
Abstract
The symbiotic bacteria that live in the human gut and the metabolites they produce have long influenced local and systemic physiological and pathological processes of the host. The gut microbiota are increasingly being recognized for its impact on a range of human diseases, including cancer, it may play a key role in the occurrence, progression, treatment, and prognosis of many types of cancer. Understanding the functional role of the gut microbiota in cancer is crucial for the development of the era of personalized medicine. Here, we review recent advances in research and summarize the important associations and clear experimental evidence for the role of the gut microbiota in a variety of human cancers, focus on the application and possible challenges associated with the gut microbiota in antitumor therapy. In conclusion, our research demonstrated the multifaceted mechanisms of gut microbiota affecting human cancer and provides directions and ideas for future clinical research.
Similar content being viewed by others

Introduction
In recent years, a large amount of evidence has shown that parasitic microorganisms in the human body are key factors in health or pathological conditions. Diseases including inflammatory bowel disease, atherosclerosis, multiple sclerosis, diabetes, and Alzheimer’s disease, among others are associated with dysbacteriosis [1, 2]. With the increase in the incidence of malignant tumors, the interactions between microbiota and cancer are increasingly emerging.
The historical record linking cancer and microbes dates back to 1868, when William Busch reported spontaneous tumor regression in cancer patients infected with Streptococcus pyogenes. Among the human microorganisms, the gut microbiota are the most widely studied; approximately 3 × 103 types of microbial cells inhabit the human body, numbering up to 4 × 1013 organisms in total, of which approximately 97% are gut microbiota. Firmicutes, Bacteroidetes, Actinomycetes, Proteobacteria, and Verrucomicrobia are the main bacterial groups present in the gut microbiota [3,4,5,6].
In 2015, Guinney et al. classified colorectal cancer (CRC) based on genes differentially expressed in tumor cells, resulting in a powerful classification system for CRC, the consensus molecular subtypes (CMS). These include CMS1 (immunoinvasive type, 14%), CMS2 (classical type, 37%), CMS3 (metabolic dysregulation type, 13%), and CMS4 (stromal invasion type, 23%) [7]. Later, Purcell et al. found that different CMS subtypes were associated with different gut microbiota compositions. Using 16 S rRNA gene sequencing, they found enrichment of Fusobacteria (15.7%) and Bacteroidetes (48.5%), and the absence of Firmicutes (<3%) and Proteobacteria (<3%) in CMS1 patients (the expression of immunoinfiltration-related genes is significantly increased) [8]. At the same time, CMS2 patients showed enrichment of Selenomas and Prevotella spp. (genes significantly associated with cell cycle were significantly upregulated in CMS2 patients). [7, 8]. This suggests that the composition of gut microbiota affects the immune and genetic patterns of CRC and other cancers, and the mechanism may be complex.
In recent years, studies in metabolomics and genomics have emphasized the dual role of gut microbiota in cancer prevention, occurrence and anticancer therapy; that is, the gut microbiota can both suppress and promote tumors [9]. In this review, we discuss how the gut microbiota play a role in human cancer and its application in anticancer therapy.
Gut microbiota play a role in the development and progression of cancer
Experimental models have shown that as the largest microbiota in human body, gut microbiota play a key role in cancer by influencing immunity, genetic material, and cell pathways [10] (Supplementary information: Supplementary Table).
In recent years, 16 S rRNA gene sequencing and metagenomic analysis have revealed widespread differences in gut microbiota diversity between cancer patients and healthy individuals [11]. 16 S rRNA is an rRNA involved in the manufacture of small subunits of prokaryotic ribosomes; it provides low-cost and reliable identification of the overall microbiome composition [12, 13]. Unlike 16 S rRNA gene sequencing, metagenomic analysis does not target a specific microbial population, nor does it sequence a single microbial population. Instead, metagenomic analysis is performed as a sum of all microbial genomes. As Laudadio et al report, metagenomic sequencing analysis can help characterize the complexity of the microbiome in greater detail than 16 S rRNA gene sequencing [14]. The relationship between the gut microbiota and cancer continues to be elucidated owing to the use of increasingly advanced microbial detection techniques.
Below, we explore the role of gut microbiota in cancer through different mechanisms, including immunity, DNA damage, cellular signaling pathways, and inflammasomes (Table 1).
