
Abstract
Excessive protein synthesis upon enhanced cell proliferation frequently results in an increase of unfolded or misfolded proteins. During hematopoietic regeneration, to replenish the hematopoietic system, hematopoietic stem cells (HSCs) are activated and undergo a rapid proliferation. But how the activated HSCs respond to the proliferation pressure is still ambiguous; The proper control of the functional reservoir in the activated HSCs remains poorly understood. Here, we show a significant upregulation of EVA1A protein associated with the increase of ER stress during hematopoietic regeneration. Deletion of Eva1a significantly enhances the regeneration capacity of HSCs by inhibiting the ER stress-induced apoptosis. Mechanistically, the expression of EVA1A protein was upregulated by CHOP, and thereby promoted the ER-mitochondria interlinking via MCL1, which resulted in mitochondria-mediated apoptosis. These findings reveal a pathway for ER stress responses of HSCs by the EVA1A mediated apoptosis, which play an important role in HSCs regeneration.
Similar content being viewed by others
Introduction
The hematopoietic system is sustained by hematopoietic stem cells (HSCs), which have the capacity for self-renewal and differentiating into all lineages of mature blood cells. Although HSCs are mostly quiescent during homeostasis, they can be quickly activated to replenish the hematopoietic system in response to various stresses, such as blood loss, infections/inflammation or chemotherapy-induced myelotoxicity [1, 2]. The regeneration capacity of HSCs is affected by multiple stress stimuli, including reactive oxygen species (ROS), nutrient fluctuation, DNA damage, and ER stress [3,4,5]. HSCs maintenance during homeostatic state has been widely investigated, however the proper control of the functional reservoir in the activated HSCs remains poorly understood.
The endoplasmic reticulum (ER) is an organelle responsible for many cellular processes, such as protein folding and secretion, lipid synthesis, and calcium storage. ER membrane physically and functionally interacts with many intracellular membranous structures that put ER at a great position to sense cellular perturbations and activate signaling pathways to restore homeostasis [6, 7]. Many sources of stress, such as viral infections, ROS, and proliferative signals, cause misfolded proteins accumulation in ER and subsequently activate unfolded protein response (UPR) to restore the protein-folding homeostasis by reducing protein synthesis and increasing protein-folding and degradative capacities of ER [8, 9]. But if the UPR cannot resolve the stress, apoptosis will be initiated to clear the damaged cells.
Although HSCs have profoundly restricted protein synthesis, recent studies suggest an essential role of ER stress response/UPR in HSCs regulation [10,11,12]. Activation of the ER stress signaling IRE1α–XBP1 preserves HSC self-renewal after exposure to LPS [10]. Reducing ER stress levels by tauroursodeoxycholic acid maintains functional HSCs in vitro [11]. Alleviation of ER stress by bile acids is required for HSC expansion during fetal hematopoiesis [12]. In addition, HSCs are more susceptible to ER stress and initiate apoptosis, which may contribute to the HSC pool clonal integrity by selectively removing damaged HSCs [5, 11]. To date, despite the significance of ER stress response in HSCs biology is approved, how ER stress regulates HSCs apoptosis and the molecular mechanism remains to be elucidated.
EVA1A, also known as TMEM166 (transmembrane protein 166) or FAM176A (family with sequence similarity 176 member A), is an ER membrane protein involved in autophagy and apoptosis [13]. It has been shown that EVA1A is elevated during embryonic neurogenesis and enhances neural stem cells self-renewal via promoting autophagy [14]. However, the connection between EVA1A and apoptosis in stem cell biology is still unknown. In this study, our data show a significant increase of ER stress with an enhanced expression of EVA1A protein in hematopoietic stem/progenitor cells (HSPCs) under hematopoietic pressure. Deletion of Eva1a significantly enhances the self-renewal and repopulation capacity of HSPCs in mice via inhibiting the ER stress-induced apoptosis. The mechanism investigation shows during ER stress, C/EBP homologous protein (CHOP) upregulated the expression of EVA1A protein, which promoted the ER-mitochondria-mediated apoptosis by interacting with MCL1.
Results
ER stress promotes EVA1A expression via CHOP
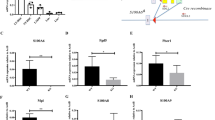
EVA1A is known as an ER membrane protein, but little is known about its role in the function of ER. To investigate a potential role of EVA1A in ER, we measured the change of EVA1A expression during ER stress. Interestingly, with the ER stress response, increased protein levels and mRNA levels of EVA1A were detected in cells treated with two chemical inducers of ER stress: thapsigargin and tunicamycin (Fig. 1A-D), suggesting a role of EVA1A in ER stress response.

A Western blot analysis of CHOP and EVA1A protein levels at indicated time points in cells treated with 3 μg/ml Tunicamycin (TM). B Western blot analysis of ATF4 and EVA1A protein levels at indicated time points in MEF cells treated with 0.2 μM Thapsigargin (TG). C, D Relative mRNA level of EVA1A in MEF cells treated with TM (C) or TG (D) at indicated time points (n = 3). E The protein level of EVA1A in WT and Chop KO MEF cells treated without or with 3 μg/ml TM for 36 h. F, G The protein level of EVA1A in IRE1 knockdown (F) or ATF6 knockdown (G) MEF cells treated without or with 3 μg/ml TM for 36 h. H Western blot analysis of the EVA1A protein level in Flag-CHOP overexpressing HEK293 cells. I The relative mRNA levels of EVA1A in HEK293 cells overexpressing Flag-CHOP (n = 3). Data are presented as mean ± SD, (**p < 0.01, and ***p < 0.001).
EVA1A is an ER membrane protein involved in apoptosis [13]. CHOP is the major transcription factor regulating ER stress-induced apoptosis. To investigate a potential role of CHOP in the ER stress-induced EVA1A expression, the expression of EVA1A were assessed in Chop knockout cells treated with or without tunicamycin. The results showed a significant increase of EVA1A protein level in the wild type (WT) cells, but not in Chop knockout cells upon tunicamycin treatment (Fig. 1E). Interestingly, the upregulation of EVA1A protein induced by tunicamycin treatment was not affected by IRE1 and ATF6 knockdown (Fig. 1F, G). Consistently, overexpression of CHOP dramatically promoted the expression of EVA1A (Fig. 1H, I). Taken together, these data indicate that ER stress promotes EVA1A expression via CHOP.
