
Abstract
The RASopathies are a group of clinically and genetically heterogeneous developmental disorders caused by dysregulation of the RAS/MAPK signalling pathway. Variants in several components and regulators of this pathway have been identified as the pathogenetic cause. In 2015, missense variants in A2ML1 were reported in three unrelated families with clinical diagnosis of Noonan syndrome (NS) and a zebrafish model was presented showing heart and craniofacial defects similar to those caused by a NS-associated Shp2 variant. However, a causal role of A2ML1 variants in NS has not been confirmed since. Herein, we report on 15 individuals who underwent screening of RASopathy-associated genes and were found to carry rare variants in A2ML1, including variants previously proposed to be causative for NS. In cases where parental DNA was available, the respective A2ML1 variant was found to be inherited from an unaffected parent. Seven index patients carrying an A2ML1 variant presented with an alternate disease-causing genetic aberration. These findings underscore that current evidence is insufficient to support a causal relation between variants in A2ML1 and NS, questioning the inclusion of A2ML1 screening in diagnostic RASopathy testing.
Similar content being viewed by others

Introduction
The RASopathies comprise a group of syndromes sharing craniofacial features, cardiac anomalies, short stature, and variable developmental delay [1]. Noonan syndrome (NS; OMIM 163950) is the most common disorder among them. The molecular basis underlying NS is dysregulation of the RAS/MAPK pathway leading to increased ERK activation. Accordingly, the demonstration of an increased phospho-ERK/ERK ratio basally and/or after receptor tyrosine kinase (RTK) stimulation is a well-established functional assay for NS-associated mutants [2]. Variants in over 15 genes encoding components and direct modulators of the RAS/MAPK-pathway have been associated with NS, and further heterogeneity is anticipated [3]. NS-associated variants act in a dominant manner, with the exception of LZTR1, for which dominant and recessive inheritance is observed [4]. In 2015, Vissers et al. reported the association of heterozygous germline variants in A2ML1 with a disorder clinically related to NS [5]. Two subsequent publications reported three additional A2ML1 variants in NS patients [6, 7]. A2ML1 encodes alpha-2-macroglobulin-like 1, a protein of the α2-macroglobulin protease inhibitor superfamily, which acts as a protease inhibitor and component of the complement system [8, 9]. It is most abundantly expressed in keratinocytes, but also found in placenta, thymus and testis [9]. Its physiological role is largely unknown and no effect on the RAS/MAPK pathway has been demonstrated so far [10]. Genomic A2ML1 variants are also reported in different clinical context, namely as genetic factor conferring susceptibility to otitis media [11, 12]. Furthermore, autoantibodies against A2ML1 are frequently detected in individuals with paraneoplastic pemphigus [13, 14]. Herein, we report our experience with A2ML1 mutational screening in 1029 patients with clinical suspicion of NS or another RASopathy.
Patients and methods
Subjects
Two prospective cohorts of 369 (University Hospital Magdeburg, Germany) and 470 (San Giovanni Rotondo, Italy) patients with clinical signs of NS/RASopathy underwent multigene panel sequencing of RASopathy-related genes including A2ML1 in a diagnostic setting. Two additional cohorts including 95 patients each (Rome, Italy; Paris, France) with clinical features suggestive of NS/RASopathy were retrospectively screened for A2ML1 variants in the frame of research projects. Informed consent was obtained from individuals or their legal guardians for genetic testing according to national legal regulations and for the anonymous publication of genotype and phenotype data. Individuals for which phenotype data are reported here were assessed by experienced clinical geneticists (Supplementary Methods).
Molecular analysis and variant classification
Genomic DNA was extracted from blood or saliva samples, according to standard procedures. Multigene panel sequencing was performed using different techniques according to standard protocols (for further details see Supplementary Methods). Standard and RASopathy-adapted ACMG-criteria were applied to classify identified and previously published variants [2, 15]. Accession version numbers on which variant descriptions are based are given in Supplementary Table 1.
