
Abstract
Bipolar disorder (BD) is a complex psychiatric disorder characterized by dysfunctions in three domains including emotional processing, cognitive processing, and psychomotor dimensions. However, the neural underpinnings underlying these clinical profiles are not well understood. Based on the reported data, we hypothesized that (i) the core neuropathology in BD is damage in fronto-limbic network, which is associated with emotional dysfunction; (ii) changes in intrinsic brain network, such as sensorimotor network, salience network, default-mode network, central executive network are associated with impaired cognition function; and (iii) beyond the dopaminergic-driven basal ganglia-thalamo-cortical motor circuit modulated by other neurotransmitter systems, such as serotonin (subcortical–cortical modulation), the sensorimotor network and related motor function modulated by other non-motor networks such as the default-mode network are involved in psychomotor function. In this review, we propose a neurocircuit-based clinical characteristics and taxonomy to guide the treatment of BD. We draw on findings from neuropsychological and neuroimaging studies in BD and link variations in these clinical profiles to underlying neurocircuit dysfunctions. We consider pharmacological, psychotherapy, and neuromodulatory treatments that could target those specific neurocircuit dysfunctions in BD. Finally, it is suggested that the methods of testing the neurocircuit-based taxonomy and important limitations to this approach should be considered in future.
Similar content being viewed by others
Introduction
Bipolar disorder (BD) is a complex psychiatric disorder characterized by severe mood fluctuations associated with dysfunctional cognition that often persist throughout the entire life of patients and abnormal psychomotor activity [1, 2]. The heterogeneous phenomenological dimensions of the various bipolar symptoms can be systematized and traced back to combined changes of affectivity, cognition, and psychomotricity. With the advances in neuroimaging technology, there are increasing opportunities to understand the causes, mechanisms, and treatment of BD. With mounting evidence suggesting that the abnormalities between cortical and subcortical regions associated with emotion, cognition, and psychomotor activity are likely risk markers for BD [3,4,5].
BD has traditionally been associated with dysfunction in the fronto-limbic neural circuits underlying emotional regulation, as well as hypersensitivity in the fronto-striatal neural circuits responsible for rewarding processing, although it is now recognized that alterations in distinct large-scale networks seems to play a fundamental role in the phenomenological dimensions, including affective, cognition, and motor functions [6,7,8,9,10]. Most of neurobiological models of BD suggest a dysfunction of the dorsal and ventral systems, composed, respectively, of (i) the prefrontal cortex (PFC), anterior cingulate cortex (ACC) and the hippocampus and (ii) the insula, amygdala, and ventral striatum [6, 11]. The emotional regulation and mood liability could be due to alterations in these two partially overlapping ventral prefrontal systems, which may result in a loss of homeostasis in emotional processing [12].
Functional neuroimaging research has revealed intrinsic brain activities, the functional architecture as organized in distinct brain networks in accordance to connectivity patterns, and related setting of input/output processing involve in the psychopathology of affectivity, psychomotricity, and thought in BD [13]. Within the last decade, evidence from neuroimaging has highlighted four related largescale intrinsic brain networks that are implicated in a range of cognitive processes and are considered to be domain-general functional networks that support many cognitive function, i.e. the default mode network (DMN), the salience network (SN), the cognitive control network (CEN), and the sensorimotor network (SMN) [14,15,16,17,18]. Abnormalities in these aspects likely are associated with different neurocircuits that interact with each other to generate the complex BD phenotype.
The expression of psychomotor dimensions of BD is traceable to modulation of motor cortex and motor network by non-motor cortical networks, i.e., default-mode network and sensory networks. Additionally, the dopamine-driven basal ganglia-thalamo-cortical motor circuit modulated by other monoaminergic neurotransmitter systems such as serotonin (subcortical–cortical modulation) is also involved in psychomotor mechanisms [19]. Based on these three dimensions that are constructs derived from fundamental phenomenological components, which can be linked to neurobiological systems and fostered by the recent advancement of sophisticated neuroimaging techniques, recent neuroimaging studies of BD demonstrate abnormalities in neural circuits supporting emotional response and regulation, and cognition processing, and producing and controlling motor behavior and the integration of sensory information. Therapeutic approaches based on the patterns of alterations in these neurocircuits and clinical factors varying across individuals could improve treatment efficacy and guide clinical practice. However, it remains unclear how these different clinical profiles are associated with specific pathophysiological mechanisms involved, in particularly the brain regions and networks that ultimately support these functions or their usefulness in guiding treatment selection.
In this review, we enlarge on previous models of the neurocircuitry of BD to propose a heuristic neurocircuit-based taxonomy, which could be used to guide the diagnosis and treatment of the disorder in future. We present hypothetical links between underlying neurocircuit dysfunctions and the three domains of clinical presentation in BD and discuss how treatments that more specifically target these circuits may benefit patients and improve therapeutic response. We suggest ways in which this neurocircuit-based taxonomy could be tested in future research to support or refute our proposed links between aspects of clinical phenomenology across different mood states, specific neurocircuit alterations, and treatment methods. Likewise, the neurocircuits are not as segregated as they may appear in the following sections and highly interactive, and the neurocircuits are certainly more complex and not limited to the neurocognitive and behavioral alterations we describe here.
Emotional regulation and processing dysfunctions of neural network in BD
Emotional hyper-reactivity and mood instability have a detrimental impact on functioning, relapses, and suicide attempts in BD patients. Excessive reactivity to negative events might be a core dimension of BD and could represent an endophenotype during both manic and mixed states in comparison to patients in remitted phases [7, 20]. Research in the field of affective neuroscience conducting neuroimaging studies sheds light on correlations between activities in different brain regions, and the findings can provide insights on the role of different regions in emotion regulation. In the past few decades, the increasing neuroimaging studies have demonstrated that the frontal-limbic connectivity abnormally is a promising biomarker of the pathophysiology and maintenance of BD, which can be considered as the core of affectivity [12, 21].
We will first provide an overview of the prefrontal cortical regions including the anatomy and the fronto-limbic functional connectivity that have been implicated in emotional processing. The functional localization and division of the human prefrontal cortex are orbitofrontal cortex (OFC), dorsomedial prefrontal cortex (dmPFC), dorsolateral prefrontal cortex (dlPFC), ventrolateral prefrontal cortex (vlPFC), and ACC [22]. The ventromedial prefrontal cortex (vmPFC) which develop relatively early comprises OFC, dmPFC, and ACC and are involved especially in the control of emotional behaviors, whereas the lateral prefrontal cortical regions (dlPFC and vlPFC) develop relatively late and are involved in higher executive functions [23]. The former neural system may be involved in automatic subprocesses, whereas the latter neural system may subserve voluntary subprocesses (Fig. 1). The combination of the functional and structural abnormalities in neural systems implicated in voluntary and automatic subprocesses of emotion regulation may underlie the mood instability of BD [24]. Those regions are the most densely connected with the amygdala and other subcortical limbic and paralimbic regions [25].

The lateral prefrontal cortical regions (dlPFC and vlPFC), which develop relatively late and are involved in higher executive functions may subserve voluntary subprocesses. The automatic subprocess includes the area of dmPFC, vmPFC, OFC, ACC, and are involved especially in the control of emotional behaviors. dlPFC dorsolateral prefrontal cortex, vlPFC ventrolateral prefrontal cortex, dmPFC dorsomedial prefrontal cortex, vmPFC ventromedial prefrontal cortex, OFC orbitofrontal cortex, ACC anterior cingulate cortex.
The fronto-limbic circuit includes subcortical and cortical brain regions involved in generating emotional responses (amygdala and nucleus accumbens) and evaluating whether those responses are appropriate or require regulation (vmPFC) [26]. The vmPFC regions relate to each other to form a network structurally and functionally that generates emotional responses and evaluates whether those responses are appropriate or require regulation (vmPFC). The ventral affective circuit is also connected with the hippocampus and the dorsal cognitive circuits, which includes dlPFC, dmPFC, dorsal caudate, and thalamus, and is involved in executive functions (e.g., working memory, planning) as well as emotional regulation (Fig. 2).