ER stress upregulates EVA1A expression during hematopoietic regeneration
Enhanced cell proliferation frequently results in an accumulation of unfolded and misfolded proteins, which is a major inducing factor of the ER stress [9, 11, 15]. To investigate how the activated HSCs responds to the proliferative stress during hematopoietic regeneration, we checked the ER stress level in HSPCs under hematopoietic stress such as 5FU or transplantation treatment. Consistently with the previous study, we found that 5FU strongly induced SLAM-HSC (Lin−cKit+Sca1+CD48−CD150+) proliferation (Fig. 2A). Interestingly, with the increase of HSCs proliferation, a significant accumulation of protein aggregation was detected in SLAM-HSCs with 5FU treatment (Fig. 2B), as demonstrated by the enhanced ProteoStat staining, a specific fluorescent dye sensitive for detecting protein aggregation. Meanwhile, a significant increase of the mRNA and protein levels of CHOP, a specific ER stress response protein, were detected in LSK (Lin−cKit+Sca1+) and Lin− (Lineage−) bone marrow (BM) cells with 5FU treatment (Fig. 2C, D), suggesting an enhanced ER stress in HSPCs under hematopoietic stress. In addition, with the ER stress response, increased mRNA levels and protein levels of EVA1A were detected in the same HSPCs with 5FU treatment (Fig. 2C, D), suggesting an ER stress-induced EVA1A expression during hematopoietic regeneration.

A–D WT mice were treated with a single dose of 5FU (150 mg/kg) and sacrificed at day 8 posttreatment. A The results present the percentage of Ki67-positive cells in SLAM-HSCs (Lin−Sca1+cKit+CD48− CD150+) after 5FU treatment (n = 4). B Representative FACS histogram (left) and quantification (right) of the level of protein aggregation in SLAM-HSCs after 5FU treatment (n = 4). C Relative mRNA levels of CHOP and EVA1A in LSK (Lin−cKit+Sca1+) cells from WT mice after 5FU treatment (n = 9). D Western blot analysis of CHOP and EVA1A protein levels in Lin− (Lineage−) cells from WT mice after 5FU treatment. E Relative mRNA levels of CHOP and EVA1A in LSK cells from lethally irradiated mice after transplanted (TP) with 2 × 106 BM cells 14 days. (n = 9) F Western blot analysis of CHOP and EVA1A protein levels in Lin− cells from mice as in (E). G Relative mRNA level of EVA1A in LSK cells from WT mice treated with 0.5 mg/kg Tunicamycin for 1 Week (n = 3). H The relative mRNA level of EVA1A in LSK cells from WT and Chop KO mice (n = 3). Data are presented as mean ± SD, (*p < 0.05, **p < 0.01, and ***p < 0.001).
To further confirm the ER stress-induced EVA1A expression during hematopoietic regeneration, the ER stress and EVA1A expression levels were also assessed in HSPCs with transplantation treatment. The results showed, with the ER stress response, the expression of EVA1A was also dramatically increased in the HSPCs after transplantation (Fig. 2E, F), as demonstrated by the upregulated mRNA and protein levels of EVA1A and CHOP in LSK and Lin− BM cells from lethally irradiated mice transplanted with 2 × 106 BM cells 14 days, consistently with our previous study showing increased proliferation and protein aggregation in the HSPCs after transplantation [16].
Lastly, to further confirm the enhanced EVA1A expression in HSPCs was stimulated by ER stress, the EVA1A expression was further analyzed in LSK cells treated with tunicamycin. The data show an enhanced EVA1A mRNA level in the LSK cells from the mice treated with tunicamycin (Fig. 2G). Consistently, Chop knockout also significantly inhibited the expression of EVA1A in LSK cells from Chop deficient mice (Fig. 2H). Taken together, these data suggest a role of EVA1A in ER stress response reactions of HSPCs during hematopoietic regeneration.
Eva1a deficiency increases the number of HSPCs
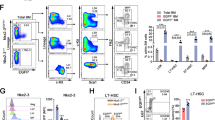
To investigate the function of EVA1A in HSPCs, we generated two hematopoietic-specific Eva1a deletion mice lines: a pIpC-inducible mouse model (Eva1aflox/floxMx-Cre, hereafter referred to as Eva1aF/F,Mx-Cre) and a genetic knockout mouse model (Eva1aflox/floxVav-Cre, hereafter referred to as Eva1aF/F,Vav-Cre) (Fig. S1A-D). In peripheral blood (PB), Eva1a depletion (Eva1aF/F,Vav-Cre) resulted in a significant increase in the frequency of B cells (B220+) and a decrease of the T cells (CD4+CD8+) frequency (Fig. 3A, B). Further analysis showed that compared with the WT mice, the absolute number of the BM cells, LSK cells, LT-HSC (Lin−cKit+Sca1+Flt3−CD150+CD48−) cells, ST-HSC (Lin−cKit+Sca1+Flt3−CD150−CD48−) cells, MPP4 (Lin−cKit+Sca1+Flt3+) cells, MPP3 (Lin−cKit+Sca1+Flt3−CD48+CD150−) cells, and MPP2 (Lin−cKit+Sca1+Flt3−CD48+CD150+) cells were increased in Eva1aF/F,Vav-Cre mice (Fig. 3C, D and Fig. S2A-C). Meanwhile, the number of MEP (Lin−cKit+Sca1−CD34−CD16/32−) cells, CMP (Lin−cKit+Sca1−CD34+CD16/32−) cells and GMP (Lin−cKit+Sca1−CD34+CD16/32+) cells were also dramatically increased in Eva1aF/F,Vav-Cre mice with the comparable CLP (Lin−cKitmidSca1midFlt3+IL−7R+) cells (Fig. 3E, F). These data indicate an increase of HSPCs in Eva1a deficient mice.

A, B Representative FACS plots (A) and Quantification of the composition (B) of T (CD4+/CD8+) cells, B (B220+) cells, and myeloid (GR1+CD11b+) in PB from WT and Eva1aF/F,Vav-Cre mice (n = 5). C, D Representative FACS plots (C) and quantification of the absolute number (D) of LSK (Lin−cKit+Sca1+) cells, LT-HSC (Lin−cKit+Sca1+Flt3−CD150+CD48−) cells, ST-HSC (Lin−cKit+Sca1+Flt3−CD150−CD48−) cells, MPP4 (Lin−cKit+Sca1+Flt3+) cells, MPP3 (Lin−cKit+Sca1+Flt3−CD48+CD150−) cells and MPP2 (Lin−cKit+Sca1+Flt3−CD48+CD150+) cells in WT and Eva1aF/F,Vav-Cre mice (n = 5). E, F Representative FACS plots (E) and quantification of the absolute number (F) of CMP (Lin−cKit+Sca1−CD34+CD16/32−) cells, GMP (Lin−cKit+Sca1−CD34+CD16/32+) cells, MEP (Lin−cKit+Sca1−CD34−CD16/32−) cells and CLP (Lin−cKitmidSca1midFlt3+IL-7R+) cells in WT and Eva1aF/F,Vav-Cre mice (n = 4). Data are presented as mean ± SD, (*p < 0.05, **p < 0.01, and ***p < 0.001).
