
Abstract
Transcriptional factor EB (TFEB), a master regulator of autophagy and lysosomal biogenesis, is generally regarded as a pro-survival factor. Here, we identify that besides its effect on autophagy induction, TFEB exerts a pro-apoptotic effect in response to the cyclopentenone prostaglandin 15-deoxy-∆-12,14-prostaglandin J2 (15d-PGJ2). Specifically, 15d-PGJ2 promotes TFEB translocation from the cytoplasm into the nucleus to induce autophagy and lysosome biogenesis via reactive oxygen species (ROS) production rather than mTORC1 inactivation. Surprisingly, TFEB promotes rather than inhibits apoptosis in response to 15d-PGJ2. Mechanistically, ROS-mediated TFEB translocation into the nucleus transcriptionally upregulates the expression of ATF4, which is required for apoptosis elicited by 15d-PGJ2. Additionally, inhibition of TFEB activation by ROS scavenger N-acetyl cysteine or inhibition of protein synthesis by cycloheximide effectively compromises ATF4 upregulation and apoptosis in response to 15d-PGJ2. Collectively, these results indicate that ROS-induced TFEB activation exerts a novel role in promoting apoptosis besides its role in regulating autophagy in response to 15d-PGJ2. This work not only evidences how TFEB is activated by 15d-PGJ2, but also unveils a previously unexplored role of ROS-dependent activation of TFEB in modulating cell apoptosis in response to 15d-PGJ2.
Similar content being viewed by others


Introduction
Autophagy is a highly conserved process for regulating the degradation of protein aggregates, lipids, and cellular organelles by lysosomes to recycle cellular contents, which plays an important role in a variety of physiology and pathological conditions [1,2,3,4]. Autophagy process is involved in the formation of double-membrane autophagosomes, followed by fusion with lysosomes and the degradation of sequestered cell contents within autophagosomes by lysosomal hydrolases [1, 2]. Transcriptional factor EB (TFEB) along with transcriptional factor E3 (TFE3) are members of the basic-helix-loop-helix leucine-zipper transcription factor microphthalmia-associated transcription factor/transcriptional factor E (TFE) family [5], they were recently identified as “master regulators” of autophagy and lysosome biogenesis [5, 6]. In response to a variety of stressors such as starvation, TFEB is translocated from the cytosol into the nucleus, where it recognizes and binds to the CLEAR-box sequence (5′-GTCACGTGAC-3′) to upregulate the expression of multiple autophagy-lysosome-related genes to induce autophagy [5, 6]. TFEB activity is mainly regulated by its phosphorylation status that can be modulated by a variety of stressors, such as starvation, ER stress [7], DNA damage [8], oxidative stress [9], mitochondrial damage [10], pathogens infection [11], and exercise [12]. Upon starvation-induced mTORC1 (mammalian target of rapamycin complex 1) inhibition, TFEB is rapidly dephosphorylated and dissociated from TFEB/14-3-3 complex; free TFEB is then translocated into the nucleus to induce autophagy and lysosome biogenesis [13,14,15]. In addition, TFEB activation can also be modulated by calcium-dependent phosphatase calcineurin [16, 17]. Though the activity and nucleus accumulation of TFEB is mainly regulated by the phosphorylation status of TFEB [18], TFEB activation can also be modulated by reactive oxygen species (ROS)-dependent TFEB oxidation [9].
Apart from starvation, increasing evidences demonstrated a variety of stressors, such as ER stress [7], DNA damage [8], oxidative stress [9], mitochondrial damage [10], pathogens infection [11], and exercise [12] also promote TFEB activation. Though TFEB is generally regarded as a pro-survival factor and plays an important role in regulating protein homeostasis via autophagy-lysosome pathway, increasingly evidences show that TFEB has more broad biological functions. These functions include regulating unfolded protein response [7], the p53 dependent cell cycle [8], and immunity [11]. Notably, although prolonged ER stress-induced TFEB activation contributes to apoptosis [7], whether other conditions induced TFEB activation could promote apoptosis is of interest to be dissected.
The cyclopentenone prostaglandin 15-deoxy-∆-12,14-prostaglandin J2 (15d-PGJ2) is a metabolite of the cyclooxygenase pathway [19]. It is produced by twice dehydration of PGD2 (Prostaglandin D2) [19]. 15d-PGJ2 was identified as an endogenous peroxisome proliferator-activated receptor-γ (PPARγ) ligand in 1995 [20, 21] and has been investigated intensively for its biological function ever since. PPARγ, a member of the superfamily of nuclear receptors, regulates a variety of target genes by forming a complex with retinoid X receptor in a ligand-dependent manner [22]. PPARγ plays an important role in regulating fatty acid storage, glucose metabolism, insulin sensitivity, inflammation and apoptosis, and is implicated in multiple diseases including obesity, diabetes, and cancer [22, 23]. As an endogenous PPARγ ligand, 15d-PGJ2 exerts multiple biological activities, including antitumor, anti-inflammatory, anti-oxidative, anti-fibrotic, in a PPARγ dependent or independent manner [23,24,25,26,27]. Importantly, apart from its inflammatory modulating property [27], the antiapoptotic effect of 15d-PGJ2 has also been gaining great attention, revealing distinct mechanisms that underlie its antiapoptotic property. These include inhibiting the expression of human telomerase reverse transcriptase and telomerase activity [28]; regulating ROS-mediated Akt inhibition [29]; and infinities mitochondrial-dependent mechanism [26, 30, 31]. Importantly, autophagy plays an important role in modulating multiple cellular process including apoptosis, whether 15d-PGJ2 is an autophagy enhancer or inhibitor is still under debate [32,33,34,35], and thus underlying the role and underlying mechanisms of 15d-PGJ2 in modulating autophagy are also important.
Here, we aimed to characterize role of TFEB—a master regulator of autophagy-lysosome pathway, in regulating autophagy and apoptosis in response to 15d-PGJ2 treatment and dissect the underlying mechanisms. Our results demonstrated that the generation of ROS is responsible for the activation of TFEB, and subsequent induction of autophagy and lysosome biogenesis in response to 15d-PGJ2. Unexpectedly, TFEB activation facilitates apoptosis elicited by 15d-PGJ2 treatment, which is distinct from traditional view that cells rely on TFEB for survival. We further uncovered that the transcriptionally upregulated ATF4 is responsible for the effect of TFEB in promoting apoptosis in response to 15d-PGJ2. Conversely, inhibition of TFEB activation by ROS scavenger NAC (N-acetyl cysteine) or inhibition of protein synthesis by cycloheximide effectively compromises both autophagy pathway and ATF4 upregulation-mediated apoptosis in response to 15d-PGJ2. Overall, our results suggest a novel role of ROS-mediated TFEB activation in promoting apoptosis besides enhancing autophagy during 15d-PGJ2 treatment.