The ventral affective circuit includes the orbitofrontal cortex (OFC), ventral striatum (particularly the nucleus accumbens (NAcc)), and the thalamus. This circuit is crucially involved in reward functions, which are largely mediated by dopaminergic signaling. The fronto-limbic circuit mainly includes the prefrontal cortex and amygdala, and hippocampus. These brain regions are structurally and functionally connected with each other to form a network that generates emotional responses and evaluates whether those responses are appropriate or require regulation. vlPFC ventrolateral prefrontal cortex, Amy amygdala, ACC anterior cingulate cortex. dmPFC dorsomedial prefrontal cortex, vmPFC ventromedial prefrontal cortex, OFC orbitofrontal cortex, ACC anterior cingulate cortex. dlPFC dorsolateral prefrontal cortex, vlPFC ventrolateral prefrontal cortex, dCaud dorsal caudate, tham thalamus, NAcc accumbens, Amy amygdala.
It is hypothesized that the abnormal emotional ventral system and hypoactive dorsal system result in a loss of homeostasis in emotional processing, which may contribute to mood liability in BD disorder. This ventral network is involved in identifying salient emotional stimuli and in mediating autonomic responses; while the dorsal system is usually involved in selective attention, planning and explicit regulation of emotional states, with its hypoactivation being underpinning of the unstable cognitive control of emotions and affect in BD [27]. The deficits of emotional processing and regulation in BD involve decreased activity of vlPFC and increased acvity of amygdala, striatal, and mPFC [28, 29]. These abnormalities may be predominately within the left vmPFC regions implicated in automatic emotion regulation, and neuroimaging studies have demonstrated greater subcortical limbic activity (including amygdala, ventral striatum, and hippocampus) to emotional stimuli in adult BD during mania, depression and euthymic, compared to healthy individuals. Patients with BD showed hypoactivation of vlPFC, which also persist in euthymia and might represent a trait feature of BD [30]. More recent work suggests that BD patients show reduced connectivity between the vlPFC, ACC, OFC, and limbic areas across mood states [31,32,33].
Moreover, dysregulation of the amygdala by PFC seems to be crucial to better understand the pathophysiology of the disease. The amygdala is an evolutionarily ancient critical area within the fronto-limbic system that is responsible for flight/fight responses to threats, and is hyperactive in mania, whereas contrasting results have been found during depression [34]. Systematic review on functional connectivity between the amygdala and its projecting areas highlights the essential role of the amygdala in attributing the stimuli’s valence, influencing emotional, and behavioral responses [35]. Several investigators observed increased left amygdala activation in the manic group during facial affect matching, while other groups reported similar findings in BD individuals during depression and remission and with other tasks. Table 1 for a summary of the fronto-limbic system results of functional magnetic resonance imaging emotion studies in bipolar.
Treatments targeting dysfunctional emotion regulation and processing in BD
Most of the treatment regimens of BD owe their discovery to combination of fortune, intellectual rigor, and the careful observation of human and animal phenotypes [36]. The understandings of neuroanatomical basis and pathophysiological underpinnings of BD are increasingly reflected in the treatments moving into clinical trial. Pharmacotherapy, cognitive-behavior intervention, and neuromodulation have been applied across all phases of the bipolar illness and are considered as evidence-based treatments for BD.
Current pharmacotherapy for BD is based on administration of mood stabilizers (lithium, valproate, carbamazepine, and lamotrigine) and increasingly, on the administration of second-generation antipsychotics. It is worth mentioning that lithium appeared to induce a rapid suppression of amygdala over-activation and to show substantial treatment response in BD. Results from neuroimaging studies in patients with BD treated with lithium support the hypothesis that enhancement of neuroplasticity is a potential mechanism underlying the therapeutic effects of lithium. Such studies have also revealed brain changes in both gray and white matter, especially in hippocampal volume [37, 38].
Currently, an increasing number of meta-analyses have evaluated the efficacy of cognitive-behavioral therapy (CBT) for BD [39,40,41]. CBT is based on the hypothesis that BD involves dysregulated cognitive reappraisal, which involves reconceptualization of the emotion-eliciting situations in a way that will decrease emotional intensity. The cognitive reappraisal aspect of CBT engages dorsal cognitive circuitry, i.e., the top-down dorsal prefrontal regions to downregulate overactivated fronto-limbic activity, which may benefit individuals with impaired emotion regulation (dorsal cognitive circuit dysfunction) [26]. CBT seems to be mediated by modulation of the amygdala-dlPFC connectivity, and these findings lead to the hypothesis that the disturbance in effective connectivity from the dlPFC to the amygdala while reappraising in BD patients is due to insufficient prefrontal control. CBT, although not explicitly addressed in treatment, has been shown to reduce emotional symptomatology [42].
Neuromodulation holds promise for a vast number of medically refractory neurological and psychiatric disorders and has been guided by advances in neuroimaging and neurophysiology, which had led to new strategies for better targeting, identification of optimal stimulation sites, and understanding of its therapeutic mechanisms. Repetitive transcranial magnetic stimulation (TMS) is a dominant noninvasive stimulation treatment that delivers short magnetic pulses via an electromagnetic coil and is operated to transiently stimulate or inhibit specific brain regions. TMS has had a growing impact on the treatment of unipolar depression and has already been approved by the FDA for use in treatment-resistant depression [43]. Although there is no clear evidence available to base upon for using TMS in treatment of BP, but it is readily advantageous in similarly leveraging for use in treatment-resistant bipolar depression.
A recent review on TMS for BD suggests the potential of repetitive TMS for reducing depressive symptoms [44]. It has been shown that the TMS protocol over the right dlPFC yields a particular benefit for mania, while over the left dlPFC leads to improvement in depressive symptoms [45]. The first meta-analysis of 19 randomized controlled trials of TMS in the treatment of acute bipolar depression in 181 patients suggests that TMS is efficacious, with overall treatment efficacy in BD comparable with in major depression disorder (MDD). Furthermore, the TMS treatment is safe with regard to treatment-emergent affective switches [46]. A preliminary study also showed that deep TMS is a potentially effective and well-tolerated add-on for pharmacotherapy in resistant bipolar depressed patients [47, 48]. The reviewed evidence collectively suggests the potential of TMS in treating BD in both depression and mania phase. However, it should be noted that there is no adequately powered trial conducted yet to further support the efficacy of TMS in BD. When compared with sham treatments, most RCTs found no significant differences in depression symptoms [49]. Paul et al. examined the effectiveness of sequential (to the right dlPFC and then the left dlPFC) bilateral repetitive transcranial magnetic stimulation in the treatment of bipolar depression and found there was no significant difference in mean reduction in depression rating scale scores or response rates between active and sham stimulation [50]. The clear diagnosis of bipolar in the depressive phase, a stable platform of medications (we suggest mood stabilizer and antipsychotics), and a personalized approach to TMS stimulation are essential and likely needed. Table 2 for summary of rTMS and tDCS clinical trials in BD.
Cognitive alterations of neural network in BD
Mounting evidence suggests BD is associated with common cognitive deficits, including executive function, memory, social cognition, and response timing [51,52,53]. A large body of neuroimaging research has focused on topological network analyses of brain circuit, suggesting that modular and hierarchical structural networks are particularly suited for the functional integration of local neuronal operations that underlie cognitive function [54]. Synchronous activity and antisynchronous activity between neural elements at rest reflects the physiological process underlying complex cognitive ability. The tightly interconnected brain networks have emerged as the fundamental, dynamically organized elements of human brain function, consisting of patterns of synchronized activity across different brain regions [55,56,57]. These intrinsic brain networks are composed of hubs involved in important cognitive functions such as behavioral regulation and attention control and has transdiagnostic features that can serve as markers of affective and psychotic pathologies [58]. How cognitive processing emerges from interacting elements within the entire brain can be more meaningfully probed using a network theory framework. Neuroimaging research has revealed that the intrinsic brain activity, which is organized in distinct large-scale networks has been implicated in a variety of cognitive functions that are commonly disrupted in BD [59].