To further confirm the HSPCs increase induced by Eva1a deletion, the absolute number of the HSPCs was also assessed in Eva1aF/F,Mx-Cre mice with seven injections of pIpC every other day. Consistently, the FACS analysis showed a significant increase in the cell number of the LSK cells, LT-HSC cells, ST-HSC cells, MPP4 cells, MPP3 cells in Eva1a deficient mice mediated by Mx-Cre (Fig. S2D). Meanwhile, a dramatically increased number of CMP and GMP cells, but not MEP and CLP cells, were also observed in Eva1aF/F,Mx-Cre mice (Fig. S2E).
Eva1a deletion enhances the regeneration capacity of HSPCs
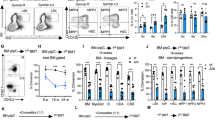
To investigate the role of EVA1A in the hematopoietic reconstitution capacity of HSPCs, we conducted competitive transplantation experiments. 1000 LSK cells sorted from WT and Eva1aF/F,Vav-Cre mice (CD45.2) were respectively mixed with 5 × 105 competitor BM cells (CD45.1) and transplanted into lethally irradiated recipient mice (CD45.1/2). Three months post-transplantation, the donor-derived cells in PB and BM were examined (Fig. S3A). The results showed that same as the donor chimerism of Eva1a deficient cells in PB, the donor chimerism of the Eva1a deficient LSK, ST (Flt3−CD34+LSK), MPP (Flt3+CD34+LSK), and LK (Lin−cKit+Sca1−) cells were dramatically enhanced in the BM of recipient mice as compared with the WT cells (Fig. S3B, C). These data suggest an enhanced regeneration capacity of the Eva1a deficient HSPCs.
To further dissect the regeneration capacity of the Eva1a deficient HSPCs, we performed serial competitive transplantation experiment, in which the transplanted HSCs experience extensive long-term regenerative stress. 300 SLAM-HSC (Lin−cKit+Sca1+CD48−CD150+) cells sorted from WT and Eva1aF/F,Vav-Cre mice (CD45.2) were respectively mixed with 5 × 105 competitor BM cells (CD45.1) and transplanted into lethally irradiated recipient mice (CD45.1/2), and four months later, the total BM cells were isolated from the primary recipients, and transplanted into the secondary recipients. The donor-derived cells in PB and BM were examined at the first and second round of transplantation (Fig. 4A). The results showed that compared with the WT cells, Eva1a depletion significantly enhanced the donor chimerism of the BM, LSK, LK, LT, ST, and MPP cells in the BM of recipient mice, and the percentage of donor-derived cells in PB of recipient mice after both the first and second transplantation (Fig. 4B-E). Meanwhile, the same serial competitive transplantation experiment was performed by the donor-derived HSCs from the pIpC-induced Eva1a deficient mice (Eva1aF/F,Mx-Cre). And a similar promotion of the regeneration capacity was observed in Eva1a deficient HSCs from pIpC-induced Eva1a deficient mice (Fig. S3D-H). These data indicate that Eva1a deletion promotes the regeneration capacity of the HSCs. In addition, to further investigate whether the regeneration capacity promotion of Eva1a deficient HSCs was due to the engraftment of the HSCs, the WT and Eva1aF/F,Mx-Cre HSCs were first transplanted without inducing Eva1a deletion. Then the recipient mice were injected with pIpC at 1 month after transplantation to induce Eva1a deletion, followed by PB chimerism analysis and secondary transplantation (Fig. 4F). The results showed Eva1a deletion similarly enhanced the regeneration capacity of HSCs after both the first and second transplantation (Fig. 4G-J). Those data confirmed that Eva1a deletion enhanced the regeneration capacity of the HSCs in a cell-intrinsic manner.

A Experimental schematic for serial competitive transplantation with SLAM-HSC (Lin−Sca1+cKit+CD48−CD150+) cells from WT and Eva1aF/F,Vav-Cre mice (results in B-E). B, D The percentage of donor-derived cells in PB at the indicated time points during 1st round (B) and 2nd round (D) of the transplantation (n = 5). C, E The Percentage of donor-derived BM, LSK (Lin−cKit+Sca1+), LK (Lin−cKit+Sca1−), LT (Flt3−CD34−LSK), ST (Flt3−CD34+LSK) and MPP (Flt3+CD34+LSK) cells in the primary recipients after transplantation 4 months (C) and the secondary recipients (E) after transplantation 3 months (n = 5). F Experimental schematic for serial competitive transplantation with SLAM-HSC cells from WT and Eva1aF/F,Mx-Cre mice (results in G–J). G, I The percentage of donor-derived cells in PB at the indicated time points during 1st round (G) and 2nd round (I) of the transplantation (n = 9~12). H, J The percentage of donor-derived BM, LSK, LK, LT, ST and MPP cells in the primary recipients after transplantation 4 months (H) and the secondary recipients (J) after transplantation 3 months (n = 9~12). K Survival curve of WT and Eva1aF/F,Vav-Cre mice following sequential 5FU treatment. 5FU was injected into mice every week (Black arrow). (n = 10 mice per group). L Absolute number of the LSK, LT, ST and MPP cells from WT and Eva1aF/F,Vav-Cre mice at day 8 after 5FU treatment (n = 4). Data are presented as mean ± SD, (*p < 0.05, **p < 0.01, and ***p < 0.001).
Lastly, to examine the regeneration capacity of Eva1a deficient HSPCs after hematopoietic ablation, Eva1a deficient mice were treated with 5FU, the number of the HSPCs and the survival rate of the mice were assessed. Eva1a deletion significantly enhanced the survival rate of the mice treated with 5FU (Fig. 4K). Meanwhile, the Eva1a deficient mice have more LSK, LT, ST, and MPP cells after 5FU treatment 8 days (Fig. 4L). Taken together, these data indicate an enhanced regeneration capacity of the Eva1a deficient HSPCs.
Eva1a deficiency does not affect HSPC proliferation and autophagy
Previous study indicates that EVA1A is involved in autophagosome formation [17]. To investigate the mechanism underlying the enhanced regeneration capacity of the Eva1a deficient HSPCs, we first analyzed the effect of Eva1a deletion on the HSPC autophagosome formation. The results showed that Eva1a deletion did not affect the autophagosome formation of the HSPCs as demonstrated by the comparable conversion of LC3I/II between WT and Eva1a deficient Lin− BM cells, even after 5FU treatment and transplantation (Fig. S4A, C, E). In addition, the autophagic degradation was also measured in the same cells. The results showed no significant difference in the autophagic degradation protein levels, such as P62, Binp3, TOM20 and TIM23 between WT and Eva1a deficient Lin− BM cells, even after 5FU treatment and transplantation (Fig. S4B, D, F). Taken together, these data suggest that Eva1a deletion does not affect HSPCs autophagy.