Materials and methods
Reagents and antibodies
Bafilomycin A1 (sc-201550), and anti-β-actin/ACTB (sc-47778) were purchased from Santa Cruz Biotechnology. Chloroquine (C6628) was purchased from Sigma-Aldrich. Torin 1 (2273-5) was purchased from BioVision Inc. 15d-PGJ2 (sc-201262A), FK-506 (sc-24649A), N-acetyl-L-cysteine (NAC, A9165). ATF4 siRNA (L-005125-00-0005) and nontarget siRNA were purchased from Dharmacon. LysoTracker® Red DND-99 (L-7528), DMEM (11965-126), FBS (10270-106), Opti-MEM I (31985-070) were purchased from Life Technologies. Anti-cleaved caspase 3 (CST 9662S), anti-cleaved caspase 9 (CST 9508S), anti-Bcl2 (CST 2870S), anti-phospho-P70S6K/RPS6KB1 (Thr389) (9234), anti-P70S6K/RPS6KB1 (9202), anti-phospho-EIF4EBP1/4EBP1 (Thr37/46) (2855), anti-4E-BP1 (53H11) (9644), ATF4 (D4B8), anti-CHOP (2895) and anti-H3F3A/histone H3 (D1H2) (4499) antibodies were purchased from Cell Signaling Technology. Anti-LAMP1 (ab24170) antibody was purchased from Abcam. HRP-conjugated goat anti-mouse (115-035-003) and goat anti-rabbit (111-035-003) secondary antibodies were purchased from Jackson ImmunoResearch. Anti-LC3B (NB100-2220) antibodies were purchased from Novus Biologicals. Anti-TFEB (13372-1-AP) was purchased from Proteintech. Anti-TFEB (A303-673A) was purchased from Bethyl Laboratories, Inc. Anti-GAPDH antibody (GTX100118) was obtained from GeneTex. Alexa Fluor®488 goat anti-mouse IgG (A-11001), Alexa Fluor®488 goat anti-Rabbit IgG (A-11034), Alexa Fluor 488 (A-11008) and Alexa Fluor®594 goat anti-rabbit IgG (A-11012) were purchased from Life Technologies.
Cell culture
HeLa, HT-22, and HEK293 cells were cultured in DMEM supplemented with 10% FBS. HeLa cells stably expressing 3×-Flag-TFEB [6, 36, 37] (CF-7) were maintained in DMEM supplemented with 10% FBS and 200 μg/mL G418. All cells were maintained with cell culture medium containing 50 U of penicillin-streptomycin mixture (Invitrogen) at 37 °C, gassed with 5% CO2.
Cell transfection
For knockdown experiments, cells were transfected with indicated siRNA using Lipofectamine RNAiMAX (13778030, Invitrogen). For overexpression experiments, cells were transfected with Lipofectamine™ 3000 (L3000015, Invitrogen) according to the manufacturer’s instructions for 24–72 h, then cells were treated with indicated drugs.
Western blotting analysis
Cells were washed twice with ice-cold phosphate-buffered saline (PBS) [137 mM NaCl, 10 mM Na2HPO4, 2 mM KH2PO4, 2.7 mM KCl (pH 7.4)] and then lysed on ice in 1×RIPA Lysis Buffer (9803, Life Technologies) with complete protease inhibitor mixture (04693124001, Roche Applied Science) and phosphatase inhibitor (B15001, Biotool). Protein concentrations were determined by Bio-Rad Bradford assays using bovine serum albumin (BSA) as standard. Equal volumes of Laemmli 2× buffer [4% SDS, 10% β-mercaptoethanol, 20% glycerol, 0.004% bromophenol blue, 0.125 M Tris-HCl (pH 6.8)] and protein lysate were mixed and denatured in boiling water for 10 min. Proteins were separated by 10%–15% SDS-PAGE, transferred, and blotted with the antibodies described. The blots were then incubated with primary and secondary antibodies. The protein signals were detected by the ECL kit (32106, Pierce) and captured by X-ray film (Fujifilm). Western blot results were quantified by using ImageJ software.
Isolation of the cytosol and the nucleus fractions
Cytosol and nucleus extracts were prepared according to a previous protocol [17, 38]. In brief, at the end of drug treatment, the cells were washed with ice-cold PBS, centrifuged, and re-suspended in cold lysis buffer containing 20 mM N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid (HEPES), pH 8.0, 1 mM ethylenediaminetetraacetic acid (EDTA), 1.5 mM MgCl2, 10 mM KCl, 1 mM DTT, 1 mM sodium orthovanadate, 1 mM NaF, 1 mM PMSF, 0.5 mg/mL benzamidine, 0.1 mg/mL leupeptin, and 1.2 mg/mL aprotinin. The cells were allowed to swell on ice for 15 min. NP-40 (10% (v/v)) was subsequently added to the cell suspensions. The samples were vortexed vigorously for 10 s. The homogenates were centrifuged for 50 s at 16,000 × g, and the supernatant was used as cytosolic extract. The nuclear pellet was re-suspended in cold extraction buffer containing 20 mM HEPES, pH 8.0, 1 mM EDTA, 1.5 mM MgCl2, 10 mM KCl, 1 mM DTT, 1 mM sodium orthovanadate, 1 mM NaF, 1 mM PMSF, 0.5 mg/mL benzamidine, 0.1 mg/mL leupeptin, 1.2 mg/mL aprotinin, and 20% glycerol. All the protein fractions were stored at −30° C until use.
Determination of ROS
ROS levels were measured by the oxidation of 2′,7′-dichlorofluorescin diacetate (DCFDA) to a fluorescence dye 2′, 7′-dichlorofluorescein (DCF) according to a previous protocol [39]. Briefly, after cells were treated with 15d-PGJ2 as indicated, cells were loaded with DCF-DA (100 μM) for 1 h. Green fluorescence was analyzed by flow cytometry (BD Accuri C6 Plus Flow Cytometer). Data analysis was done using BD FACSVia Research Loader software or FlowJo.
Apoptosis assay
The extent of apoptosis was evaluated by Annexin V-FITC/PI staining and a flow cytometer. Briefly, at the end of drug treatment, cells were harvested, washed with PBS (pH 7.4), centrifuged, and stained with Annexin V-FITC (Pharmingen) and 2 μg/mL propidium iodide in binding buffer [10 mmol/L Hepes (pH 7.4), 140 mmol/L NaCl, 2.5 mmol/L CaCl2] for 15 min at 37 °C in the dark. The samples were analyzed by flow cytometry (BD Accuri C6 Plus Flow Cytometer). Data analysis was done using BD FACSVia Research Loader software.
Determination of lysosomal contents by using LysoTracker staining
The lysosomal contents were determined using LysoTracker Red DND99 (L7528, Thermo Fisher Scientific) staining following manufacturer’s instructions. Briefly, during the last 1 h of drug treatment, cells were loaded with 75 nM LysoTracker Red DND99. After excess dye was washed out, cells were harvested, and the fluorescence intensity was observed by a flow cytometer (BD Accuri C6 Plus Flow Cytometer).
Immunocytochemistry
Cells were seeded on coverslips placed in 24-well plates. At the end of drug treatment, slides were fixed with 3.7% paraformaldehyde, permeabilized in 0.2% Triton X-100, and blocked with 3% BSA. After blocking, the slides were stained with anti-Flag (1:600) antibodies overnight at 4 °C. Alexa Fluor®594 (red) or Alexa Fluor®488 (green) secondary antibodies (1:1000) were added for 1 h at room temperature. After nuclear staining with DAPI, the slices were mounted with FluorSave reagent (345789, Calbiochem). Cells were visualized using the API DeltaVision Personal Imaging System.
The TFEB nuclear translocation analysis was done according to a previous protocol with some modification [17, 37]. Briefly, images were acquired randomly from at least ten different fields per sample. Quantitation was done from each image for each sample, and around 200 cells per treatment from three independent experiments were performed to generate the graphed values. NIH ImageJ software was used to perform the analysis of TFEB localization in the different images. The ratio value resulting from the average intensity of nuclear TFEB fluorescence was divided by the average of the cytosolic intensity of TFEB fluorescence by using imageJ. The results were normalized using negative (DMEM medium with 10% FBS) control samples in the same plate. The data are represented by the percentage of nuclear translocation at the different concentrations of each compound using Prism software (GraphPad software).
Quantitative real-time PCR
Total RNA was extracted from cells using Trizol reagent (15596026, Thermo Fisher Scientific). Reverse transcription was performed using the High-Capacity cDNA Reverse Transcription Kit (Life Technologies, 4368814). Autophagy and lysosome gene primers were used according to our previous study [37] and synthesized by Life Technologies. The oligonucleotide sequences are listed in Table S1. Real-time PCR was carried out with the Fast SYBR Green Master Mix (Life Technologies, 4385612) using the ViiA™ 7 Real-Time PCR System (Life Technologies, Carlsbad, CA, USA). Fold changes were calculated using the ΔΔCT method, and the results were normalized against an internal control (GAPDH).