Although the neurophysiological basis of cognition and emotion are typically studied independently, recent work in this field increasingly highlight evidence showing substantial overlap in brain regions and networks involved in both emotion and cognitive processes. Cognition‐emotion integration requires the dynamic coordination and flexible switching between four functional networks that are linked to key adaptive processed, including emotion regulation, goal-directed attentional control, decision making, and self-monitoring [60].
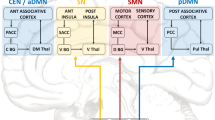
The DMN, also known as the “task-negative network”, was initially identified as brain regions showing consistently synchronized deactivation during tasks and prominent activation during rest [61] (Fig. 3). The CEN, also known as the “cognitive control network, includes the dlPFC, dorsal ACC, posterior parietal cortex, inferior temporal gyrus, and is involved in attention-demanding cognitive tasks and shows increased activity in frontal and parietal regions associated with top-down modulation of attention and working memory tasks [56, 62] (Fig. 3). The DMN and the CEN are often seen as opponent networks as the DMN is most active during rest, while CEN is most active during cognitive tasks. The SN typically consists of the fronto-insular cortex, the dorsal ACC, the amygdala, and the temporal pole, and is involved in interceptive awareness, task-set maintenance, and detection of salient stimuli from the environment [14] (Fig. 4). The SN plays a central role in switching between the DMN and CEN, and abnormalities of the SN could lead to weak salience mapping and give rise to dysfunctions of the CEN and DMN (Fig. 5). It has been reported that DMN and CEN act conversely, with the SN-mediating activity between the two networks, and both CEN and SN negatively regulate DMN function. The SMN displays coherent low-frequency (<0.1 Hz) activity fluctuations, which the sensory and motor/premotor cortical areas are reciprocally connected with the thalamus structurally and functionally [63] (Fig. 6).

The central executive network (CEN), also known as the “cognitive control network, includes the dorsolateral prefrontal cortex (dlPFC), dorsal ACC, posterior parietal cortex, inferior temporal gyrus, and precentral gyrus, and is involved in attention-demanding cognitive tasks and shows increased activity in frontal and parietal regions associated with top-down modulation of attention and working memory tasks. mPFC medial prefrontal cortex, PCC posterior cingulate cortex, PCU precuneus cortex, LTC laterl temporal cortex, Hip hippocampus, dlPFC dorsolateral prefrontal cortex, dACC dorsal ACC, dmPFC dorsomedial prefrontal cortex, PPC posterior parietal cortex, ITG inferior temporal gyrus.

The salience network (SN) consisting of the insular cortex, the dorsal ACC, the amygdala, and the temporal pole, is involved in interceptive awareness, task-set maintenance, and detection of salient stimuli from the environment. dACC dorsal ACC, IC insular cortex, Amy amygdala, TP temporal, VTA/SN ventral tegmental area/ substantia nigra. SMA supplementary motor area, PMC primary motor cortex, SC somatosensory cortex, IC insular cortex, AC auditory cortex, VC visual cortex.

The DMN/SMN balance was tilted toward the DMN in depression and toward the SMN in mania with bipolar disorder.

The sensorimotor network (SMN) mainly comprises the motor cortex (precentral gyrus, premotor and supplementary motor cortex) and middle cingulate cortex, somatosensory cortex (postcentral gyrus and parietal cortex), auditory cortex (superior temporal gyrus), as well as visual cortex (occipital cortex and associated dorsal parietal and inferior temporal areas) and is involved in exteroceptive awareness and motor activity.
Recently, evidence from fMRI studies point to aberrant activity of networks in BD as common neuronal correlates of cognitive impairment [64,65,66,67]. It has been shown that cognitive impairments arise from disruption of neuroplasticity and associated functional as well as structural changes in cognition-relevant neurocircuitries, with identified abnormalities in above four networks [8, 9, 68, 69]. The major finding indicates that BD is associated with alterations in both frontal and posterior DMN structures, mainly in the prefrontal, posterior cingulate and inferior parietal cortex [66]. Moreover, the hyperactivity of DMN has been clearly shown to ruminations and excessive internal (self-focused) thought, potentially playing a core role in depressive states. Martino et al. conducted a series of studies on BD investigating the different phases separately. It was found that the network balance in intrinsic brain activity was tilted toward the DMN in the depression phase but was shifted toward the SMN during mania [9]. The functional connectivity analyses revealed significantly stronger positive correlations between dlPFC and DMN (subgenual cingulate cortex) regions in asymptomatic patients with BD, suggesting BD patients may have greater difficulty switching from internally focused processing to task‐related processing in the presence of competing cognitive‐affective demands, compared to controls [70, 71]. Thus, the cognitive impairments in BD are likely caused by a failure to recruit key regions in the CEN to suppress task irrelevant DMN activity during cognitive performance [72].
In addition to decreased frontoparietal CEN recruitment, cognitive dysregulation in BD is also associated with disrupted SN recruitment [73]. A recent study has shown that bipolar depression is distinguished from both unipolar depression and healthy controls by significantly altered bilateral dorsal anterior insula functional connectivity to the left inferior parietal lobule (IPL), a key node of the frontoparietal CEN [74]. Another major finding is the cardinal role of the SN in BD disorder, characterized by a decrease in FC in several frontal and parietal regions and an increase in FC with the postcentral gyrus, precentral gyrus, temporal, and occipital regions. This conclusion is consistent with the meta-analysis suggesting the frontoparietal network may be the core common deficit in psychiatric disorders [75]. It has also been shown that cognitive impairment occurs across the psychosis spectrum and is associated with reduced cingulo-opercular network (CON), and subcortical network efficiency observed across the psychosis spectrum [76]. For example, a recent meta-analysis focused on the structural and functional brain abnormalities in unaffected relatives of individuals with BD. The results revealed hyperactivations in the fronto-striatal regions as well as hypoactivation in parietal regions during cognitive tasks, activation in the amygdala during emotional processing, and in the OFC during reward-related tasks [77]. In unaffected relatives, the right inferior frontal gyrus had greater volume and higher activation during cognitive tasks [77]. Regarding the preclinical studies investigating cognitive function in BD, we concluded the patients’ cognitive impairments may also be exacerbated by a failure to suppress neural activity in the DMN, most consistently the medial PFC and be associated with the aberrant encoding and working memory-related activity in the fronto-parietal, temporal regions during acute mood episodes and remission [78, 79]. A highly consistent finding was that aberrant functional connectivity within subcortical and PFC structures was linked with impaired? Working memory performance in BD [80, 81].
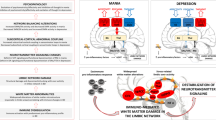
In BD, graph analysis studies indicate distinctive abnormalities of node-level and inter-hemispheric connectivity patterns, altered DMN dysconnectivity and decreased white matter organization of anterior limbic system tracts supported by reductions in efficiency and clustering connecting cingulate and subcortical limbic structures [82]. Magioncalda et al. proposed that BD was primarily characterized by immune-inflammatory factors causing white matter damage mainly encompassing the limbic network. Consequent phasic reconfigurations of intrinsic brain activity might superimpose onto structural damage in gray matter. The extent of gray matter damage could be related to cognitive deterioration that persist even during euthymia [83]. Specifically, the decreased organization of posterior white matter has been linked to impaired cognitive functions rather than emotional symptoms of BD [84]. White matter abnormality is a consistent finding in BD, as shown by several structural imaging studies since the 1990s and diffusion magnetic resonance imaging studies since the 2000s [85]. The longitudinal investigations measuring dynamics of functional connectivity across different mood states in the same patients, have shown that interoceptive sensorimotor activation pattern is more frequent during hypomania compared to other mood states, and is predictive for more severe symptoms of irritability and motor agitation. In contrast, a default-mode activation pattern is more frequent during depression compared to other mood states and compared to controls [86]. The white matter changes seem to represent a core and stable alteration in BD, while alterations in brain network balances seem to vary across the manic and depressive phases (Table 3).