Given the increase of HSPCs number in Eva1a deficient mice and the enhanced regeneration capacity of HSPCs induced by Eva1a deletion, we examined the proliferation of the Eva1a deficient HSPCs. Ki67 staining showed a comparable frequency of cycling SLAM-HSCs between WT and Eva1a deficient mice (Fig. S5A). Bromodeoxyuridine (BrdU) labeling further confirmed the comparable proliferation of the WT and Eva1a deficient SLAM-HSCs (Fig. S5B). Cycling hematopoietic cells are more sensitive to 5FU cytotoxicity [18]. To further confirm the proliferation of Eva1a deficient HSPCs, we assessed the proliferation of Eva1a deficient HSPCs under 5FU treatment. The Ki67 staining showed a comparable proliferation of the WT and Eva1a deficient LSK after 5FU treatment (Fig. S5C). Lastly, the equivalent proliferation was also observed after transplantation in Eva1a knockout LSK cells (Fig. S5D). Taken together, these data suggest that Eva1a deficiency does not affect HSPCs proliferation.
Eva1a deletion reduces HSPC apoptosis during hematopoietic regeneration
To further unravel the mechanism underlying the enhanced regeneration capacity of the Eva1a deficient HSPCs, we analyzed the HSPCs apoptosis during hematopoietic regeneration. The results showed compared with the WT mice, a significantly reduced frequency of Annexin V positive cells in LSK and SLAM-HSCs of the Eva1a deficient mice after 5FU treatment (Fig. 5A, B). Caspase 3/7 activity assay further confirmed a lower apoptosis in Eva1a deficient SLAM-HSCs after 5FU treatment than that in WT cells (Fig. 5C). Meanwhile, the apoptosis rate was also assessed in the WT and Eva1a deficient HSPCs after transplantation. Annexin V staining analysis indicated a significantly lower apoptosis in Eva1a deficient LSK cells after transplantation than that in WT cells (Fig. 5D). Consistently, compared to the WT cells, a dramatically lower caspase 3/7 activity was detected after transplantation in Eva1a deficient LSK cells (Fig. 5E). Taken together, these data suggest that Eva1a deletion reduces HSPCs apoptosis during hematopoietic regeneration.

A, B Representative FACS plots (left) and the percentage (right) of Annexin V positive LSK (Lin−cKit+Sca1+) cells (A) and SLAM-HSC (Lin−Sca1+cKit+CD48−CD150+) cells (B) in WT and Eva1aF/F,Vav-Cre mice after 5FU treatment 8 days (n = 5). C Caspase 3/7 activity of the SLAM-HSC cells from WT and Eva1aF/F,Vav-Cre mice after 5FU treatment 8 days (n = 8). D CD45.1 mice after lethal irradiation were transplanted with 1 × 106 BM cells from WT and Eva1aF/F,Vav-Cre mice for 14 days. Representative FACS plots (left) and bar graph (right) depict the percentage of Annexin V positive cells in donor-derived LSK cells (n = 5). E Caspase 3/7 activity of the LSK cells as in (D) (n = 3). Data are presented as mean ± SD, (*p < 0.05, and ***p < 0.001).
ER stress-induced EVA1A and MCL1 interaction stimulates mitochondria-associated apoptosis
To investigate the molecular mechanism underlying the apoptosis inhibition of the Eva1a deficient HSPCs, we first analyzed the localization of EVA1A under ER stress. Interestingly, a significantly mitochondrial localization of EVA1A was observed in MEF cells after tunicamycin treatment as demonstrated by the colocalization of EVA1A with TOM20, a classical mitochondrial marker (Fig. 6A). And overexpressing GFP-EVA1A in HEK293 cells induced a significant mitochondria-associated apoptosis as demonstrated by the enhanced caspase 3/7/9 activity, Annexin V positive cells and the decreased mitochondrial membrane potential (Fig. S6A-D). These data suggested a role of EVA1A in mitochondria-associated apoptosis. To confirm this, we further checked the protein levels of caspase under ER stress. The results showed compared to the WT cells, Eva1a deletion significantly inhibited the protein levels of cleaved caspase 3 and Parp accompanying by a comparable caspase 8 and cleaved caspase 12 in MEF cells treated with or without tunicamycin (Fig. 6B). Notably, a significant reduction of cleaved caspase 9, a caspase involved in mitochondria-associated apoptosis, was observed in Eva1a deficient MEF cells treated with tunicamycin (Fig. 6B). And Eva1a deletion significantly reduced the cytochrome C release from mitochondria as demonstrated by a decreased localization of the diffuse cytochrome C in Eva1a deficient MEF cells after tunicamycin treatment (Fig. 6C and Fig. S7A, B). Meanwhile, Eva1a deletion dramatically inhibited the fragmentation of mitochondrial after tunicamycin treatment (Fig. S7C). These data indicate an inhibited mitochondria-associated apoptosis in Eva1a deficient cells.

ER stress-induced EVA1A and MCL1 interaction stimulates mitochondria-associated apoptosis. A Representative images (left) and quantification (right) showing the colocalization of EVA1A and TOM20 in MEF cells treated without or with 3 μg/ml Tunicamycin (TM) for 48 h. The colocalization coefficient was represented as percentage of signals of EVA1A that were positive for TOM20. Quantifications were performed using Volocity software (n > 20 cells). Scale bar, 10 μm. B Western blot analysis of the cleaved Caspase 9, cleaved Caspase 3, cleaved Caspase 12, cleaved PARP and Caspase 8 proteins level in WT and Eva1a knockout MEF cells treated without or with 3 μg/ml TM for 48 h. C Representative images (top) and quantification (bottom) showing cytochrome C localization in WT and Eva1a knockout MEF cells treated without or with TM for 24 h (n > 35 cells). Scale bar, 20 μm. D HEK293 cells expressing GFP or GFP-EVA1A were treated with 3 μg/ml TM for 48 h and immunoprecipitated with anti-GFP antibody, then the immunocomplexes were analyzed by Western blot. E HEK293 cells were treated without or with TM and immunoprecipitated with anti-MCL1 antibody, then the immunocomplexes were analyzed by Western blot. F MEF cells were treated with TM, or TM + A1210447 for 36 h. Bar graph depicts the percentage of Annexin V positive cells (n = 3). G Caspase 3/7 activity of the MEF cells treated as in (F) (n = 5). Data are presented as mean ± SD, (**p < 0.01, and ***p < 0.001).