Statistical analysis
All data were analyzed by GraphPad Prism (GraphPad Software). Unless specified, data are means ± SEM. Unpaired Student’s t test was used when comparing drug treatments with vehicle group within the same cell type. Multiple comparisons between groups were performed by one-way ANOVA. A probability value of P < 0.05 was considered to be statistically significant.
Results
TFEB translocated from the cytosol to the nucleus in response to 15d-PGJ2
In normal conditions, TFEB is mainly localized in the cytosol in an inactive form [13,14,15, 40]. In response to various stressors such as starvation, TFEB rapidly moves into the nucleus to induce the expression of multiple genes for inducing autophagy and restoring homeostasis [13,14,15, 40]. To determine whether 15d-PGJ2 could induce the activation of TFEB, we treated CF-7 cells [6] (HeLa cells stably expressing 3XFlag-TFEB) with different concentrations of 15d-PGJ2 (0–30 µM) for 12 h. Immunostaining results showed that 15d-PGJ2 effectively promoted the translocation of TFEB from the cytoplasm into the nucleus in a dose-dependent manner (Fig. 1a, b). Moreover, we also treated cells with 30 µM 15d-PGJ2 for different durations (0–9 h) and found that 15d-PGJ2 rapidly promoted the nuclear accumulation of TFEB (Fig. 1c, d). As expected, a known mTORC1 inhibitor Torin 1 effectively induced this accumulation of TFEB in the nucleus (Fig. 1). Consistent with immunostaining results, the induction of TFEB accumulation in the nucleus upon 15d-PGJ2 treatment was further confirmed by isolation of cytosolic and nucleus fractions followed by immunoblot assay (Fig. 1e, f). Together, these results suggest that 15d-PGJ2 promotes TFEB nuclear accumulation. Moreover, 15d-PGJ2 also promotes the nuclear translocation of TFEB in HT-22 (mouse hippocampal neuronal cell line), and HEK293 cells (Fig. S1a, b), suggesting that 15d-PGJ2 might promote TFEB translocation into nucleus in multiple types of cells.

a 15d-PGJ2 promotes the accumulation of TFEB in the nucleus in a dose-dependent manner. The cytosol and nucleus distribution of TFEB in HeLa cells stably expressing 3×Flag-TFEB (CF-7) were detected after treating with indicated concentrations of 15d-PGJ2 for 12 h, or a Torin 1 (250 nM positive control) for 3 h. b Quantification of the number of cells with nuclear TFEB localization in a. c 15d-PGJ2 induces the accumulation of TFEB in the nucleus in a time-dependent manner. The cytosol and nucleus distribution of TFEB in CF-7 cells were detected after treating cells with 30 µM 15d-PGJ2 for indicated durations. d Quantification of the number of cells with nuclear TFEB localization in c. e, f The expression of Flag-TFEB in the cytosol (Cyt.) and the nucleus (Nuc.) were detected by Western blotting after being treated with indicated compounds for 3 h. Representative data are shown from three independent experiments. Quantification data are presented as mean ± SEM of three replicates in a representative experiment. At least 200 cells were analyzed in each treatment group. Scale bar 15 µm. *P < 0.05; **P < 0.01.
15d-PGJ2 enhances autophagosomal and lysosome biogenesis
Since TFEB is a master regulator of autophagosomal and lysosome biogenesis, we asked whether 15d-PGJ2-induced the nucleus accumulation of TFEB enhances autophagosomal and lysosomal biogenesis. As shown in Fig. 2a, b, 15d-PGJ2 significantly increased the levels of LC3-II, a well-known autophagy marker [41], in a dose- and time-dependent manner. mTORC1 inhibitor Torin 1 also increased LC3-II levels. To ascertain whether the increased LC3-II was due to the induction of the formation of autophagosomes or inhibition of LC3-II degradation, we found that 15d-PGJ2 further increased LC3-II in the presence of lysosomal inhibitor chloroquine (CQ) [41] (Fig. 2c), indicating that 15d-PGJ2 enhances autophagy rather than blocks lysosomal degradation. The role of 15d-PGJ2 in promoting autophagy flux was further confirmed by the increased red-only puncta (autolysosomes) after treating cells with 15d-PGJ2 in HeLa cells stably expressing mRFP-GFP-LC3 (tfLC3) construct [41] (Fig. 2d). Overall, these results suggest that 15d-PGJ2 enhances autophagy flux. Furthermore, since TFEB upregulates the expression of autophagy-lysosome-related genes to enhance lysosome biogenesis, we first assessed whether 15d-PGJ2, under conditions that result in TFEB accumulation in the nucleus, could induce the expression of autophagy-lysosome-related genes. As shown by q-PCR results, 15d-PGJ2 treatment significantly increased mRNA levels of several autophagy and lysosome-related genes such as MAP1LC3, SQSTM1, ATP6VE1H, CTSD, LAMP1, and SCPEP1. (Fig. 2e). 15d-PGJ2 treatment also enhanced lysosome marker LAMP1 levels (Fig. 2f). Importantly, after staining cells with LysoTracker Red DND99, we found that 15d-PGJ2 treatment increased fluorescent intensity compared with vehicle-treated control cells as reflected by flow cytometry assay (Fig. 2g), indicating that 15d-PGJ2 enhances lysosome contents. Overall, these results indicate that 15d-PGJ2 treatment not only enhances autophagy but also promotes lysosome biogenesis.

a 15d-PGJ2 increases LC3-II levels in a dose-dependent manner. Torin 1 was used as a positive control. b 15d-PGJ2 increases LC3-II levels in a time-dependent manner. c 15d-PGJ2 promotes autophagy flux. d 15d-PGJ2 increases the formation of autolysosomes as evidenced by the increased red-only puncta after 15d-PGJ2 treatment in HeLa cells stably expressing tf-LC3 plasmids. e 15d-PGJ2 increased the expression of multiple autophagy-lysosomal pathway-related genes as detected by real-time PCR. f 15d-PGJ2 increased LAMP1 levels. g 15d-PGJ2 increases lysosome contents. After HeLa cells were treated with indicated concentrations of 15d-PGJ2 for 16 h, cells were loaded with LysoTracker Red DND-99 (75 nm) for 1 h and the intensity was recorded by a flow cytometer. Representative data are shown from three independent experiments. Quantitative data are presented as the mean ± SEM from at least three independent experiments. Scale bar 10 µm. *P < 0.05; **P < 0.01.
ROS is required for 15d-PGJ2-induced TFEB nuclear localization
To understand how 15d-PGJ2 activates TFEB, we first determined whether PPARγ is responsible for TFEB activation in response to 15d-PGJ2 because 15d-PGJ2 is a well-known PPARγ ligand. As shown in Fig. 3a, PPARγ antagonist GW9662 did not block 15d-PGJ2-induced TFEB nuclear localization, suggesting that 15d-PGJ2-induced TFEB nuclear localization may be independent of PPARγ. Next, we determined whether the nuclear localization of TFEB observed in 15d-PGJ2-treated cells was due to the inhibition of mTORC1, a main molecule for regulating TFEB activity. We treated cells with 15d-PGJ2 for different durations, and found that treatment of cells with 15d-PGJ2, under conditions that promoted TFEB accumulation in the nucleus, did not obviously inhibit the phosphorylation of p70S6K and 4EBP1 (Fig. 3b), two known downstream effectors of mTORC1, though 15d-PGJ2 effectively promoted TFEB accumulation in the nucleus. In contrast, Torin 1 induced strong mTORC1 inactivation (Fig. 3b). These results indicate that 15d-PGJ2 treatment-induced TFEB nuclear localization do not require mTORC1 inactivation. Furthermore, 15d-PGJ2 did not inhibit the phosphorylation of TFEB at Ser211 and Ser142 (Fig. 3b), two main phosphorylation residues that control TFEB intracellular localization modulated by mTORC1, further indicating that 15d-PGJ2-induced TFEB nuclear localization is independent of mTORC1 inhibition. It has been reported that calcium signaling-dependent activation of the phosphatase PPP3/calcineurin promoted TFEB dephosphorylation and activation [16]. We noticed this, therefore, we determined whether calcium and PPP3/calcineurin regulate TFEB nuclear localization in response to 15d-PGJ2. As shown in Fig. 3c, pretreatment of CF-7 cells with calcium chelator BAPTA-AM, or a commonly used PPP3/calcineurin inhibitor FK506 did not noticeably alter 15d-PGJ2-dependent TFEB nuclear localization, suggesting that calcium and PPP3/calcineurin are not required for TFEB nuclear localization in response to 15d-PGJ2.