Treatments targeting cognition regulation network dysfunctions in BD
In the past several years, studies have explored the application of rTMS in BD utilizing resting state electroencephalography (EEG) data. Changes in resting state networks (RSNs) between pre and post rTMS were explored utilizing EEG functional network analysis, evaluated by exact low-resolution electromagnetic tomography. Bilateral stimulation of dlPFC produced changes in the activity of the SMN and resulted in significant changes in the executive functions, verbal memory, and depression symptoms [87]. One study exploring deep (H1-coil) rTMS interventions focused on the dlPFC for cognitive improvement in BD shows that the deep rTMS is a safe antidepressant intervention in bipolar patients, who usually present marked cognitive impairment. Another randomized, placebo-controlled, double-blind trial also revealed that no cognitive impairment was found for BD patients treated with transcranial direct current stimulation over the dlPFC [88]. The optimal brain area to be stimulated to promote cognitive enhancement is the critical issue [89].
The studies on the neural correlates of TMS-induced cognitive function have shown that fMRI-based neurofeedback targeted at dlPFC and superior frontal gyrus and suppression of activity in DMN regions like the hippocampus in which activity was correlated with working memory performance resulted in cognitive improvement [90, 91]. Miskowiak et al found that 8 weeks of treatment with erythropoietin (EPO) increased working memory capacity in UD and BD, which was accompanied by enhanced task-related activity in the right superior frontal gyrus and deactivation of the left hippocampus. Furthermore, Meusel et al. showed that cognitive remediation therapy increased task-related activity in lateral PFC, medial frontal gyrus, superior temporal, and lateral parietal regions. Similarly, Haldane et al. found that lamotrigine treatment in remitted BD patients increased working memory-related activation over time in bilateral superior frontal gyri, cingulate gyri, and left medial frontal gyrus, even in the absence of changes in performance [92]. A preliminary study in youth with BD reported that the patients with bipolar depression treated with lamotrigine had decreased amygdala activation while viewing negative stimuli as depressive symptoms improved [93]. The same research group reported that potentially less efficient baseline activation of left dlPFC and increased baseline activation of left vlPFC during an affective task predicted greater treatment response of quetiapine in youth BD patients [94]. This study provides a neural signature for bipolar patient and indicates that individuals with BD can show both hypoactivity in the dlPFC and prolonged activation of mPFC simultaneously, suggesting the dorsal and ventral prefrontal subregions as promising candidates for biological markers of treatment response in bipolar depression. A better understanding of how these brain regions respond to different treatments may help guide more personalized treatments for patients with BD.
To date, nonpharmacological approach has been implemented to remediate cognitive impairments among people with BD using computerized cognitive training [95]. The cognitive remediation (CR) could tackle trait-like neurocognitive deficits in various cognitive domains such as attention, executive functions, verbal memory and learning, and social cognition. One study investigating the neural correlates induced by CR using fMRI showed increased activation in the left hippocampus that correlated with improvements in a recollection memory task, as well as increased activations in lateral and medial prefrontal and lateral parietal regions that correlated with memory improvements in a 2-back memory task [91].
Psychomotor alterations of neural network in BD
Although historically neglected, one of the most important findings in research of core behavioral features of BD is hyperactivity, reckless actions, impulsivity, and agitation during the manic phase, as well as physical and mental sluggishness, akinesia, volitional inhibition, and decreased activity levels during the depressive phase of illness. Manic and depressive episodes of BD show opposite psychomotor symptoms and the exact neurophysiological mechanism of psychomotor excitation and inhibition in mania and depression remain unclear. There are only a few MRI studies on the neural correlates of psychomotor phenomena. These psychomotor symptoms may depend on distinct patterns of alterations in the SMN, DMN, and CEN [9, 96, 97]. The SMN and related motor function are modulated by non-motor networks, such as DMN and CEN. The SMN mainly comprises the motor cortex (precentral gyrus, premotor and supplementary motor cortex) and middle cingulate cortex, somatosensory cortex (postcentral gyrus and parietal cortex), auditory cortex (superior temporal gyrus), as well as visual cortex (occipital cortex and associated dorsal parietal and inferior temporal areas), and is involved in exteroceptive awareness and motor activity [98]. The SMN is functionally connected with distinct regions of the thalamus, coherently with the structural connections, constituting segregated thalamo-cortical loops. The thalamo-cortical loops modulate the intrinsic activity and are directly connected with the external or internal environment, such that the sensory cortices receive inputs from the environment through the sensorial thalamic nuclei, while the motor and other specific cortices send outputs to the effector systems [99]. The thalamo-cortical loops are also connected with the basal ganglia and constitute different in-parallel cortico-striatopallido-thalamo-cortical loops. A recent study has reported that the BD patients show reduction in the cohesiveness of the SMN that likely disrupts the processing of sensorimotor information within the brain [100]. The alterations of interhemispheric dysfunction in SMN via corpus callosum using diffusion tensor imaging were reported in BD patients, which could be one of the pathophysiological bases of BD [101]. Other work has indicated that distinct changes within SMN and DMN during different phases of BD, i.e. predominant disruption of local connectivity within the SMN during depression, while disruption of both local and distant connectivity in the DMN during mania, consistent with the evidence showing an opposite pattern of alterations in mania and depression [102]. Recently, Martino and Magioncalda proposed a new theory that different combinations in neurotransmitter signaling favor network balancing into distinct functional brain states, which manifest in different combinations of excitation or inhibition in psychomotricity, affectivity, and thought, resulting in the manic, depressive, and mixed states of BD [13]. In this model, intrinsic brain activity is organized in distinct units in accordance with connectivity patterns and related setting of input/output processing, underlying different behavioral/phenomenological dimensions. An external unit (mainly involving the SMN) responds to the external environment and sets the exteroceptive input/somatomotor output processing, underlying the psychomotor dimension. An internal unit (mainly involving the SN) responds to the internal/body environment and sets the interoceptive input/visceromotor output processing, underlying the affective dimension. An index of intrinsic neuronal activity that favors the motor response to incoming stimuli, the more of neuronal variability in the SMN, the more excitation of psychomotor activity following cyclothymic temperament. Neuronal variability in SMN is higher in cyclothymic temperament and lower in depressive temperament [103]. The disbalance between SMN and DMN has been detected in BD, which a predominance of SMN occurs in mania along with greater global signal representation in SMN areas and reduced connectivity within the DMN [9]. Conversely, a predominance of DMN occurs in depression, as suggested by tilting in SMN/DMN balance toward the DMN at the expense of SMN [9, 104]. Northoff et al has proposed that the psychomotor mechanisms and their biochemical modulation were an example of a dimensional approach as suggested in RDoC and spatiotemporal psychopathology [19]. They identified three neural mechanisms: (i) serotoninergic modulation of dopamine-based subcortical–cortical motor circuit, (ii) reciprocal balances of DMN and sensory networks with SMN, and (iii) local synchronization of SMN with the brain’s global activity [19].
Treatments targeting psychomotor symptoms in BD
The alterations in large-scale brain intrinsic activity, subcortical-cortical coupling, and neurotransmitters signaling, such as dopamine (DA) and serotonin (5-HT) have been independently detected in BD [9, 105, 106]. Based on the reported data, Conio et al. propose that the dopaminergic nigrostriatal and mesocorticolimbic pathway and serotonergic pathway modulate the balance of functional connectivity in SMN and DMN. The alterations in DA and 5-HT signaling lead to subcortical–cortical functional reorganization, which results in brain intrinsic activity (such as DMN and SMN) disbalancing, finally manifesting in manic or depressive states of BD [104]. Previous studies also demonstrate that altered DA and 5-HT transmission, which modulates activity of SMN and the DMN and balance between these networks, results in excitation or inhibition of affectivity, psychomotricity, and thought [5, 13]. The use of some DA antagonists and partial agonists to treat acute mania, bipolar depression and maintenance treatment has been proved by the Food and Drug Administration (FDA). It is likely that the altered DA neurotransmission caused by the ligands, which is associated with FC and activity in the SMN, SN, and DMN, improves psychomotor activity as well as affective-related functions, and contributes to the clinical efficacy of these drugs.