To further investigate the molecular mechanism underlying the inhibited mitochondria-associated apoptosis in Eva1a deficient cells, we dissected the interaction of EVA1A with a series of mitochondrial proteins. Interestingly, a significant interaction of EVA1A with MCL1, a protein involved in mitochondria-associated apoptosis regulation, was detected in cells after tunicamycin treatment accompanying by a decreased interaction of MCL1 with Bak (Fig. 6D, E). And this decreased interaction of MCL1 with Bak was abolished after Eva1a deletion (Fig. S6E). These data suggest a function of EVA1A on apoptosis by interacting with MCL1 and impairing the interaction of MCL1 with Bak. Lastly, the function of EVA1A on apoptosis via MCL1 was further confirmed by treating the cells with MCL1 inhibitor. Both the Annexin V staining and caspase 3/7 activity analysis showed MCL1 inhibitor A1210447 treatment impaired the apoptosis inhibition in Eva1a deficient MEF cells after tunicamycin treatment (Fig. 6F, G). Taken together, these data suggest during ER stress, EVA1A stimulated a mitochondria-associated apoptosis via inducing the interaction of EVA1A and MCL1.
Eva1a deficiency inhibits the mitochondria-associated apoptosis in HSPCs via MCL1 during ER stress
ER stress plays an important role in HSPC regulation [10,11,12]. In this study, our data indicate that the EVA1A expression was stimulated by ER stress during hematopoietic regeneration (Fig. 2), and Eva1a deletion reduces HSPCs apoptosis during hematopoietic regeneration (Fig. 5). These data suggest an important role of ER stress in the regeneration capacity of HSPCs regulated by EVA1A. To test this hypothesis, the number of HSPCs was also assessed in Eva1a deficient mice treated with tunicamycin, a classical ER stress inducer. The FACS analysis showed that after tunicamycin treatment, Eva1a deficient mice had more LSK, LT-HSC, ST-HSC, MPP3, MPP4, CMP, and GMP cells in BM than that in WT mice (Fig. 7A, B). Eva1a deletion significantly alleviated the number reduction of LT-HSC cells in the mice treated with tunicamycin (Fig. 7C). In addition, the apoptosis rate was assessed in the WT and Eva1a deficient HSPCs after tunicamycin treatment in vitro. Annexin V staining analysis indicated a significantly lower apoptosis in Eva1a deficient LSK cells after tunicamycin treatment than that in WT cells (Fig. 7D). Consistently, compared to the WT cells, a dramatically lower caspase 3/7 activity was observed in Eva1a deficient LSK cells after tunicamycin treatment in vitro (Fig. 7E). These data indicate Eva1a deficiency inhibits ER stress-induced apoptosis in HSPCs.

A–C WT and Eva1aF/F,Vav-Cre mice were intraperitoneally injected with 0.5 mg/kg Tunicamycin (TM) every 3 days. A The absolute number of the LSK, LT-HSC, ST-HSC, MPP2, MMP3, and MPP4 cells in the TM treated mice were measured at 7th day (n = 4). B The absolute number of CMP, GMP and MEP cells in the mice as in (A) (n = 3). C The absolute number of the LT-HSCs in the mice treated with or without TM at 7th day (n = 3). D LSK cells were sorted from WT and Eva1aF/F,Vav-Cre mice by flow cytometry, and in vitro-treated with or without 3 μg/ml TM for 36 h in SFEM medium containing 50 ng/ml SCF and 50 ng/ml TPO. Bar graph depict the percentage of Annexin V positive cells (n = 3). E Caspase 3/7 activity of the LSK cells as in (D) (n = 3). F Caspase 9 activity of the WT and Eva1a knockout LSK cells treated with or without 3 μg/ml TM for 36 h in vitro (n = 3–5). G LSK cells from WT and Eva1aF/F,Vav-Cre mice were treated with or without 3 μg/ml TM for 36 h in vitro. Representative FACS plots (left) and bar graph (right) depict the percentage of JC-1 red fluorescence negative cells (n = 3). H LSK cells from WT and Eva1aF/F,Vav-Cre mice were treated with TM or TM + A1210447 for 36 h in vitro. Bar graph depicts the percentage of Annexin V positive cells (n = 5). I Caspase 3/7 activity of the WT and Eva1a knockout LSK cells treated as in (H). (n = 5). J, K WT and Eva1aF/F,Vav-Cre mice were intraperitoneally injected with TM or TM + S63458 every 3 days. The percentage of Annexin V positive LSK (J) and SLAM-HSC (K) cells were measured at 7th day (n = 4~5). Data are presented as mean ± SD, (*p < 0.05, **p < 0.01, and ***p < 0.001).
Given ER stress-induced EVA1A and MCL1 interaction stimulates mitochondria-associated apoptosis in MEF cell, we also examined the mitochondria-associated apoptosis in HSPCs under ER stress. Consistently, compared to the WT cells, a dramatically lower caspase 9 activity was observed in Eva1a deficient LSK cells after tunicamycin treatment (Fig. 7F). Meanwhile, a higher mitochondrial membrane potential was detected in Eva1a deficient LSK cells after tunicamycin treatment than that in WT LSK cells (Fig. 7G). These data indicate an inhibited mitochondria-associated apoptosis in Eva1a deficient HSPCs during ER stress.
Lastly, the function of EVA1A on apoptosis via MCL1 was further confirmed in HSPCs by treating the HSPCs with MCL1 inhibitor in vitro and vivo. Both the Annexin V staining and caspase 3/7 activity analysis showed MCL1 inhibitor A1210447 treatment impaired the apoptosis inhibition in Eva1a deficient LSK cells after tunicamycin treatment in vitro (Fig. 7H, I). And a similar phenomenon was also detected in the LSK and SLAM-HSC cells from WT and Eva1a deficient mice treated with tunicamycin or tunicamycin and S63845 (Fig. 7J, K), another MCL1 inhibitor which shows no cytotoxicity to the hemopoietic system [19]. Taken together, these data suggest Eva1a deficiency inhibits the mitochondria-associated apoptosis in HSPCs via MCL1 during ER stress.
Discussion
In this study, we report a previously unknown function of EVA1A protein in HSPCs homeostasis and regeneration (Fig. S9). Our results indicate that EVA1A is an ER stress response protein and involves in ER-mitochondria-mediated apoptosis by interacting with MCL1. Deletion Eva1a significantly enhances the regeneration capacity of HSPCs.