a PPARγ antagonist GW9662 does not block 15d-PGJ2-induced TFEB activation. After pretreatment of CF-7 cells with PPARγ antagonist GW9662 (10 µM) for 30 min followed by adding 15d-PGJ2 into cells for another 12 h, the cytoplasm and the nuclear distribution of TFEB were detected by immunostaining. b 15d-PGJ2 does not inhibit the mTOR signaling pathway. After cells were treated with 15d-PGJ2 (30 µM) for indicated durations, the expression of p-p70S6K /P70S6K, p-4EBP1/4EBP1, p-TFEB142, p-TFEB 211 were detected by Western blotting; Torin 1 (250 nm) was used as a positive control. c Calcium chelator BAPTA-AM or calcineurin inhibitor FK506 does not inhibit 15-dPGJ2-induced TFEB activation. After pretreatment of CF-7 cells with BAPTA-AM (20 µM) or calcineurin inhibitor FK506 (10 µM) for 30 min followed by adding 15d-PGJ2 into cells for another 12 h, the cytoplasm and the nucleus distribution of TFEB were detected by immunostaining. d PP2A inhibitor OA (okadaic acid) does not inhibit 15-dPGJ2-induced TFEB activation. After pretreatment of CF-7 cells with OA (400 nM) for 30 min followed by adding 15d-PGJ2 into cells for another 12 h, the cytoplasm and the nucleus distribution of TFEB was detected by immunostaining. Representative data are shown from three independent experiments. Scale bar 15 µm. ns no significance.
ROS has been reported to promote TFEB nucleus accumulation [9, 42, 43]. Given this observation, we sought to investigate whether ROS contributes to 15d-PGJ2 treatment-induced TFEB nuclear localization. We first measured ROS levels by determining the fluorescence intensity of 2′, 7′ –dichlorofluorescein (DCF) with a flow cytometer. As shown in Fig. 4a, b, 15d-PGJ2 increased ROS levels in a time-dependent manner, which is consistent with several studies showing that 15d-PGJ2 treatment could generate ROS [27, 44]. Intriguingly, ROS scavenger NAC (N-acetylcysteine) not only reduced 15-dPGJ2-induced ROS levels (Fig. 4c), but also effectively attenuated the nuclear localization of TFEB as reflected by immunostaining and Western blotting results (Fig. 4d, e), suggesting that ROS is required for 15d-PGJ2-induced TFEB nuclear localization. Consistent with these observations, NAC obviously inhibited 15d-PGJ2-induced expression of LC3-II (Fig. 4g), the formation of autolysosomes (Fig. 4f, tf-LC3 staining), the increased lysosome contents (Fig. 4h, Lysotracker Red staining), and the upregulated expression of autophagy-lysosomal related genes (Fig. S2). Overall, these results indicate that ROS is necessary for 15d-PGJ2-induced TFEB nuclear localization and the subsequent autophagy and lysosome biogenesis.

a, b 15d-PGJ2 induces the generation of ROS. After treating cells with 15d-PGJ2 for indicated durations and loading cells with DCF-DA (100 μM) for 1 h, the ROS intensity was measured with a flow cytometer and quantified. c ROS scavenger N-acetyl cysteine (NAC) effectively inhibits 15d-PGJ2-induced production of ROS. d, e NAC inhibits 15d-PGJ2-induced TFEB activation. After pretreatment of CF-7 cells with NAC (5 mM) for 30 min followed by adding 15d-PGJ2 into cells for another 12 h, the cytoplasm and the nuclear distribution of TFEB were detected by immunostaining (d) and Western blotting (e). Scale bar 15 µm. f Pretreatment of CF-7 cells with NAC inhibited 15d-PGJ2-induced formation of autolysosomes (red-only puncta) as reflected by tf-LC3 staining and quantification data. Scale bar 10 µm. g Pretreatment of CF-7 cells with NAC inhibited 15d-PGJ2-induced increase of LC3-II as reflected by Western blot assay. h Pretreatment of CF-7 cells with NAC inhibited 15d-PGJ2-induced increase of lysosome contents as reflected by Lysotrack Red staining. Representative data are shown from three independent experiments. *P < 0.05; **P < 0.01.
TFEB promotes 15d-PGJ2-induced cell apoptosis
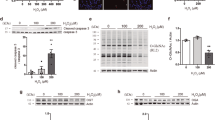
To further assess the physiological significance of TFEB in response to 15d-PGJ2, we treated cells with 15d-PGJ2 and found that 15d-PGJ2 promoted cell apoptosis, which is consistent with previous reports [19, 44]. By using TFEB KO HeLa cells (Fig. 5a), we surprisingly found that TFEB-deficient cells were less susceptible to cell death in response to 15d-PGJ2 as reflected by MTT assay (Fig. 5b). Cell morphology analysis further showed that TFEB-depleted cells were less sensitive to 15d-PGJ2-induced cell death (Fig. 5c). We then performed Annexin V and PI co-staining to determine cell apoptosis. As shown in Fig. 5d, knockout of TFEB reduced cell apoptosis induced by 15d-PGJ2 compared with wild-type cells. The effect of TFEB in promoting apoptosis was further verified by measuring the levels of apoptosis-related proteins Bcl-2, cleaved caspase 3, and cleaved caspase 9 levels in wild-type and TFEB KO cells after treatment with 15d-PGJ2. As shown in Fig. 5e, TFEB KO restored 15d-PGJ2-induced changes of cleaved caspase 3, cleaved caspase 9, and Bcl-2 levels. Finally, we overexpressed TFEB in the TFEB KO cells, and found that reconstitution of TFEB increased cell apoptosis as reflected by increased the expression of cleaved caspase 3 (Fig. S3), cell morphology and Annexin V-FITC staining. Together, these results suggest that TFEB promotes cell apoptosis in response to 15d-PGJ2.

a Western blotting was used to detect the expression of TFEB in WT and TFEB KO cells. b TFEB deficiency increased cell viability (MTT assay) in response to 15d-PGJ2. c TFEB KO cells were less sensitive to 15d-PGJ2-induced cell death as evidenced by phase-contrast imaging results. d TFEB-depleted cells were less susceptible to apoptosis in response to 15d-PGJ2 as reflected by Annexin V-FITC/PI double staining. e TFEB depletion inhibited the levels of cleaved caspase 3 and cleaved caspase 9, while it enhanced the decrease in Bcl-2 levels in response to 15d-PGJ2. Representative data are shown from three independent experiments. *P < 0.05.