The psychomotor abnormalities are characterized by specific symptom pattern and constellation of motor, emotional, and cognitive symptoms [107, 108]. The essential intermediate unit between the prefrontal areas and limbic affective areas during emotion regulation is the supplementary motor area (SMA), which has been involved in stimulus reconceptualization and cognitive demands [26]. The study investigating anatomy and function of the somatosensory cortex in euthymic bipolar women has shown increased resting state functional connectivity between the right somatosensory cortex and the brain regions involved with affective regulation (insular cortex, inferior frontal gyrus and OFC) [109]. Chronic motor cortex stimulation has been reported as a treatment method for Parkinson’s disease and neuropathic pain [110, 111]. A recent case series highlights the efficacy of add-on high-definition transcranial direct current stimulation (HD-tDCS) over sham stimulation in alleviating obsessive compulsory disorder with comorbid BD when anodal stimulation of pre-SMA was provided with right orbitofrontal placement of cathode [112]. The data from a few studies suggest that tDCS stimulating the prefronto-cerebellar improve the impaired neurocognition, neurological soft signs, and sleep in BD during euthymia [113]. The improvement in sensory integration, motor sequencing and coordination, executive and visuospatial function after the tDCS session supports the hypothesis of prefronto-thalamic-cerebellar circuitry involvement in neurocognition with euthymic BD [114]. These stimulation methods (rTMS and tDCS) may be applied in SMN systems that the latter modulate subcortical–cortical motor circuits and thereby alleviate psychomotor symptoms.
Conclusion
The neurobiological models that draw links between internal cognitive deficits, emotional processing, psychomotor activity dysfunction and neurocircuitry are beneficial because this approach meaningfully increases our understanding of BD with respect to clinical management. We propose a heuristic model of how to integrate information generated by recent neuroimaging studies into guiding principles toward treatments of BD. We hypothesized that (1) the core pathological alteration in BD is damage of fronto-limbic network that mainly results in emotional dysfunction; (2) that changes in intrinsic brain network, such as SMN, SN, DMN, CEN are associated with alterations of cognitive function; and (3) beyond the dopamine-driven basal ganglia-thalamo-cortical motor circuit modulated by other neurotransmitter systems such as 5-HT (subcortical–cortical modulation), the SMN and related motor function modulated by other non-motor networks such as the DMN are involved in psychomotor function. Although cognitive, emotional, and psychomotor domains are typically studied independently, basic research and emergent findings in BD suggest that there are important ties between cognitive deficits, the emotional and psychomotor disturbances observed in BD. Understanding these relationships is critical for elucidating relevant aspects related to functionality and vulnerability within BD and is essential for the development of novel treatment interventions. The following strategies based on neurocircuitry include: (1) enhancing PFC function to downregulate overactivated fronto-limbic activity via the engagement of top-down dorsal prefrontal regions, to alleviate impairments in emotion regulation; (2) modulating neurotransmitter transmission to balance the activities of excitatory and inhibitory neural networks involved in psychomotricity, affectivity and cognition; and (3) non-invasive neurostimulation in different cortical systems that modulate subcortical–cortical motor circuits to alleviate psychomotor symptoms. If the therapies of BD targeting specific circuit and corresponding clinical profiles, the treatment efficacy will be improved and guide clinical practice.
References
Solé B, Jiménez E, Torrent C, Reinares M, Bonnin CDM, Torres I, et al. Cognitive impairment in bipolar disorder: treatment and prevention strategies. Int J Neuropsychopharmacol. 2017;20:670–80.
Faurholt-Jepsen M, Brage S, Vinberg M, Christensen EM, Knorr U, Jensen HM, et al. Differences in psychomotor activity in patients suffering from unipolar and bipolar affective disorder in the remitted or mild/moderate depressive state. J Affect Disord. 2012;141:457–63.
Brotman MA, Tseng WL, Olsavsky AK, Fromm SJ, Muhrer EJ, Rutenberg JG, et al. Fronto-limbic-striatal dysfunction in pediatric and adult patients with bipolar disorder: impact of face emotion and attentional demands. Psychol Med. 2014;44:1639–51.
Caetano SC, Olvera RL, Glahn D, Fonseca M, Pliszka S, Soares JC. Fronto-limbic brain abnormalities in juvenile onset bipolar disorder. Biol Psychiatry. 2005;58:525–31.
Martino M, Magioncalda P, Conio B, Capobianco L, Russo D, Adavastro G, et al. Abnormal functional relationship of sensorimotor network with neurotransmitter-related nuclei via subcortical-cortical loops in manic and depressive phases of bipolar disorder. Schizophr Bull. 2020;46:163–74.
Phillips ML, Swartz HA. A critical appraisal of neuroimaging studies of bipolar disorder: toward a new conceptualization of underlying neural circuitry and a road map for future research. Am J Psychiatry. 2014;171:829–43.
Henry C, Van den Bulke D, Bellivier F, Roy I, Swendsen J, M’Baïlara K, et al. Affective lability and affect intensity as core dimensions of bipolar disorders during euthymic period. Psychiatry Res. 2008;159:1–6.
Ongür D, Lundy M, Greenhouse I, Shinn AK, Menon V, Cohen BM, et al. Default mode network abnormalities in bipolar disorder and schizophrenia. Psychiatry Res. 2010;183:59–68.
Martino M, Magioncalda P, Huang Z, Conio B, Piaggio N, Duncan NW, et al. Contrasting variability patterns in the default mode and sensorimotor networks balance in bipolar depression and mania. Proc Natl Acad Sci USA. 2016;113:4824–9.
Anticevic A, Cole MW, Repovs G, Murray JD, Brumbaugh MS, Winkler AM, et al. Characterizing thalamo-cortical disturbances in schizophrenia and bipolar illness. Cerebral cortex (New York, NY: 1991). 2014;24:3116–30.
Chen CH, Suckling J, Lennox BR, Ooi C, Bullmore ET. A quantitative meta-analysis of fMRI studies in bipolar disorder. Bipolar Disord. 2011;13:1–15.
Vai B, Bollettini I, Benedetti F. Corticolimbic connectivity as a possible biomarker for bipolar disorder. Expert Rev Neurother. 2014;14:631–50.
Martino M, Magioncalda P. Tracing the psychopathology of bipolar disorder to the functional architecture of intrinsic brain activity and its neurotransmitter modulation: a three-dimensional model. Molecular Psychiatry. 2021. https://doi.org/10.1038/s41380-020-00982-2.
Seeley WW, Menon V, Schatzberg AF, Keller J, Glover GH, Kenna H, et al. Dissociable intrinsic connectivity networks for salience processing and executive control. J Neuroscience. 2007;27:2349–56.
Dosenbach NU, Fair DA, Miezin FM, Cohen AL, Wenger KK, Dosenbach RA, et al. Distinct brain networks for adaptive and stable task control in humans. Proc Natl Acad Sci USA. 2007;104:11073–8.
Dosenbach NU, Fair DA, Cohen AL, Schlaggar BL, Petersen SE. A dual-networks architecture of top-down control. Trends Cogn Sci. 2008;12:99–105.
Bassett DS, Yang M, Wymbs NF, Grafton ST. Learning-induced autonomy of sensorimotor systems. Nat Neurosci. 2015;18:744–51.
Syan SK, Smith M, Frey BN, Remtulla R, Kapczinski F, Hall GBC, et al. Resting-state functional connectivity in individuals with bipolar disorder during clinical remission: a systematic review. J Psychiatry Neurosci. 2018;43:298–316.