Under ER stress, the fate of the cell is determined by the intensity and duration of the stress and the context of cell. The UPR is firstly activated as an adaptive response to restore the balance of ER homeostasis by restricting new protein synthesis and promoting protein folding and degradation, and apoptosis will be induced after persistent or strong ER stress. In this study, we found that hematopoietic regeneration induces a significant ER stress of HSPCs. Eva1a deletion inhibits the ER stress-induced apoptosis and enhances the regeneration capacity of HSPCs during hematopoietic regeneration. These data are consistent with previous reports that the suppression of ER stress-associated apoptosis enhances HSC reconstitution capacity [11, 12]. It is reported that compared to their progenitors, HSCs are more predisposed to apoptosis under ER stress [5]. Consistently, our data show a higher apoptosis in SLAM-HSCs compared to the LSK cells under tunicamycin-induced ER stress (Fig. 7J, K). In addition, compared to the WT mice, an enhanced number of the HSCs and their progenitors were both observed in Eva1a deficient mice after tunicamycin treatment (Fig. 7A, B). These data suggest a significant function of EVA1A on the ER stress-induced hematopoietic stem and progenitor cells apoptosis, which finally enhanced the hematopoietic stem and progenitor cell maintenance and the hematopoietic reconstitution capacity of the Eva1a deficient mice. Recently, the UPR signaling pathway, such as IRE1α–XBP1 was reported to preserve the self-renewal of HSCs under ER stress [10], suggesting the proper management of ER stress signaling is essential for maintaining HSCs homeostasis. In this study, besides the inhibition of apoptosis induced by Eva1a deletion during hematopoietic regeneration, a mild expression increases of the UPR target gene, such as Bip and Xbp1-s, but not Chop were also observed in Eva1a deficient HSPCs under steady state (Fig. S8). These data suggest Eva1a deletion may lead to a mild UPR to clean the damaged proteins, which may play a role in the increased HSPCs in Eva1a deficient mice under steady state. One of the most likely reasons is the mild ER stress may increase the threshold of the ER stress-induced HSC impairment by enhancing the chaperone protein such as Bip, and upregulate the efficiency of HSC unfolded protein response. However, how the Eva1a deficiency leads to the mild UPR under HSC steady state is not deeply investigated in this study. It will be interesting to further clarify the mechanism in future study.
CHOP is known as a key mediator of ER stress-induced apoptosis, but the molecular mechanism remains far from being completely understood. Previous studies show CHOP regulates apoptosis by transcriptionally regulating Bcl-2 family proteins such as BIM, BCL2, and PUMA [20,21,22]. However, other work showed a role of CHOP in apoptosis by transcriptionally increasing protein synthesis leading to cell death without any change of the Bcl-2 family proteins [23]. These works suggest a different mechanism of CHOP in apoptosis under different cell types or contexts. In this study, we identified a novel function of CHOP that upregulated the expression of EVA1A protein, which promoted the ER-mitochondria-mediated apoptosis by interacting with MCL1. Meanwhile, our data show Eva1a deficiency enhances the regeneration capacity of HSPCs via reducing apoptosis, which is consistent with our previous study showing Chop deletion improves HSCs regeneration through decreasing apoptosis [16]. EVA1A is an ER membrane protein involved in autophagy and apoptosis [13]. To date, the molecular mechanism of the EVA1A-induced apoptosis is still unknown. In this study, we found that ER stress induced a CHOP-dependent expression of EVA1A which interacted with MCL1, an anti-apoptotic Bcl-2 family protein required for HSC survival [24], leading to a drop in mitochondrial membrane potential and the release of cytochrome C from mitochondria which finally induced apoptosis. Regrettably, although a potential role of EVA1A in autophagosome formation was reported [17], a comparable autophagy was observed in WT and Eva1a deletion HSPCs in this study. However, it is possible that the EVA1A-involved autophagy play a role in the tissue with high metabolic activity, such as liver and heart [25, 26] or contexts under dramatical metabolic fluctuations, such as starvation [27].
MCL1 is an essential regulator of HSC self-renewal and survival [24, 28]. In this study, our data show a significant binding of EVA1A with MCL1 leading a reduction of MCL1 and Bak interaction. Eva1a deletion significantly inhibited the HSPC apoptosis during hematopoietic regeneration confirming the important role of the MCL1 in HSCs. In addition, MCL1 is required for the mitochondrial homeostasis. MCL1 deletion results in mitochondria disassemble, cristae morphological abnormalities and defects in electron-transport-chain enzymatic function [29]. Given the significant interaction of EVA1A and MCL1, a potential role of the mitochondrial homeostasis in the Eva1a deficient HSC will be interesting in the future study. In addition, we found a significant increase of B cells, and a decrease of T cells in the PB of Eva1a deficient mice under steady state. But Eva1a deletion did not change the number of CLP cells in BM. Given the important role of apoptosis in the development and activity of T and B cells [30], a potential role of the EVA1A in the differentiation or maturation of T and B cells will be interesting in future study.
Materials And Methods
Mice
Eva1aflox/flox (hereafter referred to as Eva1aF/F) mice were kindly provided by Dr. Chen Yingyu [14]. To obtain tissue-specific or inducible Eva1a knockout mice, the Eva1aF/F mice were crossed with Vav-Cre, Mx-Cre and ERT-Cre transgenic mice. Chop knockout mice was used as described previously [16]. B6.SJL-PtprcaPep3b/Boy (CD45.1) mice or CD45.1/CD45.2 heterozygous mice were used as recipients or competitors for HSCs transplantation [31]. All of these strains were maintained in a C57BL/6 background and were housed in a specific pathogen-free environment. The Animal Care and Ethics Committee of Jinan University approved all animal experiments in this study.
Cell culture, transfection and knockdown
MEF, Huh7 and HEK293 cells were grown in DMEM with 10% FBS at 37 °C under 5% CO2. No mycoplasma contamination was detected. Transient transfection was performed by Lipofectamine 3000/2000 (Invitrogen) according to the manufacturer’s instructions. For the IRE1 and ATF6 knockdown, siRNA duplexes designed against conserved targeting sequences were transfected into cells at a final concentration of 20 nM using Lipofectamine 2000 as specified by the manufacturer. The following siRNA duplexes were used: 5′-GCUGAACUACUUGAGGAAUUA-3′ for IRE1; 5′-GGCAAAGCAGCAGUCGAUUAU-3′ for ATF6. For liquid culture, HSPCs were cultured in serum-free expansion medium (SFEM) (Stem cell Technologies) with 50 ng/ml stem cell factor (Pepro Tech), and 100 U/ml penicillin/streptomycin.
Flow cytometry and cell sorting
Bone marrow (BM) cells from femurs, tibiae, and iliac crests were isolated as reported previously [31]. Before cell sorting, BM cells were first enriched with anti-antigen-presenting cell microbeads (MiltenyiBiotec) and then stained with surface markers. Prepared samples were analyzed on the LSR FortessaTM cell analyzer (BD Biosciences) or sorted on the FACS AriaIII cell sorter (BD Biosciences). Peripheral blood from the postorbital vein was collected and analyzed on LSR FortessaTM cell analyzer (BD Biosciences). Detailed methods and antibodies were as our lab paper described previously [31].
