008).
Hayama, R., Agashe, B., Luley, E. & Coupland, K. G. A circadian rhythm set by dusk determines the expression of ft homologs and the short-day photoperiodic flowering response in pharbitis. Plant Cell19, 2988–3000 (2007).
Zhan, K. K. et al. Genome-wide identification of PIFs in grapes (Vitis vinifera L.) and their transcriptional analysis under Lighting/Shading Conditions. Genes9, 451 (2018).
Li, J. X. et al. Identification of genes associated with lemon floral transition and flower development during floral inductive water deficits: A hypothetical model. Front. Plant Sci.8, 1013 (2017).
Finn, R. D., Clements, J. & Eddy, S. R. HMMER web server: interactive sequence similarity searching. Nucleic Acids Res.39, W29–W37 (2011).
Kong, A. Y. Y. et al. Soil aggregates control N cycling efficiency in long-term conventional and alternative cropping systems. Nutrient Cycl. Agroecosystems.79, 45–58 (2007).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 -ΔΔCT method. Methods25, 402–408 (2001).
Bu, J. D., Zhao, J. & Liu, M. J. Expression stabilities of candidate reference genes for RT-qPCR in Chinese jujube (Ziziphus jujuba Mill.) under a variety of conditions. PLoS ONE11, 1–11 (2016).
Xue, C. L. et al. Genome-wide analysis of the WRKY gene family and their positive responses to phytoplasma invasion in Chinese jujube. BMC Genomics.20, 1–14 (2019).
Acknowledgements
This work was supported by the National Key R&D Program Project Funding (2018YFD1000607), the National Natural Science Foundation of China (31772285), the Significance Fund of the Hebei Province Natural Science Foundation (C2017204114), the Foundation for 100 Innovative Talents of Hebei Province (SLRC2019031), and the Hebei Special Funds for Intellectual Introduction and Talent Training (A2016002054). These funding bodies had no role in the design of the research, sample collection, data analysis or writing of the manuscript.
Author information
Authors and Affiliations
Contributions
J.Z. and M.L. designed the experiments. X.M., Y.L., and J.Z. performed the main experiments, analyzed the results and wrote the manuscript. Y.Y., Y.Z., and H.L. participated in the data analysis. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Meng, X., Li, Y., Yuan, Y. et al. The regulatory pathways of distinct flowering characteristics in Chinese jujube. Hortic Res 7, 123 (2020). https://doi.org/10.1038/s41438-020-00344-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41438-020-00344-7

{kind=link}
{kind=link}