Northoff G, Hirjak D, Wolf RC, Magioncalda P, Martino M. All roads lead to the motor cortex: psychomotor mechanisms and their biochemical modulation in psychiatric disorders. Mol Psychiatry. 2021;26:92–102.
M’Bailara K, Atzeni T, Colom F, Swendsen J, Gard S, Desage A, et al. Emotional hyperreactivity as a core dimension of manic and mixed states. Psychiatry Res. 2012;197:227–30.
Vai B, Bertocchi C, Benedetti F. Cortico-limbic connectivity as a possible biomarker for bipolar disorder: where are we now? Expert Rev Neurother. 2019;19:159–72.
Carlén M. What constitutes the prefrontal cortex? Science 2017;358:478–82.
Fuster JM. Frontal lobe and cognitive development. J Neurocytol. 2002;31:373–85.
Phillips ML, Ladouceur CD, Drevets WC. A neural model of voluntary and automatic emotion regulation: implications for understanding the pathophy-siology and neurodevelopment of bipolar disorder. Mol Psychiatry. 2008;13:833–57.
Ongür D, Price JL. The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys and humans. Cerebral Cortex (New York, NY: 1991). 2000;10:206–19.
Kohn N, Eickhoff SB, Scheller M, Laird AR, Fox PT, Habel U. Neural network of cognitive emotion regulation—an ALE meta-analysis and MACM analysis. Neuroimage 2014;87:345–55.
Wessa M, Linke J. Emotional processing in bipolar disorder: behavioural and neuroimaging findings. Int Rev Psychiatry. 2009;21:357–67.
Lawrence NS, Williams AM, Surguladze S, Giampietro V, Brammer MJ, Andrew C, et al. Subcortical and ventral prefrontal cortical neural responses to facial expressions distinguish patients with bipolar disorder and major depression. Biol Psychiatry. 2004;55:578–87.
Strakowski SM, Adler CM, Almeida J, Altshuler LL, Blumberg HP, Chang KD, et al. The functional neuroanatomy of bipolar disorder: a consensus model. Bipolar Disord. 2012;14:313–25.
Hassel S, Almeida JR, Kerr N, Nau S, Ladouceur CD, Fissell K, et al. Elevated striatal and decreased dorsolateral prefrontal cortical activity in response to emotional stimuli in euthymic bipolar disorder: no associations with psychotropic medication load. Bipolar Disord. 2008;10:916–27.
Cerullo MA, Fleck DE, Eliassen JC, Smith MS, DelBello MP, Adler CM, et al. A longitudinal functional connectivity analysis of the amygdala in bipolar I disorder across mood states. Bipolar Disord. 2012;14:175–84.
Townsend JD, Torrisi SJ, Lieberman MD, Sugar CA, Bookheimer SY, Altshuler LL. Frontal-amygdala connectivity alterations during emotion downregulation in bipolar I disorder. Biol Psychiatry. 2013;73:127–35.
Radaelli D, Sferrazza Papa G, Vai B, Poletti S, Smeraldi E, Colombo C, et al. Fronto-limbic disconnection in bipolar disorder. Eur Psychiatry. 2015;30:82–8.
Altshuler L, Bookheimer S, Proenza MA, Townsend J, Sabb F, Firestine A, et al. Increased amygdala activation during mania: a functional magnetic resonance imaging study. Am J Psychiatry. 2005;162:1211–3.
Bigot M, Alonso M, Houenou J, Sarrazin S, Dargel AA, Lledo PM, et al. An emotional-response model of bipolar disorders integrating recent findings on amygdala circuits. Neurosci Biobehav Rev. 2020;118:358–66.
Mitchell PB, Hadzi-Pavlovic D. Lithium treatment for bipolar disorder. Bull World Health Organ. 2000;78:515–7.
Berk M, Dandash O, Daglas R, Cotton SM, Allott K, Fornito A, et al. Neuroprotection after a first episode of mania: a randomized controlled maintenance trial comparing the effects of lithium and quetiapine on grey and white matter volume. Transl Psychiatry. 2017;7:e1011.
Yucel K, McKinnon MC, Taylor VH, Macdonald K, Alda M, Young LT, et al. Bilateral hippocampal volume increases after long-term lithium treatment in patients with bipolar disorder: a longitudinal MRI study. Psychopharmacology. 2007;195:357–67.
Chiang KJ, Tsai JC, Liu D, Lin CH, Chiu HL, Chou KR. Efficacy of cognitive-behavioral therapy in patients with bipolar disorder: a meta-analysis of randomized controlled trials. PLoS ONE. 2017;12:e0176849.
Szentagotai A, David D. The efficacy of cognitive-behavioral therapy in bipolar disorder: a quantitative meta-analysis. J Clin Psychiatry. 2010;71:66–72.
Lam DH, Burbeck R, Wright K, Pilling S. Psychological therapies in bipolar disorder: the effect of illness history on relapse prevention - a systematic review. Bipolar Disord. 2009;11:474–82.
Zhang L, Opmeer EM, van der Meer L, Aleman A, Curcic-Blake B, Ruhe HG. Altered frontal-amygdala effective connectivity during effortful emotion regulation in bipolar disorder. Bipolar Disord. 2018;20:349–58.
Connolly KR, Helmer A, Cristancho MA, Cristancho P, O’Reardon JP. Effectiveness of transcranial magnetic stimulation in clinical practice post-FDA approval in the United States: results observed with the first 100 consecutive cases of depression at an academic medical center. J Clin Psychiatry. 2012;73:e567–73.
Gold AK, Ornelas AC, Cirillo P, Caldieraro MA, Nardi AE, Nierenberg AA, et al. Clinical applications of transcranial magnetic stimulation in bipolar disorder. Brain Behav. 2019;9:e01419.
Grisaru N, Chudakov B, Yaroslavsky Y, Belmaker RH. Transcranial magnetic stimulation in mania: a controlled study. Am J Psychiatry. 1998;155:1608–10.
McGirr A, Karmani S, Arsappa R, Berlim MT, Thirthalli J, Muralidharan K, et al. Clinical efficacy and safety of repetitive transcranial magnetic stimulation in acute bipolar depression. World Psychiatry. 2016;15:85–6.
Harel EV, Zangen A, Roth Y, Reti I, Braw Y, Levkovitz Y. H-coil repetitive transcranial magnetic stimulation for the treatment of bipolar depression: an add-on, safety and feasibility study. World J Biol Psychiatry. 2011;12:119–26.
Tavares DF, Myczkowski ML, Alberto RL, Valiengo L, Rios RM, Gordon P, et al. Treatment of bipolar depression with deep TMS: results from a Double-Blind, Randomized, Parallel Group, Sham-Controlled Clinical Trial. Neuropsychopharmacology 2017;42:2593–601.
Hett D, Marwaha S. Repetitive transcranial magnetic stimulation in the treatment of bipolar disorder. Ther Adv Psychopharmacol. 2020;10. https://doi.org/10.1177/2045125320973790.
Fitzgerald PB, Hoy KE, Elliot D, McQueen S, Wambeek LE, Daskalakis ZJ. A negative double-blind controlled trial of sequential bilateral rTMS in the treatment of bipolar depression. J Affect Disord. 2016;198:158–62.
Bora E, Bartholomeusz C, Pantelis C. Meta-analysis of Theory of Mind (ToM) impairment in bipolar disorder. Psychol Med. 2016;46:253–64.
Mann-Wrobel MC, Carreno JT, Dickinson D. Meta-analysis of neuropsychological functioning in euthymic bipolar disorder: an update and investigation of moderator variables. Bipolar Disord. 2011;13:334–42.
Bourne C, Aydemir O, Balanza-Martinez V, Bora E, Brissos S, Cavanagh JT, et al. Neuropsychological testing of cognitive impairment in euthymic bipolar disorder: an individual patient data meta-analysis. Acta Psychiatr Scand. 2013;128:149–62.