Chemical Treatment
For 5FU (Sigma) treatment, mice were intraperitoneally injected 5FU at 150 mg/kg and sacrificed at the indicated time point. For 5FU treatment survival assay, the mice were persistently and intraperitoneally injected 5FU at 150 mg/kg every week. For the pIpC (Sigma) treatment, mice received 20 mg/kg every other day for 2 weeks. For the tunicamycin treatment, the mice were intraperitoneally injected 0.5 mg/kg tunicamycin every 3 days and sacrificed at the indicated time point. For S63845 (APExBIO) treatment, mice were intraperitoneally injected S63845 at 10 mg/kg every 3 days and sacrificed at the indicated time point. For the in vitro experiments, HSPCs were sorted and cultured in SFEM (Stem cell Technologies) with 50 ng/ml SCF and 50 ng/ml TPO, then were treated with 3 μg/ml tunicamycin or 10 μM A1210447 as indicated in the figure legends. Cell lines were cultured in DMEM with 10% FBS, and treated with 3 μg/ml tunicamycin or 0.2 μM thapsigarginas indicated in the figure legends.
Immunoprecipitation and western blot
For immunoprecipitation, cells were lysed with NP40 lysis buffer (50 mM Tris-HCl pH 7.5, 100 mM NaCl, 1% NP-40, 1 mM EDTA, 1 mM DTT, 10% glycerol) containing protease inhibitors. After centrifugation, the supernatants were incubated with antibody overnight and then Protein A/G agarose for 2 h at 4 °C. Immunocomplexes were washed and analyzed by Western blot. For Western blot, the proteins from lysed cells were denatured and separated with SDS-PAGE. Then, the proteins were transferred to PVDF membranes, blocked and incubated with the corresponding primary and secondary antibodies. The specific bands were analyzed by the Western blot infrared imaging system (LI-COR Biosciences). The following antibodies were used: EVA1A: A8070, ABclonal; CHOP: 2895, Cell signaling Technology (CST); IRE1: 14C10, CST; ATF6: 65880 T, CST; Actin: AC026, ABclonal; Flag: 20543-1-AP, Proteintech; Caspase 8: 4927, CST; C-Caspase 9: 9509, CST; C-Caspase 3: 9664, CST; Caspase 12: 2202, CST; C-Parp: 9548, CST; GFP: ab290, Abcam; MCL1: 94296, CST; Bak: 12105, CST; LC3: 27543,Sigma; P62: 18420-1-AP, Proteintech; BNIP3: ab10433, Abcam; TIM23: 111263-A, Proteintech; TOM20: ab56783, Abcam.
Immunostaining and confocal microscopy
For immunostaining, cells were fixed in 4% formaldehyde in PBS for 10 minutes at room temperature. After washing with PBS, they were incubated in PBS containing 10% FCS to block nonspecific sites of antibody adsorption. Then, the cells were incubated with appropriate primary and secondary antibodies in 0.1% saponin as indicated in the figure legends. Images were taken in multi-tracking mode on a laser scanning confocal microscope (LSM880, Carl Zeiss) with a 63х plan apochromat 1.4 NA objective. The following antibodies were used: EVA1A: NBP1-92517, NOVUS; TOM20: ab56783, Abcam; Cytochrome C: ab133504, Abcam.
RNA isolation and Real-time PCR Analysis
Total RNA was isolated from the cells using RNeasy Micro Kit (Qiagen) according to the manufacturer’s instructions. RNA was reversed by the PrimeScript cDNA synthesis Kit (Takara). The quantitative-PCR was performed on a 7300 Real-Time PCR system for 40 cycles. The expression of β-actin was used as the internal control. At least three biological replicates were performed for each experiment.
Apoptosis and cell cycle assays
For apoptosis assay, BM cells were stained with antibodies for the surface markers, and incubated with the Annexin V-FITC antibody at room temperature for 15 minutes. DAPI (1 μg/ml) was added before analysis by FACS. For the in vitro apoptosis assay, the cells treated with the drug in vitro were directly stained with the Annexin V-FITC antibody, and DAPI was added before analysis by FACS. For caspase 3/7, 9 activity assays, HSPCs were plated into 96-well white luminescence plates and the Caspase3/7, 9 activities were analyzed according to the manufacturer’s instructions (Promega). For the mitochondrial membrane potential assay, the cells treated with the drug in vitro were determined by FACS after loading the cells with JC-1 dye according to the manufacturer’s instructions (Beyotime). For the BrdU incorporation assay, BrdU (100 mg/kg) was intraperitoneally injected, and followed by administration of BrdU (1 mg/ml) in the drinking water for 10 days. BrdU incorporation was determined by FACS analysis using the BrdU Flow Kit (BD Biosciences). For Ki67 staining, after the surface markers staining, the cells were fixed using BrdU Flow Kit and staining with Ki67 antibody (BD Pharmingen).
Transplantation analysis
Competitive transplantation was performed by transplanting 300 SLAM-HSCs (CD150+CD48−Lin−Sca1+cKit+) sorted from donor mice (CD45.2) with 5 × 105 BM cells from age-matched WT mice (competitor, CD45.1) into lethally irradiated recipient mice (CD45.1/CD45.2). For the secondary transplantation, 1 × 106 chimeric BM cells from the primary recipients were transplanted into the secondary recipient mice at 4th month after primary transplantation. Peripheral blood cells from recipients were analyzed over time. Donor-derived bone marrow cells were collected from recipient mice at indicated time point after transplantation.
Protein aggregation analysis
Cells were fixed and permeabilized by BD Fixation/Permeabilization Solution kit (BD Biosciences). The protein aggregation level in the cells were analyzed by FACS using ProteoStat Dye (Enzo Life Sciences).
Statistical analyses
The sample sizes were described in the figure legend and were determined as at least three biologically independent animals according to previous studies performed by our group. Animals with the same genotype and gender and similar age (3~4 months) were randomly assigned to experimental groups. Investigators were not blinded during the group allocation during the experiment. All statistical analyses were performed with GraphPad Prism 7 software. Data are presented as the mean ± SD. The statistical significance of the differences between groups was calculated using the unpaired Student’s two-tailed t-test. The survival curve was analyzed by using a log-rank (Mantel-Cox) test. The significance level was set at 0.05. *p < 0.05, **p < 0.01, ***p < 0.001.
Data availability
The datasets supporting the conclusions of this article are included within the article and its Additional files.
References
Baldridge MT, King KY, Boles NC, Weksberg DC, Goodell MA. Quiescent haematopoietic stem cells are activated by IFN-gamma in response to chronic infection. Nature 2010;465:793–7.
Wilson A, Laurenti E, Oser G, van der Wath RC, Blanco-Bose W, Jaworski M, et al. Hematopoietic stem cells reversibly switch from dormancy to selfrenewal during homeostasis and repair. Cell 2008;135:1118–29.
Geiger H, de Haan G, Florian MC. The ageing haematopoietic stem cell compartment. Nat Rev Immunol. 2013;13:376–89.