Results
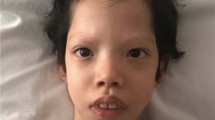
We identified 15 individuals with rare and novel non-synonymous variants in A2ML1 (Supplementary Table 1). In ten cases, variants were inherited from a clinically unaffected parent (case 3, 5, 6, 8, 9, 10, 12, 13, 14, 15). Seven individuals were found to have a concurrent known (likely) causal genetic variant explaining the phenotype: six (case 2, 3, 4, 7, 8, 13) had variants in established RASopathy genes (Supplementary Table 1) and one turned out to have a microdeletion 7q11.2 confirming clinical diagnosis of Williams-Beuren syndrome (case 12). In patient 3, we detected a known HRAS variant, reported to cause a mild Costello syndrome-phenotype [16], together with an A2ML1 variant, c.1769C>T, p.(Ala590Val), located closely to the site of the reported missense variant p.(Arg592Leu) [5]. The HRAS variant was inherited from her similarly affected mother while the A2ML1 variant was inherited from the unaffected father. Subject 7 harboured a known NS-causing RAF1 variant, c.770C>T, p.(Ser257Leu) [17], and subject 8 (previously published [18]) harboured a likely causal de novo variant in MAP2K1 c.275T>G, p.(Leu92Arg) consistent with a clinical phenotype of cardio-facio-cutaneous (CFC) syndrome [2]. Both were also carrying the same A2ML1 variant p.(Arg802His) that had been previously reported as a de novo variant in the index family reported by Vissers et al. [5]. For subject 8, the A2ML1 variant was inherited from the unaffected father. The clinical presentation of patients with concurrent known RASopathy-causing variants was fitting with the expected phenotype and no additional or extraordinarily severe symptoms were exhibited that could not be accounted by their respective variant in CBL, HRAS, RIT1, RAF1, MAP2K1 and PTPN11.
In one of the eight remaining individuals (case 1) without an alternate variant in an established gene for NS, parental DNAs were not available to test segregation, but careful re-analysis of the phenotype indicated that clinical features were insufficient to meet the criteria for a definite clinical diagnosis of NS [19]. The A2ML1 variant observed in this case was formally classified as VUS. Subject 11 inherited the A2ML1 variant from her father who was also considered as possibly affected by NS, but this variant was classified as LBEN/BEN, because of its population frequency. A summary of all variants from this study as well as variants previously reported in the literature and their classification is shown in Supplementary Table 1. All identified variants were submitted to ClinVar (www.ncbi.nlm.nih.gov/clinvar/ under the submission number SUB7750339. Accession numbers are shown in Supplementary Table 1).
Discussion
Three previous reports associated A2ML1 variants with NS/NS-like-phenotypes [5,6,7]. Vissers et al. identified a de novo c.2405G>A, p.(Arg802His) variant by trio exome sequencing in a girl with craniofacial and thoracic anomalies suggestive of NS, intellectual disability, anxiety disorder, easy bruising, failure to thrive in infancy, prenatally detected hygroma colli and polyhydramnios. Two additional variants, c.2405G>T, p.(Arg802Leu) and c.1775G>T, p.(Arg592Leu), were detected by targeted sequencing in a cohort of 155 individuals with a clinical suspicion of NS. Both variants were familial, and carriers displayed variable symptoms mostly belonging to the various non-specific anomalies that may occur in NS (pigmented nevi, anxiety disorder, easy bruising, intellectual disability, prenatally detected polyhydramnios or failure to thrive). Only one out of seven A2ML1 variant carriers had a heart defect typical of NS (pulmonary stenosis) and only two had short stature. One carrier was completely asymptomatic [5]. Several A2ML1 variant carriers from these families were classified as having facial features of NS. However, after critical review of the published facial photographs, the presence of a “suggestive face” or “NS-like-features” in cases 3, 5 and 7 from this publication is disputable [5], although it has to be admitted that there is a considerable subjective bias judging the phenotype and evaluation of photographs may not be equal to personal examination. We therefore argue that with strict application of published diagnostic criteria [19] hardly anyone of the families 2 and 3 in the paper by Vissers et al. would meet the diagnosis of NS. Absence of a clear NS phenotype in two A2ML1 variant carriers was also reported by Leung et al. [7]. Van Trier et al. reported the variant c.4061+1G>A in one patient from a cohort of 97 individuals with NS, who were examined for external ear abnormalities and hearing impairment but no phenotypic details were provided [6]. Consistently, application of diagnostic criteria for NS [19] revealed that the majority of A2ML1 variant carriers from our cohort, who had no alternate variant in an established RASopathy gene, did not meet the criteria for a definite diagnosis of NS (Supplementary Table 1).