Park HJ, Friston K. Structural and functional brain networks: from connections to cognition. Science 2013;342:1238411.
Beckmann CF, DeLuca M, Devlin JT, Smith SM. Investigations into resting-state connectivity using independent component analysis. Philos Trans R Soc Lond B Biol Sci. 2005;360:1001–13.
Smith SM, Fox PT, Miller KL, Glahn DC, Fox PM, Mackay CE, et al. Correspondence of the brain’s functional architecture during activation and rest. Proc Natl Acad Sci USA. 2009;106:13040–5.
Biswal B, Yetkin FZ, Haughton VM, Hyde JS. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn Reson Med. 1995;34:537–41.
Bellani M, Bontempi P, Zovetti N, Gloria Rossetti M, Perlini C, Dusi N, et al. Resting state networks activity in euthymic bipolar disorder. Bipolar Disord. 2020;22:593–601.
McPhilemy G, Nabulsi L, Kilmartin L, Whittaker JR, Martyn FM, Hallahan B, et al. Resting-state network patterns underlying cognitive function in bipolar disorder: a graph theoretical analysis. Brain Connect. 2020;10:355–67.
Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol. 2011;106:1125–65.
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci USA. 2001;98:676–82.
Menon V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn Sci. 2011;15:483–506.
Yuan R, Di X, Taylor PA, Gohel S, Tsai YH, Biswal BB. Functional topography of the thalamocortical system in human. Brain Struct Funct. 2016;221:1971–84.
Wang J, Wang Y, Wu X, Huang H, Jia Y, Zhong S, et al. Shared and specific functional connectivity alterations in unmedicated bipolar and major depressive disorders based on the triple-network model. Brain Imaging Behav. 2020;14:186–99.
Wang J, Wang Y, Huang H, Jia Y, Zheng S, Zhong S, et al. Abnormal dynamic functional network connectivity in unmedicated bipolar and major depressive disorders based on the triple-network model. Psychol Med. 2020;50:465–74.
Zovetti N, Rossetti MG, Perlini C, Maggioni E, Bontempi P, Bellani M, et al. Default mode network activity in bipolar disorder. Epidemiol Psychiatr Sci. 2020;29:e166.
Liu M, Wang Y, Zhang A, Yang C, Liu P, Wang J, et al. Altered dynamic functional connectivity across mood states in bipolar disorder. Brain Res. 2021;1750:147143.
Baker JT, Holmes AJ, Masters GA, Yeo BT, Krienen F, Buckner RL, et al. Disruption of cortical association networks in schizophrenia and psychotic bipolar disorder. JAMA Psychiatry. 2014;71:109–18.
Lopez-Larson MP, Shah LM, Weeks HR, King JB, Mallik AK, Yurgelun-Todd DA, et al. Abnormal functional connectivity between default and salience networks in pediatric bipolar disorder. Biol Psychiatry Cogn Neurosci Neuroimaging. 2017;2:85–93.
Rai S, Griffiths KR, Breukelaar IA, Barreiros AR, Chen W, Boyce P, et al. Default-mode and fronto-parietal network connectivity during rest distinguishes asymptomatic patients with bipolar disorder and major depressive disorder. Transl Psychiatry. 2021;11:547.
Favre P, Polosan M, Pichat C, Bougerol T, Baciu M. Cerebral correlates of abnormal emotion conflict processing in euthymic bipolar patients: a functional MRI Study. PLoS ONE. 2015;10:e0134961.
Zarp Petersen J, Varo C, Skovsen CF, Ott CV, Kjaerstad HL, Vieta E, et al. Neuronal underpinnings of cognitive impairment in bipolar disorder: a large data-driven functional magnetic resonance imaging study. Bipolar Disord. 2021. https://doi.org/10.1111/bdi.13100.
Goya-Maldonado R, Brodmann K, Keil M, Trost S, Dechent P, Gruber O. Differentiating unipolar and bipolar depression by alterations in large-scale brain networks. Hum Brain Mapp. 2016;37:808–18.
Ellard KK, Zimmerman JP, Kaur N, Van Dijk KRA, Roffman JL, Nierenberg AA, et al. Functional connectivity between anterior insula and key nodes of frontoparietal executive control and salience networks distinguish bipolar depression from unipolar depression and healthy control subjects. Biol Psychiatry Cogn Neurosci Neuroimaging. 2018;3:473–84.
Tu PC, Bai YM, Li CT, Chen MH, Lin WC, Chang WC, et al. Identification of common thalamocortical dysconnectivity in four major psychiatric disorders. Schizophr Bull. 2019;45:1143–51.
Sheffield JM, Kandala S, Tamminga CA, Pearlson GD, Keshavan MS, Sweeney JA, et al. Transdiagnostic associations between functional brain network integrity and cognition. JAMA Psychiatry. 2017;74:605–13.
Cattarinussi G, Di Giorgio A, Wolf RC, Balestrieri M, Sambataro F. Neural signatures of the risk for bipolar disorder: a meta-analysis of structural and functional neuroimaging studies. Bipolar Disord. 2019;21:215–27.
Alonso-Lana S, Valentí M, Romaguera A, Sarri C, Sarró S, Rodríguez-Martínez A, et al. Brain functional changes in first-degree relatives of patients with bipolar disorder: evidence for default mode network dysfunction. Psychol Med. 2016;46:2513–21.
Adler CM, Holland SK, Schmithorst V, Tuchfarber MJ, Strakowski SM. Changes in neuronal activation in patients with bipolar disorder during performance of a working memory task. Bipolar Disord. 2004;6:540–9.
McKenna BS, Sutherland AN, Legenkaya AP, Eyler LT. Abnormalities of brain response during encoding into verbal working memory among euthymic patients with bipolar disorder. Bipolar Disord. 2014;16:289–99.
Stegmayer K, Usher J, Trost S, Henseler I, Tost H, Rietschel M, et al. Disturbed cortico-amygdalar functional connectivity as pathophysiological correlate of working memory deficits in bipolar affective disorder. Eur Arch Psychiatry Clin Neurosci. 2015;265:303–11.
O’Donoghue S, Holleran L, Cannon DM, McDonald C. Anatomical dysconnectivity in bipolar disorder compared with schizophrenia: a selective review of structural network analyses using diffusion MRI. J Affect Disord. 2017;209:217–28.
Magioncalda P, Martino M. A unified model of the pathophysiology of bipolar disorder. Mol Psychiatry. 2021. https://doi.org/10.1038/s41380-021-01091-4.
Nortje G, Stein DJ, Radua J, Mataix-Cols D, Horn N. Systematic review and voxel-based meta-analysis of diffusion tensor imaging studies in bipolar disorder. J Affect Disord. 2013;150:192–200.
Mahon K, Burdick KE, Szeszko PR. A role for white matter abnormalities in the pathophysiology of bipolar disorder. Neurosci Biobehav Rev. 2010;34:533–54.
Rey G, Bolton TAW, Gaviria J, Piguet C, Preti MG, Favre S, et al. Dynamics of amygdala connectivity in bipolar disorders: a longitudinal study across mood states. Neuropsychopharmacology. 2021;46:1693–701.
Kazemi R, Rostami R, Khomami S, Baghdadi G, Rezaei M, Hata M, et al. Bilateral transcranial magnetic stimulation on DLPFC changes resting state networks and cognitive function in patients with bipolar depression. Front Hum Neurosci. 2018;12:356.
Tortella G, Sampaio-Junior B, Moreno ML, Moffa AH, da Silva AF, Lafer B, et al. Cognitive outcomes of the bipolar depression electrical treatment trial (BETTER): a randomized, double-blind, sham-controlled study. Eur Arch Psychiatry Clin Neurosci. 2021;271:93–100.
Myczkowski ML, Fernandes A, Moreno M, Valiengo L, Lafer B, Moreno RA, et al. Cognitive outcomes of TMS treatment in bipolar depression: safety data from a randomized controlled trial. J Affect Disord. 2018;235:20–6.