TFEB-mediated ATF4 upregulation is responsible for 15d-PGJ2-induced cell apoptosis
Previous studies have shown that prolonged ER stress induces TFEB activation, promoting cell apoptosis by direct transcriptional upregulation of ATF4 (activating transcription factor 4) and other UPR (unfold protein response) genes [7]. To determine how TFEB activation contributes to 15d-PGJ2-induced apoptosis, we first examined whether 15d-PGJ2 could induce the expression of ATF4 and its downstream target CHOP (C/EBP homologous protein). As shown in Fig. 6a, 15d-PGJ2 treatment for 24 h upregulated the expression of ATF4 and CHOP in a dose-dependent manner. In contrast, Torin 1, a well-known mTORC1 inhibitor that effectively promoted TFEB activation, did not obviously increase ATF4 and CHOP levels (Fig. 6a), suggesting that not all TFEB activators could increase ATF4 levels. Interestingly, the increased expression of ATF4 and CHOP in response to 15d-PGJ2 was TFEB-dependent because TFEB deficiency almost completely blocked the upregulation of ATF4 and CHOP in both protein and mRNA levels (Fig. 6b, c). Interestingly, reconstitution of TFEB in the TFEB KO cells increased the expression of ATF4, and CHOP (Fig. S3a). Similarly, siRNA-mediated knockdown of the expression of TFEB also inhibited the upregulation of ATF4 and CHOP (Fig. S4).

a 15d-PGJ2 treatment increased the expression of ATF4 and its downstream target CHOP after 24 h treatment. b TFEB is required for 15d-PGJ2-induced ATF4 upregulation. In WT and TFEB KO cells, after treatment with 30 μM 15d-PGJ2 or vehicle control, the expression of ATF4 and CHOP was determined. c TFEB deletion attenuated 15d-PGJ2-induced expression of ATF4 and CHOP at mRNA levels. d RNAi-mediated knockdown of ATF4 attenuated 15d-PGJ2-induced expression of ATF4, CHOP as well as apoptosis-related proteins cleaved caspase 3 and cleaved caspase 9. e Knockdown of the expression of ATF4 inhibited 15d-PGJ2-induced cell death as evidenced by phase-contrast imaging results. f The effects of ATF4 knockdown on inhibiting 15d-PGJ2-induced apoptosis were further confirmed by Annexin V-FITC/PI double staining. g Protein synthesis inhibitor cycloheximide (CHX, 10 µg/mL) inhibited 15d-PGJ2-induced expression of ATF4 and CHOP. h CHX (10 µg/mL) inhibited apoptosis after 24 h of 15d-PGJ2 treatment as reflected by phase-contrast imaging results. i CHX (10 µg/mL) inhibited 15d-PGJ2-induced upregulation of cleavage caspase 3. j CHX-treated cells were less susceptible to apoptosis in response to 15d-PGJ2 verified by Annexin V-FITC/PI double staining results. Representative data are shown from three independent experiments. *P < 0.05.
ATF4 is a transcriptional factor that modulates both survival and apoptosis in a cell type- or tissue-specific manner and constitutes a response to different stressors [45,46,47,48]. For instance, ATF4 could regulate apoptosis by promoting the expression of CHOP, which is a transcription factor that inhibits the expression of Bcl-2 and induces mitochondria-dependent cell apoptosis [45,46,47,48]. To investigate whether the upregulated ATF4 contributes to cell apoptosis in response to 15d-PGJ2, we firstly knocked down the expression of ATF4 and found that, as expected, ATF4 depletion attenuated 15d-PGJ2-induced expression of CHOP (Fig. 6d). Moreover, ATF4 depletion compromised 15d-PGJ2-induced upregulation of cleaved caspase 3 and cleaved caspase 9 levels (Fig. 6d), suggesting that ATF4 may be required for 15d-PGJ2-induced apoptosis. Furthermore, cell morphology analysis showed that ATF4 depleted cells were less sensitive to 15d-PGJ2-induced cell death (Fig. 6e). Finally, we performed Annexin V and PI co-staining to determine cell apoptosis using a flow cytometer and further confirmed that knockdown of the expression of ATF4 reduced cell apoptosis in response to 15d-PGJ2 (Fig. 6f). These results suggest that the TFEB/ATF4 axis contributes to 15d-PGJ2-induced cell apoptosis.
To further confirm that transcriptional upregulated ATF4/CHOP may be responsible for 15d-PGJ2-induced apoptosis of cells, we treated cells with a de novo protein synthesis inhibitor cycloheximide (CHX). As expect, CHX effectively blocked 15d-PGJ2-induced upregulation of ATF4 and CHOP (Fig. 6g). Moreover, Western blotting results showed that CHX inhibited 15d-PGJ2-induced elevated cleaved caspase 3 levels (Fig. 6i). Additionally, both cell morphology and Annexin V-FITC/PI double staining results demonstrated that CHX-treated cells are less sensitive to cell apoptosis in response to 15d-PGJ2 (Fig. 6h, j). These results indicate that the transcriptionally upregulated expression of ATF4/CHOP may play a role in 15d-PGJ2-induced apoptosis. Altogether, these results demonstrate that TFEB-mediated ATF4 upregulation contributes to 15d-PGJ2-induced cell apoptosis.
Inhibition of TFEB activation by ROS scavenger NAC compromises not only 15d-PGJ2-induced ATF4 upregulation but also cell apoptosis
Since our results show that reduction of ROS levels by NAC (N-acetyl cysteine) is effective in blocking TFEB translocation into the nucleus (Fig. 4d, e), and TFEB-induced ATF4 upregulation is required for apoptosis in response to 15d-PGJ2 (Figs. 5 and 6), we next asked whether ROS plays a role in regulating ATF4 expression and apoptosis upon 15d-PGJ2 treatment. To answer this question, we treated cells with ROS scavenger NAC, and found that, under conditions effectively inhibiting TFEB translocation into the nucleus, NAC blocked 15d-PGJ2-induced upregulation of ATF4 and CHOP (Fig. 7a). In agreement with the result that TFEB and ATF4 are required for 15d-PGJ2-induced apoptosis, NAC was sufficient to inhibit 15d-PGJ2-induced cell death as reflected by MTT assay (Fig. 7b). Consistently, NAC reversed the changes of several apoptotic-related proteins such as cleaved caspase 3, cleaved caspase 9, and Bcl-2 upon 15d-PGJ2 treatment (Fig. 7c). Morphology analysis (Fig. 7d) and Annexin V-FITC/PI double staining (Fig. 7e) results further showed that NAC could inhibit 15d-PGJ2-induced apoptosis. Overall, these results suggest that inhibition of TFEB translocation into the nucleus by ROS scavenger NAC is effective in preventing 15d-PGJ2-induced ATF4/CHOP upregulation and apoptosis.

a ROS scavenger NAC effectively inhibited 15-dPGJ2-induced upregulation of ATF4 and CHOP. After pretreatment of cells with NAC (5 mM) followed by adding 15d-PGJ2 (30 μM) for another 24 h, the expression of ATF4 and CHOP were determined. b MTT assay showed that NAC inhibited 15d-PGJ2-induced cell death. c Western blotting results showed that NAC reversed 15d-PGJ2-induced changes of cleaved caspase 3, cleaved caspase 9, and Bcl-2 levels. d Phase-contrast imaging results showed that NAC inhibited 15d-PGJ2 treatment-induced cell death. e After pretreatment of cells with NAC followed by addition of 30 μM 15d-PGJ2 for another 24 h, Annexin V-FITC/PI double staining results further verified that NAC-treated cells were less susceptible to apoptosis in response to 15d-PGJ2. Representative data are shown from three independent experiments. **P < 0.01.
Discussion
TFEB is a pro-survival factor involved in promoting autophagy-lysosome pathway. Besides, it also exerts other biological functions, including promoting apoptosis trigged by ER stressor. Whether other conditions induced TFEB translocation into the nucleus also promote apoptosis is largely unclear. In this study, we have identified a novel role of ROS-activated TFEB translocation into the nucleus in promoting autophagy and apoptosis in response to the cyclopentenone prostaglandin 15d-PGJ2. The generation of ROS, rather than mTOR inhibition or calcium-dependent calcineurin signaling, is required for TFEB translocation into the nucleus elicited by 15d-PGJ2. Upon translocation into the nucleus, on the one hand, TFEB enhances autophagy and lysosome biogenesis by upregulating the expression of multiple autophagy-lysosome-related genes; on the other hand, TFEB transcriptionally upregulates the expression of ATF4 to promote apoptosis. (Fig. 8). Thus, these results not only reveal how 15d-PGJ2 activates TFEB, but also provide a novel pro-apoptotic effect of TFEB in response to 15d-PGJ2.