Yahata T, Takanashi T, Muguruma Y, Ibrahim AA, Matsuzawa H, Uno T, et al. Accumulation of oxidative DNA damage restricts the self-renewal capacity of human hematopoietic stem cells. Blood 2011;118:2941–50.
van Galen P, Kreso A, Mbong N, Kent DG, Fitzmaurice T, Chambers JE, et al. The unfolded protein response governs integrity of the haematopoietic stem-cell pool during stress. Nature 2014;510:268–72.
Rutkowski DT, Hegde RS. Regulation of basal cellular physiology by the homeostatic unfolded protein response. J Cell Biol. 2010;189:783–94.
Frakes AE, Dillin A. The UPRER: sensor and coordinator of organismal homeostasis. Mol Cell. 2017;66:761–71.
Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science 2011;334:1081–6.
Hetz C. The unfolded protein response: controlling cell fate decisions under ER stress and beyond. Nat Rev Mol Cell Biol. 2012;13:89–102.
Liu L, Zhao M, Jin X, Ney G, Yang KB, Peng F, et al. Adaptive endoplasmic reticulum stress signalling via IRE1α-XBP1 preserves self-renewal of haematopoietic and pre-leukaemic stem cells. Nat Cell Biol. 2019;21:328–37.
Miharada K, Sigurdsson V, Karlsson S. Dppa5 improves hematopoietic stem cell activity by reducing endoplasmic reticulum stress. Cell Rep. 2014;7:1381–92.
Sigurdsson V, Takei H, Soboleva S, Radulovic V, Galeev R, Siva K, et al. Bile acids protect expanding hematopoietic stem cells from unfolded protein stress in fetal liver. Cell Stem Cell. 2016;18:522–32.
Wang L, Yu C, Lu Y, He P, Guo J, Zhang C, et al. TMEM166, a novel transmembrane protein, regulates cell autophagy and apoptosis. Apoptosis 2007;12:1489–502.
Li M, Lu G, Hu J, Shen X, Ju J, Gao Y, et al. EVA1A/TMEM166 regulates embryonic neurogenesis by autophagy. Stem Cell Rep. 2016;6:396–410.
Luo B, Lee AS. The critical roles of endoplasmic reticulum chaperones and unfolded protein response in tumorigenesis and anticancer therapies. Oncogene 2013;32:805–18.
Shi ZC, Diao DJ, Zhao YN, Luo Y, Li YF, Liu DD, et al. C/EBP homologous protein deficiency enhances hematopoietic stem cell function via reducing ATF3/ROS-induced cell apoptosis. Aging Cell. 2021;20:e13382.
Hu J, Li G, Qu L, Li N, Liu W, Xia D, et al. TMEM166/EVA1A interacts with ATG16L1 and induces autophagosome formation and cell death. Cell Death Dis. 2016;7:e2323.
Cheng T, Rodrigues N, Shen H, Yang Y, Dombkowski D, Sykes M, et al. Hematopoietic stem cell quiescence maintained by p21cip1/waf1. Science 2000;287:1804–8.
Kotschy A, Szlavik Z, Murray J, Davidson J, Maragno AL, Le Toumelin-Braizat G, et al. The MCL1 inhibitor S63845 is tolerable and effective in diverse cancer models. Nature 2016;538:477–82.
Puthalakath H, O'Reilly LA, Gunn P, Lee L, Kelly PN, Huntington ND, et al. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 2007;129:1337–49.
McCullough KD, Martindale JL, Klotz LO, Aw TY, Holbrook NJ. Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Mol Cell Biol. 2001;21:1249–59.
Cazanave SC, Elmi NA, Akazawa Y, Bronk SF, Mott JL, Gores GJ. CHOP and AP-1 cooperatively mediate PUMA expression during lipoapoptosis. Am J Physiol Gastrointest Liver Physiol. 2010;299:236–43.
Han J, Back SH, Hur J, Lin YH, Gildersleeve R, Shan J, et al. ER-stress-induced transcriptional regulation increases protein synthesis leading to cell death. Nat Cell Biol. 2013;15:481–90.
Opferman JT, Iwasaki H, Ong CC, Suh H, Mizuno S, Akashi K, et al. Obligate role of anti-apoptotic MCL-1 in the survival of hematopoietic stem cells. Science 2005;307:1101–4.
Lin X, Cui M, Xu D, Hong D, Xia Y, Xu C, et al. Liver-specific deletion of Eva1a/Tmem166 aggravates acute liver injury by impairing autophagy. Cell Death Dis. 2018;9:768.
Zhang S, Lin X, Li G, Shen X, Niu D, Lu G, et al. Knockout of Eva1a leads to rapid development of heart failure by impairing autophagy. Cell Death Dis. 2017;8:e2586.
Lu GD, Ang YH, Zhou J, Tamilarasi J, Yan B, Lim YC, et al. CCAAT/enhancer binding protein α predicts poorer prognosis and prevents energy starvation-induced cell death in hepatocellular carcinoma. Hepatology 2015;61:965–78.
Campbell CJ, Lee JB, Levadoux-Martin M, Wynder T, Xenocostas A, Leber B, et al. The human stem cell hierarchy is defined by a functional dependence on Mcl-1 for self-renewal capacity. Blood 2010;116:1433–42.
Perciavalle RM, Stewart DP, Koss B, Lynch J, Milasta S, Bathina M, et al. Anti-apoptotic MCL-1 localizes to the mitochondrial matrix and couples mitochondrial fusion to respiration. Nat Cell Biol. 2012;14:575–83.
Nagata S, Tanaka M. Programmed cell death and the immune system. Nat Rev Immunol. 2017;17:333–40.
Wang H, Diao D, Shi Z, Zhu X, Gao Y, Gao S, et al. SIRT6 controls hematopoietic stem cell homeostasis through epigenetic regulation of Wnt signaling. Cell Stem Cell. 2016;18:495–507.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (82030039, 92049304), the National Key Research and Development Program of China (2020YFA0112404, 2021YFA1100103), the Program for Guangdong Introducing Innovative and Entrepreneurial Teams (2017ZT07S347), the Science Foundation for Distinguished Young Scholars of Guangdong Province (2019B151502008), and the Open Project of the State Key Laboratory of Trauma, Burn and Combined Injury, Third Military Medical University (SKLKF202002).
Author information
Authors and Affiliations
Contributions
BL, and ZYJ designed the study. BL, YYZ, QFW, YTF, XLZ, and ZKW, performed experiments and analyzed data. WWY, and HW, provided technical assistance. WWY, and ZYC, edited the manuscript. ZFS, WX, WFH, and YGQ, provided additional expertise. BL, and ZYJ supervised the study and wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Professor Paolo Pinton
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, B., Zhou, Y., Wu, Q. et al. EVA1A regulates hematopoietic stem cell regeneration via ER-mitochondria mediated apoptosis. Cell Death Dis 14, 71 (2023). https://doi.org/10.1038/s41419-023-05559-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-023-05559-9