Vissers et al. could not detect significant RAS/MAPK pathway activation for transfected A2ML1 mutant HEK-293T and COS7 cells. Expression of the mutants in zebrafish, however, was observed to lead to craniofacial and cardiac defects similar to those induced by a NS-causing Shp2 mutant, but abnormalities were milder and appeared later. Leung et al. reported in vitro and in vivo functional assessment of the variants c.256C>T, p.(Pro86Ser) and c.4261C>T, p.(Gln1421*) and could not show activation of the RAS/MAPK pathway, nor abnormal development of zebrafish embryos [7]. In the frame of this study, no functional studies where performed that could either support or disprove the data published by Vissers et al. [5]. A2ML1 is a large gene with considerable variation in the normal population. Analysis of constraint metrics of population databases showed that the probability of loss of function intolerance for A2ML1 is very low (ExAC/gnomAD pLI 0.00, gnomAD observed/expected ratio 0.77). Vissers et al. also reported further 36 non-pathogenic A2ML1 variants from a total of 295 cases (Supplementary Information) [5], adding to this perception. Taken together, the previously published evidence regarding association of A2ML1 with NS has left several points of concern: (i) failure to demonstrate an impact of A2ML1 variants on RAS/MAPK activation by well-established assays, (ii) presence of reported NS-associated A2ML1 variants in the general population, (iii) incomplete segregation of a definite NS phenotype with familial A2ML1 variants, and (iv) lack of plausible concept for effect of mutant A2ML1 proteins (gain of function, loss of function, dominant negative). This was also noted in a recent article reviewing gene-disease associations for RASopathies [3].
The present study provides further evidence against a driver role of A2ML1 in RASopathies. In more than 1000 individuals that underwent A2ML1 sequencing, we did not observe any variant in A2ML1 that could be considered as pathogenic or likely pathogenic for NS. Instead, inheritance from an unaffected parent in cases where samples could be obtained (10/15) and a high rate of concurrence with other causal aberrations fully explaining the phenotype (7/15) argue against pathogenicity for most identified A2ML1 variants. Specifically, we demonstrate concurrence with a NS causing RAF1 variant and a likely pathogenic de novo MAP2K1 variant as well as inheritance from an unaffected parent for the previously reported variant p.(Arg802His) [5]. Re-assessment of previously reported A2ML1 variants also demonstrated that three out of six had a population frequency that is far greater than expected for NS (Supplementary Table 1) [2].
In conclusion, our findings genuinely challenge the role of A2ML1 variants as cause of NS. It has been argued that incomplete penetrance might explain the observed lack of segregation [5], which is difficult to exclude definitely. However, concurrence of A2ML1 variants with clearly disease-causing variants in alternate genes in individuals affected by NS, and particularly the fact that this was observed for the same variant that was originally reported as a de novo NS-associated variant, leave little room for a NS-causing role of those A2ML1 variants. We do not exclude that variants in A2ML1 may cause any other human phenotype, but at least for the variants observed in this study, the presented findings do not support a dominant pathogenic effect. Based on this evidence, we conclude that screening of A2ML1 in the diagnostic setting is questionable, and particularly advice against A2ML1 testing in prenatal settings, as there is imminent risk of misjudgement.
References
Tartaglia M, Gelb BD, Zenker M. Noonan syndrome and clinically related disorders. Best Pr Res Clin Endocrinol Metab. 2011;25:161–79.
Gelb BD, Cavé H, Dillon MW, Gripp KW, Lee JA, Mason-Suares H, et al. ClinGen’s RASopathy Expert Panel consensus methods for variant interpretation. Genet Med. 2018;20:1334–45.
Grant AR, Cushman BJ, Cavé H, Dillon MW, Gelb BD, Gripp KW, et al. Assessing the gene-disease association of 19 genes with the RASopathies using the ClinGen gene curation framework. Hum Mutat. 2018;39:1485–93.
Johnston JJ, van der Smagt JJ, Rosenfeld JA, Pagnamenta AT, Alswaid A, Baker EH, et al. Autosomal recessive Noonan syndrome associated with biallelic LZTR1 variants. Genet Med. 2018;20:1175–85.
Vissers LELM, Bonetti M, Paardekooper Overman J, Nillesen WM, Frints SGM, de Ligt J, et al. Heterozygous germline mutations in A2ML1 are associated with a disorder clinically related to Noonan syndrome. Eur J Hum Genet. 2015;23:317–24.
van Trier DC, van Nierop J, Draaisma JMT, van der Burgt I, Kunst H, Croonen EA, et al. External ear anomalies and hearing impairment in Noonan Syndrome. Int J Pediatr Otorhinolaryngol. 2015;79:874–8.