ROS production promotes the translocation of TFEB from the cytoplasm into the nucleus in response to 15d-PGJ2. In the nucleus, TFEB not only enhances autophagosomal and lysosomal biogenesis via regulating the expression of multiple autophagosomal-lysosome-related genes, but also triggers apoptosis by upregulating the expression of ATF4 and its target CHOP. Conversely, ROS scavenger N-acetyl cysteine (NAC) blocks 15d-PGJ2-induced TFEB translocation into the nucleus, which attenuates the autophagosomal and lysosomal biogenesis and inhibits apoptosis in response to 15d-PGJ2 treatment.
The activity of TFEB is mainly regulated by its phosphorylation status. mTORC1 is the main kinase responsible for TFEB phosphorylation [13,14,15], and calcineurin is the main phosphatase that de-phosphorylates TFEB [16]. Upon dephosphorylation, TFEB is rapidly translocated into the nucleus to enhance autophagy and lysosome biogenesis [5]. 15d-PGJ2-induced TFEB translocation into the nucleus is probably independent of mTOR inhibition because mTOR phosphorylation status did not correlate with the nuclear accumulation of TFEB in response to 15d-PGJ2 (Fig. 3b). In contrast, 15d-PGJ2-induced ROS production was responsible for TFEB translocation into the nucleus since ROS scavenger NAC almost completely blocked TFEB translocation in response to 15d-PGJ2 (Fig. 4d). ROS was reported to activate TFEB via three different mechanisms, which are ROS-mediated PP2A activation to dephosphorylate TFEB [43], ROS-mediated calcineurin activation to dephosphorylate TFEB [42], and ROS-mediated direct oxidation of cysteine residue of TFEB [9]. Interestingly, ROS (H2O2)-mediated TFEB activation is via direct cysteine oxidation and independent of its dephosphorylation [9]. Our results showed that PP2A inhibitor OA (okadaic acid) (Fig. 3d), calcium chelator and calcineurin inhibitor FK506 (Fig. 3c) did not inhibit TFEB translocation into the nucleus upon 15d-PGJ2 treatment, indicating that 15d-PGJ2-induced ROS generation may lead to direct oxidation and subsequent activation of TFEB. This hypothesis was further verified by the results that 15d-PGJ2 did not obviously induce the dephosphorylation of TFEB (Fig. 3b). However, it remains unclear how 15d-PGJ2 promotes the formation of ROS, though we found that the ROS scavenger NAC was effective in inhibition of TFEB translocation into the nucleus. Notably, though we observed that a short (e.g. less than 6 h) 15d-PGJ2 treatment, under conditions that effectively promoted TFEB nucleus accumulation, did not obviously inhibit mTORC1. However, prolonged 15d-PGJ2 treatment may inhibit mTORC1 via ROS production and ATF4 signaling. Since it has been reported that ROS inhibits mTORC1 via inhibiting Akt signaling [49] and ATF4 inhibits mTORC1 via upregulating REDD1 (regulated in development and DNA damage responses 1) [50] and Sestrin 2 [51]. Inactivated mTORC1 then further de-phosphorylates and activates TFEB; thus TFEB-activated ATF4 may further enhance TFEB signaling through a positive feedback mechanism in response to 15d-PGJ2. Overall, these results establish an important link between the endogenous prostaglandin 15d-PGJ2-induced ROS and TFEB translocation into the nucleus.
Since TFEB plays a major role in promoting cell survival by upregulating autophagosomal and lysosome biogenesis [1, 5, 40], we initially assumed that 15d-PGJ2-induced TFEB translocation into the nucleus was a cytoprotective response and may inhibit apoptosis. Unexpectedly, TFEB KO cells were less susceptible to apoptosis in response to 15d-PGJ2 (Fig. 5), suggesting that 15d-PGJ2-induced TFEB translocation into the nucleus promotes apoptosis. To determine how TFEB contributes to apoptosis in response to 15d-PGJ2, we focused on ATF4 as a previously study showed that ATF4 is a directly target of TFEB in response to ER stressor tunicamycin [7]. ATF4 is a transcription factor that is constitutively expressed only at low levels. In contrast, in response to cell stress it can be rapidly induced [45]. For instance, upon ER stress, accumulated misfolded proteins in the ER lumen induce UPR to restore cellular homeostasis [45, 47, 48]. The activated UPR branch PERK then phosphorylates eIF2α, resulting in inhibition of general protein translation but selective promotion of the translation of several genes including ATF4 [47, 48]. ATF4 can modulate both survival and apoptosis in a condition-specific manner [45, 47, 48]. It promotes cell survival by upregulating the expression of multiple genes involved in autophagy, amino acid metabolism, and redox homeostasis [45, 47, 48]. However, ATF4 also enhances cell apoptosis by inducing the expression of CHOP [47, 48, 52], which modulates the expression of several apoptotic-proteins, including Bcl-2 family members, to induce mitochondria-dependent apoptotic pathway [47, 48, 52]. Here, we reveal that 15d-PGJ2 upregulates the expression of ATF4/CHOP in an TFEB-dependent manner in response to 15d-PGJ2 and knockdown of the expression of ATF4 prevented 15d-PGJ2-induced apoptosis. Accordingly, in both TFEB KO and ATF4 knockdown cells, upon 15d-PGJ2 treatment, the levels of CHOP and cleaved caspase 3 were reduced and antiapoptotic protein Bcl-2 was increased (Fig. 6). In agreement with these findings, inhibiting TFEB translocation into the nucleus by ROS scavenger NAC or inhibition of protein synthesis by CHX not only attenuated the upregulation of ATF4 and CHOP but also inhibited apoptosis in response to 15d-PGJ2. Therefore, these observations demonstrate that TFEB facilitates apoptosis by promoting ATF4/CHOP expression in response to 15d-PGJ2 treatment.
Notably, because TFEB and TFE3 have similar functions and share common mechanisms for activation, though our results clearly demonstrated that knockout of TFEB almost completely inhibited the increased ATF4 in response to 15d-PGJ2 (Fig. 6b), it is still possible that 15d-PGJ2 also promotes TFE3 translocation into the nucleus, which may play a role in regulating ATF4 expression and apoptosis in response to 15d-PGJ2. Interestingly, a previously study showed that ER stressors tunicamycin activated TFEB [7], and 15d-PGJ2 was reported to induce ER stress [53]. However, 15d-PGJ2-induced TFEB translocation into the nucleus is independent of its role in inducing ER stress since pretreatment of cells with ER stress inhibitor 4-PBA (4-phenylbutyric acid) did not block TFEB translocation into the nucleus in response to 15d-PGJ2 (Fig. S5). Moreover, ER stressor tunicamycin activated TFEB via calcineurin signaling, which is different from the mechanism of 15d-PGJ2 in inducing TFEB activation since calcineurin inhibitor did not attenuate 15d-PGJ2-induced TFEB activation. Altogether, these results indicate that the underlying mechanism of TFEB translocation into the nucleus in response to 15d-PGJ2 and ER stressor are different, though both of them upregulate the expression of ATF4 in TFEB dependent manner [7].