Miskowiak KW, Vinberg M, Glerup L, Paulson OB, Knudsen GM, Ehrenreich H, et al. Neural correlates of improved executive function following erythropoietin treatment in mood disorders. Psychol Med. 2016;46:1679–91.
Meusel LA, Hall GB, Fougere P, McKinnon MC, MacQueen GM. Neural correlates of cognitive remediation in patients with mood disorders. Psychiatry Res. 2013;214:142–52.
Haldane M, Jogia J, Cobb A, Kozuch E, Kumari V, Frangou S. Changes in brain activation during working memory and facial recognition tasks in patients with bipolar disorder with Lamotrigine monotherapy. Eur Neuropsychopharmacol. 2008;18:48–54.
Chang KD, Wagner C, Garrett A, Howe M, Reiss A. A preliminary functional magnetic resonance imaging study of prefrontal-amygdalar activation changes in adolescents with bipolar depression treated with lamotrigine. Bipolar Disord. 2008;10:426–31.
Chang K, DelBello M, Garrett A, Kelley R, Howe M, Adler C, et al. Neurofunctional correlates of response to quetiapine in adolescents with bipolar depression. J Child Adolesc Psychopharmacol. 2018;28:379–86.
Bellani M, Biagianti B, Zovetti N, Rossetti MG, Bressi C, Perlini C, et al. The effects of cognitive remediation on cognitive abilities and real-world functioning among people with bipolar disorder: a systematic review: Special Section on “Translational and Neuroscience Studies in Affective Disorders”. Section Editor, Maria Nobile MD, PhD. This section of JAD focuses on the relevance of translational and neuroscience studies in providing a better understanding of the neural basis of affective disorders. The main aim is to briefly summaries relevant research findings in clinical neuroscience with particular regards to specific innovative topics in mood and anxiety disorders. J Affect Disord. 2019;257:691–7.
Mamah D, Barch DM, Repovs G. Resting state functional connectivity of five neural networks in bipolar disorder and schizophrenia. J Affect Disord. 2013;150:601–9.
Meda SA, Ruaño G, Windemuth A, O’Neil K, Berwise C, Dunn SM, et al. Multivariate analysis reveals genetic associations of the resting default mode network in psychotic bipolar disorder and schizophrenia. Proc Natl Acad Sci USA. 2014;111:E2066–75.
Wang P, Kong R, Kong X, Liegeois R, Orban C, Deco G, et al. Inversion of a large-scale circuit model reveals a cortical hierarchy in the dynamic resting human brain. Sci Adv. 2019;5:eaat7854.
Amaral D. The functional organization of perception and movement. In: Kandel E, Schwartz J, Jessell T, Siegelbaum S, Hudspeth A, (eds). Principles of Neural Science - Fifth edition. USA: McGraw Hill 2013. p 356–369.
Doucet GE, Bassett DS, Yao N, Glahn DC, Frangou S. The role of intrinsic brain functional connectivity in vulnerability and resilience to bipolar disorder. Am J Psychiatry. 2017;174:1214–22.
Ishida T, Donishi T, Iwatani J, Yamada S, Takahashi S, Ukai S, et al. Interhemispheric disconnectivity in the sensorimotor network in bipolar disorder revealed by functional connectivity and diffusion tensor imaging analysis. Heliyon 2017;3:e00335.
Russo D, Martino M, Magioncalda P, Inglese M, Amore M, Northoff G. Opposing changes in the functional architecture of large-scale networks in bipolar mania and depression. Schizophr Bull. 2020;46:971–80.
Conio B, Magioncalda P, Martino M, Tumati S, Capobianco L, Escelsior A, et al. Opposing patterns of neuronal variability in the sensorimotor network mediate cyclothymic and depressive temperaments. Hum Brain Mapp. 2019;40:1344–52.
Conio B, Martino M, Magioncalda P, Escelsior A, Inglese M, Amore M, et al. Opposite effects of dopamine and serotonin on resting-state networks: review and implications for psychiatric disorders. Mol Psychiatry. 2020;25:82–93.
Cousins DA, Butts K, Young AH. The role of dopamine in bipolar disorder. Bipolar Disord. 2009;11:787–806.
Kato T. Current understanding of bipolar disorder: toward integration of biological basis and treatment strategies. Psychiatry Clin Neurosci. 2019;73:526–40.
Bernard JA, Mittal VA. Updating the research domain criteria: the utility of a motor dimension. Psychol Med. 2015;45:2685–9.
Sarkheil P, Odysseos P, Bee I, Zvyagintsev M, Neuner I, Mathiak K. Functional connectivity of supplementary motor area during finger-tapping in major depression. Compr Psychiatry. 2020;99:152166.
Minuzzi L, Syan SK, Smith M, Hall A, Hall GB, Frey BN. Structural and functional changes in the somatosensory cortex in euthymic females with bipolar disorder. Aust N Z J Psychiatry. 2018;52:1075–83.
Velasco F, Argüelles C, Carrillo-Ruiz JD, Castro G, Velasco AL, Jiménez F, et al. Efficacy of motor cortex stimulation in the treatment of neuropathic pain: a randomized double-blind trial. J Neurosurg. 2008;108:698–706.
Zwartjes DG, Heida T, Feirabend HK, Janssen ML, Visser-Vandewalle V, Martens HC, et al. Motor cortex stimulation for Parkinson’s disease: a modelling study. J Neural Eng. 2012;9:056005.
Parlikar R, Sreeraj VS, Chhabra H, Thimmashetty VH, Parameshwaran S, Selvaraj S, et al. Add-on HD-tDCS for obsessive-compulsive disorder with comorbid bipolar affective disorder: a case series. Asian J Psychiatr. 2019;43:87–90.
Donde C, Neufeld NH, Geoffroy PA. The impact of transcranial direct current stimulation (tDCS) on bipolar depression, mania, and euthymia: a systematic review of preliminary data. Psychiatr Q. 2018;89:855–67.
Minichino A, Bersani FS, Bernabei L, Spagnoli F, Vergnani L, Corrado A, et al. Prefronto-cerebellar transcranial direct current stimulation improves visuospatial memory, executive functions, and neurological soft signs in patients with euthymic bipolar disorder. Neuropsychiatr Dis Treat. 2015;11:2265–70.
Acknowledgements
The authors would like to acknowledge the contributions of individuals with BD and their families to BD neuroimaging research, without whose selflessness, no discoveries would be possible. We would further like to acknowledge the efforts of investigators across the globe and their willingness to share data and results which spurs research on BD neuroimaging.
Author information
Authors and Affiliations
Contributions
BB, as corresponding author, read the literatures and proposed the ideas, then recapitulated them. DC contributed in response to the reviewer’s question in revised manuscript. So, the author was added to the revised manuscript. YB edited figures 1–6.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bi, B., Che, D. & Bai, Y. Neural network of bipolar disorder: Toward integration of neuroimaging and neurocircuit-based treatment strategies. Transl Psychiatry 12, 143 (2022). https://doi.org/10.1038/s41398-022-01917-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-022-01917-x
This article is cited by
-
Research on the effect of different aerobic activity on physical fitness and executive function in primary school students
Scientific Reports (2024)
-
Network dysfunction of sadness facial expression processing and morphometry in euthymic bipolar disorder
European Archives of Psychiatry and Clinical Neuroscience (2024)
-
Does treatment with autophagy-enhancers and/or ROS-scavengers alleviate behavioral and neurochemical consequences of low-dose rotenone-induced mild mitochondrial dysfunction in mice?
Molecular Psychiatry (2023)
-
Prefrontal, parietal, and limbic condition-dependent differences in bipolar disorder: a large-scale meta-analysis of functional neuroimaging studies
Molecular Psychiatry (2023)
-
Using nonlinear analysis and neural network to classify bipolar I disorder electroencephalogram signals from normal electroencephalograms
Network Modeling Analysis in Health Informatics and Bioinformatics (2023)