Leung GKC, Luk HM, Tang VHM, Gao WW, Mak CCY, Yu MHC, et al. Integrating functional analysis in the next-generation sequencing diagnostic pipeline of RASopathies. Sci Rep. 2018;8:2421.
El Bouchikhi I, Belhassan K, Moufid FZ, Iraqui Houssaini M, Bouguenouch L, Samri I, et al. Noonan syndrome-causing genes: Molecular update and an assessment of the mutation rate. Int J Pediatr Adolesc Med. 2016;3:133–42.
Galliano M-F, Toulza E, Gallinaro H, Jonca N, Ishida-Yamamoto A, Serre G, et al. A novel protease inhibitor of the α 2 -macroglobulin family expressed in the human epidermis. J Biol Chem. 2006;281:5780–9.
Aoki Y, Niihori T, Inoue S-i, Matsubara Y. Recent advances in RASopathies. J Hum Genet. 2016;61:33–9.
Santos-Cortez RLP, Chiong CM, Reyes-Quintos MRT, Tantoco MLC, Wang X, Acharya A, et al. Rare A2ML1 variants confer susceptibility to otitis media. Nat Genet. 2015;47:917–20.
Santos-Cortez RLP, Reyes-Quintos MRT, Tantoco MLC, Abbe I, Llanes EGdV, Ajami NJ, et al. Genetic and environmental determinants of otitis media in an indigenous filipino population. Otolaryngol Head Neck Surg. 2016;155:856–62.
Amber KT, Valdebran M, Grando SA. Paraneoplastic autoimmune multiorgan syndrome (PAMS): beyond the single phenotype of paraneoplastic pemphigus. Autoimmun Rev. 2018;17:1002–10.
Numata S, Teye K, Tsuruta D, Sogame R, Ishii N, Koga H, et al. Anti-Alpha-2-Macroglobulin-Like-1 autoantibodies are detected frequently and may be pathogenic in paraneoplastic pemphigus. J Invest Dermatol. 2013;133:1785–93.
Richards S, Aziz N, Bale S, Bick D, Das S, Gastier-Foster J, et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet Med. 2015;17:405–24.
Gripp KW, Morse LA, Axelrad M, Chatfield KC, Chidekel A, Dobyns W, et al. Costello syndrome: Clinical phenotype, genotype, and management guidelines. Am J Med Genet A. 2019;179:1725–44.
Razzaque MA, Nishizawa T, Komoike Y, Yagi H, Furutani M, Amo R, et al. Germline gain-of-function mutations in RAF1 cause Noonan syndrome. Nat Genet. 2007;39:1013–7.
Bochet P, Ramond F, Touraine R, Thomas T, Marotte H. Arthritis associated to cardio-facio-cutaneous syndrome related to a MAP2K1 mutation. Joint Bone Spine. 2019;87:169.
van der Burgt I. Noonan syndrome. Orphanet J Rare Dis. 2007;2:4.
Acknowledgements
We thank the subjects and their families for participating in this study.
Funding
This work was supported by the ERN-ITHACA networking (AV and MT) and grants from the German Federal Ministry of Education and Research—BMBF (German Network for RASopathy Research “GeNeRARe”, FKZ: 01GM1519A; European Joint Programme on Rare Diseases (NSEuroNet) to MT, HC, MZ, FKZ: 01GM1602A), from the German Research Foundation (DFG; ZE 524/10-1) to MZ, from AIRC (IG 21614) to MT, from the Ministry of Health (RC2018 and RC2019) to ADL and MT and from the Ministry of Health, Czech Republic and National Center for Medical Genomics to MH (IP6003/0064203 and LM2018132). Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Brinkmann, J., Lissewski, C., Pinna, V. et al. The clinical significance of A2ML1 variants in Noonan syndrome has to be reconsidered. Eur J Hum Genet 29, 524–527 (2021). https://doi.org/10.1038/s41431-020-00743-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41431-020-00743-3
This article is cited by
-
Germline variants in patients developing second malignant neoplasms after therapy for pediatric acute lymphoblastic leukemia—a case-control study
Leukemia (2024)
-
Molecular and clinical profile of patients referred as Noonan or Noonan-like syndrome in Greece: a cohort of 86 patients
European Journal of Pediatrics (2022)