15d-PGJ2, the first identified endogenous PPARγ ligand, has received extensive attention because it functions as a potential regulator of multiple biological processes [24]. However, increasing evidence has shown that some physiological functions of this small molecule are independent on PPARγ [24]. In agreement with a previous finding that 15d-PGJ2 induces apoptosis via ROS formation rather than PPARγ activation [29, 44], 15d-PGJ2-induced TFEB activation is also independent of PPARγ (Fig. 3a). ROS was reported to initiate mitochondria-mediated apoptosis by inhibiting Akt and promoting the JNK pathway [45]. There are also other targets of 15d-PGJ2 such as androgen receptor and glucocorticoid receptor [54, 55]. Here, we showed that though 15d-PGJ2-induced autophagy and lysosome biogenesis are via ROS-mediated TFEB activation, which promotes rather than inhibits cell apoptosis, suggesting that TFEB/ATF4 are critical links between 15d-PGJ2-mediated ROS and apoptosis.
Collectively, our study unveils a previously unexplored role of ROS-mediated TFEB translocation into the nucleus that coordinates both 15d-PGJ2-induced autophagy and apoptosis in response to 15d-PGJ2 treatment, providing novel insight into how small molecules in promoting TFEB translocation into the nucleus, a master regulator of autophagy and lysosome biogenesis, to modulate cell fitness. Our results extend the importance of ROS-activated TFEB translocation in enhancing rather than inhibiting autophagy in response to 15d-PGJ2. Given the therapeutic potential of small molecule modulators of TFEB in treating multiple diseases including neurodegenerative diseases and cancers [40], the identification of novel roles of small molecule TFEB activators are critically important.
References
Dikic I, Elazar Z. Mechanism and medical implications of mammalian autophagy. Nat Rev Mol Cell Biol. 2018;19:349–64.
Boya P, Reggiori F, Codogno P. Emerging regulation and functions of autophagy. Nat Cell Biol. 2013;15:713–20.
Yang C, Zhang W, Dong X, Fu C, Yuan J, Xu M, et al. A natural product solution to aging and aging-associated diseases. Pharmacol Ther. 2020;216:107673.
Cai CZ, Yang C, Zhuang XX, Yuan NN, Wu MY, Tan JQ, et al. NRBF2 is a RAB7 effector required for autophagosome maturation and mediates the association of APP-CTFs with active form of RAB7 for degradation. Autophagy. 2021;17:1112–30.
Napolitano G, Ballabio A. TFEB at a glance. J Cell Sci. 2016;129:2475–81.
Settembre C, Di Malta C, Polito VA, Garcia Arencibia M, Vetrini F, Erdin S, et al. TFEB links autophagy to lysosomal biogenesis. Science. 2011;332:1429–33.
Martina JA, Diab HI, Brady OA, Puertollano R. TFEB and TFE3 are novel components of the integrated stress response. EMBO J. 2016;35:479–95.
Brady OA, Jeong E, Martina JA, Pirooznia M, Tunc I, Puertollano R. The transcription factors TFE3 and TFEB amplify p53 dependent transcriptional programs in response to DNA damage. eLife. 2018;7:e40856.
Wang R, Tan J, Chen T, Han H, Tian R, Tan Y, et al. ATP13A2 facilitates HDAC6 recruitment to lysosome to promote autophagosome-lysosome fusion. J Cell Biol. 2019;218:267–84.
Nezich CL, Wang C, Fogel AI, Youle RJ. MiT/TFE transcription factors are activated during mitophagy downstream of Parkin and Atg5. J Cell Biol. 2015;210:435–50.
Brady OA, Martina JA, Puertollano R. Emerging roles for TFEB in the immune response and inflammation. Autophagy. 2018;14:181–9.
Erlich AT, Brownlee DM, Beyfuss K, Hood DA. Exercise induces TFEB expression and activity in skeletal muscle in a PGC-1α-dependent manner. Am J Physiol Cell Physiol. 2018;314:C62–c72.
Settembre C, Zoncu R, Medina DL, Vetrini F, Erdin S, Erdin S, et al. A lysosome-to-nucleus signalling mechanism senses and regulates the lysosome via mTOR and TFEB. EMBO J. 2012;31:1095–108.
Roczniak-Ferguson A, Petit CS, Froehlich F, Qian S, Ky J, Angarola B, et al. The transcription factor TFEB links mTORC1 signaling to transcriptional control of lysosome homeostasis. Sci Signal. 2012;5:ra42.
Martina JA, Chen Y, Gucek M, Puertollano R. MTORC1 functions as a transcriptional regulator of autophagy by preventing nuclear transport of TFEB. Autophagy. 2012;8:903–14.
Medina DL, Di Paola S, Peluso I, Armani A, De Stefani D, Venditti R, et al. Lysosomal calcium signalling regulates autophagy through calcineurin and TFEB. Nat Cell Biol. 2015;17:288–99.
Yang C, Zhu Z, Tong BC, Iyaswamy A, Xie WJ, Zhu Y, et al. A stress response p38 MAP kinase inhibitor SB202190 promoted TFEB/TFE3-dependent autophagy and lysosomal biogenesis independent of p38. Redox Biol. 2020;32:101445.
Puertollano R, Ferguson SM, Brugarolas J, Ballabio A. The complex relationship between TFEB transcription factor phosphorylation and subcellular localization. EMBO J. 2018;37:e98804.
Straus DS, Glass CK. Cyclopentenone prostaglandins: new insights on biological activities and cellular targets. Med Res Rev. 2001;21:185–210.
Forman BM, Tontonoz P, Chen J, Brun RP, Spiegelman BM, Evans RM. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell. 1995;83:803–12.
Kliewer SA, Lenhard JM, Willson TM, Patel I, Morris DC, Lehmann JM. A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentiation. Cell. 1995;83:813–9.
Janani C, Ranjitha Kumari BD. PPAR gamma gene–a review. Diabetes Metab Syndr. 2015;9:46–50.
Reginato MJ, Krakow SL, Bailey ST, Lazar MA. Prostaglandins promote and block adipogenesis through opposing effects on peroxisome proliferator-activated receptor gamma. J Biol Chem. 1998;273:1855–8.
Li J, Guo C, Wu J. 15-Deoxy-∆-(12,14)-prostaglandin J2 (15d-PGJ2), an endogenous ligand of PPAR-γ: function and mechanism. PPAR Res. 2019;2019:7242030.
Kansanen E, Kivelä AM, Levonen AL. Regulation of Nrf2-dependent gene expression by 15-deoxy-Delta12,14-prostaglandin J2. Free Radic Biol Med. 2009;47:1310–7.
Landar A, Shiva S, Levonen AL, Oh JY, Zaragoza C, Johnson MS, et al. Induction of the permeability transition and cytochrome c release by 15-deoxy-Delta12,14-prostaglandin J2 in mitochondria. Biochem J. 2006;394:185–95.
Muri J, Feng Q, Wolleb H, Shamshiev A, Ebner C, Tortola L, et al. Cyclopentenone prostaglandins and structurally related oxidized lipid species instigate and share distinct pro- and anti-inflammatory pathways. Cell Rep. 2020;30:4399–417. e7
Toaldo C, Pizzimenti S, Cerbone A, Pettazzoni P, Menegatti E, Daniela B, et al. PPARgamma ligands inhibit telomerase activity and hTERT expression through modulation of the Myc/Mad/Max network in colon cancer cells. J Cell Mol Med. 2010;14:1347–57.
Shin SW, Seo CY, Han H, Han JY, Jeong JS, Kwak JY, et al. 15d-PGJ2 induces apoptosis by reactive oxygen species-mediated inactivation of Akt in leukemia and colorectal cancer cells and shows in vivo antitumor activity. Clin Cancer Res. 2009;15:5414–25.
Ceaser EK, Ramachandran A, Levonen AL, Darley-Usmar VM. Oxidized low-density lipoprotein and 15-deoxy-delta 12,14-PGJ2 increase mitochondrial complex I activity in endothelial cells. Am J Physiol Heart Circ Physiol. 2003;285:H2298–308.
Diers AR, Higdon AN, Ricart KC, Johnson MS, Agarwal A, Kalyanaraman B, et al. Mitochondrial targeting of the electrophilic lipid 15-deoxy-Delta12,14-prostaglandin J2 increases apoptotic efficacy via redox cell signalling mechanisms. Biochem J. 2010;426:31–41.
Tae IH, Park EY, Dey P, Son JY, Lee SY, Jung JH, et al. Novel SIRT1 inhibitor 15-deoxy-Δ12,14-prostaglandin J2 and its derivatives exhibit anticancer activity through apoptotic or autophagic cell death pathways in SKOV3 cells. Int J Oncol. 2018;53:2518–30.
Chen K, Li J, Li S, Feng J, Wu L, Liu T, et al. 15d-PGJ2 alleviates ConA-induced acute liver injury in mice by up-regulating HO-1 and reducing hepatic cell autophagy. Biomed Pharmacother. 2016;80:183–92.
Qin H, Tan W, Zhang Z, Bao L, Shen H, Wang F, et al. 15d-prostaglandin J2 protects cortical neurons against oxygen-glucose deprivation/reoxygenation injury: involvement of inhibiting autophagy through upregulation of Bcl-2. Cell Mol Neurobiol. 2015;35:303–12.
Xu F, Li J, Ni W, Shen YW, Zhang XP. Peroxisome proliferator-activated receptor-γ agonist 15d-prostaglandin J2 mediates neuronal autophagy after cerebral ischemia-reperfusion injury. PLoS One. 2013;8:e55080.
Settembre C, Zoncu R, Medina DL, Vetrini F, Erdin S, Erdin S, et al. A lysosome‐to‐nucleus signalling mechanism senses and regulates the lysosome via mTOR and TFEB. EMBO J. 2012;31:1095–108.
Song JX, Sun YR, Peluso I, Zeng Y, Yu X, Lu JH, et al. A novel curcumin analog binds to and activates TFEB in vitro and in vivo independent of MTOR inhibition. Autophagy. 2016;12:1372–89.
Kim JW, Li MH, Jang JH, Na HK, Song NY, Lee C, et al. 15-Deoxy-Δ12, 14-prostaglandin J2 rescues PC12 cells from H2O2-induced apoptosis through Nrf2-mediated upregulation of heme oxygenase-1: potential roles of Akt and ERK1/2. Biochem Pharmacol. 2008;76:1577–89.
Yang CB, Pei WJ, Zhao J, Cheng YY, Zheng XH, Rong JH. Bornyl caffeate induces apoptosis in human breast cancer MCF-7 cells via the ROS- and JNK-mediated pathways. Acta Pharmacol Sin. 2014;35:113–23.
Raben N, Puertollano R. TFEB and TFE3: linking lysosomes to cellular adaptation to stress. Annu Rev Cell Dev Biol. 2016;32:255–78.
Klionsky DJ, Abdelmohsen K, Abe A, Abedin MJ, Abeliovich H, Acevedo Arozena A, et al. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy. 2016;12:1–222.
Zhang X, Cheng X, Yu L, Yang J, Calvo R, Patnaik S, et al. MCOLN1 is a ROS sensor in lysosomes that regulates autophagy. Nat Commun. 2016;7:12109.
Martina JA, Puertollano R. Protein phosphatase 2A stimulates activation of TFEB and TFE3 transcription factors in response to oxidative stress. J Biol Chem. 2018;293:12525–34.
Wang JJ, Mak OT. Induction of apoptosis by 15d-PGJ2 via ROS formation: an alternative pathway without PPARγ activation in non-small cell lung carcinoma A549 cells. Prostaglandins Other Lipid Mediat. 2011;94:104–11.
Wortel IMN, van der Meer LT, Kilberg MS, van Leeuwen FN. Surviving stress: modulation of ATF4-mediated stress responses in normal and malignant cells. Trends Endocrinol Metab. 2017;28:794–806.
Pakos-Zebrucka K, Koryga I, Mnich K, Ljujic M, Samali A, Gorman AM. The integrated stress response. EMBO Rep. 2016;17:1374–95.
Frakes AE, Dillin A. The UPR(ER): sensor and coordinator of organismal homeostasis. Mol Cell. 2017;66:761–71.
Oakes SA, Papa FR. The role of endoplasmic reticulum stress in human pathology. Annu Rev Pathol. 2015;10:173–94.
Zhao Y, Hu X, Liu Y, Dong S, Wen Z, He W, et al. ROS signaling under metabolic stress: cross-talk between AMPK and AKT pathway. Mol Cancer. 2017;16:79.
Brüning A, Rahmeh M, Friese K. Nelfinavir and bortezomib inhibit mTOR activity via ATF4-mediated sestrin-2 regulation. Mol Oncol. 2013;7:1012–8.
Kim HJ, Joe Y, Kim SK, Park SU, Park J, Chen Y, et al. Carbon monoxide protects against hepatic steatosis in mice by inducing sestrin-2 via the PERK-eIF2α-ATF4 pathway. Free Radic Biol Med. 2017;110:81–91.
Matsumoto H, Miyazaki S, Matsuyama S, Takeda M, Kawano M, Nakagawa H, et al. Selection of autophagy or apoptosis in cells exposed to ER-stress depends on ATF4 expression pattern with or without CHOP expression. Biol Open. 2013;2:1084–90.
Sperandio M, Demasi APD, Martinez EF, Saad SO, Pericole FV, Vieira KP, et al. 15d-PGJ(2) as an endoplasmic reticulum stress manipulator in multiple myeloma in vitro and in vivo. Exp Mol Pathol. 2017;102:434–45.
Paakinaho V, Kaikkonen S, Levonen AL, Palvimo JJ. Electrophilic lipid mediator 15-deoxy-Δ12,14-prostaglandin j2 modifies glucocorticoid signaling via receptor SUMOylation. Mol Cell Biol. 2014;34:3202–13.
Kaikkonen S, Paakinaho V, Sutinen P, Levonen AL, Palvimo JJ. Prostaglandin 15d-PGJ(2) inhibits androgen receptor signaling in prostate cancer cells. Mol Endocrinol. 2013;27:212–23.
Acknowledgements
We would like to thank Prof. Richard J. Youle (US National Institute of Neurological Disorders and Stroke) and Prof. Myung-Shik Lee (Yonsei University College of Medicine) for providing TFEB knockout HeLa cells. We thank Martha Dahlen for the English editing. This study was supported by the Hong Kong General Research Fund (GRF/HKBU12101417, GRF/HKBU12100618), the National Natural Science Foundation of China (81703487, and 81773926), Shenzhen Science and Technology Innovation Commission (JCYJ20180302174028790, JCYJ20180507184656626), the Hong Kong Health and Medical Research Fund (HMRF17182541, HMRF17182551), and research funds from Hong Kong Baptist University (HKBU/RC-IRCs/17-18/03).
Author information
Authors and Affiliations
Contributions
ML and CBY designed research; JL and CBY performed research, analyzed data; CBY draft the manuscript; BCKT, ZYW, ZZ, CFS, SGS, JXW, AI, KS, SYH, KHC, JXS, JQT, and JHL helped organize the experiment and provided the technical support; CBY, JL, JXS, and ML, reviewed and edited the paper; ML administrated the project. All authors have given approval to the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Supplementary information
Rights and permissions
About this article

Cite this article
Yang, Cb., Liu, J., Tong, B.CK. et al. TFEB, a master regulator of autophagy and biogenesis, unexpectedly promotes apoptosis in response to the cyclopentenone prostaglandin 15d-PGJ2. Acta Pharmacol Sin 43, 1251–1263 (2022). https://doi.org/10.1038/s41401-021-00711-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41401-021-00711-7
Keywords
This article is cited by
-
PPARɑ Ligand Caudatin Improves Cognitive Functions and Mitigates Alzheimer’s Disease Defects By Inducing Autophagy in Mice Models
Journal of Neuroimmune Pharmacology (2023)