Immune microenvironment
Gut microbiota and antitumor immunity
In recent years, more and more studies have focused on the regulatory effect of microflora on host immunity. Researchers often associate tumor immunology with microbiota [15], especially non-pathogenic microbiota, which is often not the direct cause of some diseases, but affects local and distant pathological processes, among which gut microbiota has a complex and critical interaction mechanism with the immune system. The gut microbiota helps the immune system tolerate foreign antigens from food; furthermore, it helps the immune system recognize and eliminate pathogenic bacteria, thereby preventing the invasion of pathogens [16]. In general, gut microbes and products produced by microbes (such as short-chain fatty acids [SCFAs]) activate immune responses by interacting with immune cells expressing toll-like receptors (TLRs). For example, SCFAs will promote the differentiation of naïve T-cells to Th1 cells, thereby enhancing immunity, and dendritic cells (DCs) activated by SCFAs migrates from the GI tract to the mesenteric lymph nodes, inducing naïve T-cells to transform into effector T-cells, different types of effector T-cells migrate to the gastrointestinal tract again and stimulate local immune responses, while the remaining cells enter the systemic circulation, affecting system-wide immunity [17,18,19,20,21,22]. In short, the gut microbiota interact with immune cells or alters the immunogenicity of tumor cells, affecting the antitumor effect of the host immune system and even leading to differential responses to immunotherapy [23]. These complex and important regulatory processes are described in detail below (Fig. 1).

The gut microbiota and metabolites in the intestinal lumen are sensed by dendritic cells (DCs), which then induce the transformation of naive T-cells into various effector T-cells. In this process, butyrate inhibits DC activation of naive T-cells and secretion of IL-6, −8, and −12, and other factors, while promoting the transformation of naive T-cells into Treg. Bacteroides fragilis toxin (BFT) promoted the transformation of naive T-cells into Tfh cells and Th17. Staphylococcal enterotoxin B promotes the transformation of naive T-cells into Th9 cells. Inosine and short-chain fatty acids promote the transformation of naive T-cells into Th1 cells, and trimethylamine N-oxide promotes the secretion of IL-2 and −3, and other factors by Th1 cells. In terms of innate immunity, myeloid-derived suppressor cells reach the intestinal tract through the blood and secrete cancer-promoting factors, such as Arg-1, nitric oxide synthase (NOS), and reactive oxygen species. In this process, FAP2 plays a promoting role. Simultaneously, FAP2 binds and blocks the receptor TIGIT on NK T-cells, thus inhibiting the NK T-mediated tumor cell attack process. Secondary bile acid decreases the expression of CXCL16 on the surface of antigen-presenting cells (APCs) and prevent the aggregation of CXCR6+ NK T-cells. Intracellular RIG-I on APCs recognizes the abnormal DNA of the bacterial community and transmits the signal to mitochondrial anti-viral signaling proteins (MAVS) on the mitochondrial membrane, which in turn activates the NF-kB signal and releases IFN-α, a process in which MAVS plays a catalytic role.
Gut microbiota and innate immunity
Physiological protective barrier is one of the important components of innate immunity, including intestinal and symbiotic bacteria. It has been reported that the abundance of Lactobacillus and Bifidobacteria in breast milk plays an important role in the construction of neonatal innate immunity [24]. During innate immunity, Fusobacterium nucleatum can inhibit host natural killer (NK) cells and recruit myeloid-derived suppressor cells (MDSCs) at the site of infection, thereby indirectly promoting the occurrence of cancer. This process is mediated by the bacterial virulence factor FAP2, which binds to and blocks the NK receptor TIGIT, thereby inhibiting the NK-mediated immune system’s attack on tumor cells [25]. Similarly, in a mouse model of CRC, Fusobacterium inhibited T-cell responses by recruiting tumor-infiltrating immune cells and manipulating the innate immune system, producing an immune microenvironment conducive to colorectal tumor progression [26]. Secondary bile acids produced by gut microbiota decreased CXCL16 (the only ligand for CXCR6 (CXC Receptor 6)) expression on hepatocyte surface, demonstrated by higher levels of CXCL16 mRNA in the liver of germ-free mice (about twice), which prevent the aggregation and immune monitoring of CXCR6+ T-cells, thereby causing liver cancer. This immune escape reaction can be eliminated by antibiotic treatment such as vancomycin [27].
Gut microbiota and primary lymphoid organs
The gut microbiota also induce TNF-α expression via tumor-associated natural bone marrow cells, mediating TNF-dependent early tumor necrosis [28]. In addition, after bone marrow transplantation, a decrease in the number of gut microbiota count may exacerbate systemic infection and increase the radiation sensitivity, whereas higher gut microbiota diversity significantly improved the efficacy of allogeneic hematopoietic stem cell transplantation in leukemia patients. Specifically, there were 104 deaths among 354 patients in the high-diversity group and 136 deaths among 350 patients in the low-diversity group [29]. Subsequent studies have shown that certain compounds produced by gut microbial metabolism, such as propionic acid and tryptophan, enhance the function of bone marrow cells and neutrophils derived from bone marrow transplantation and prevent hematopoietic injury caused by bone marrow transplantation [30,31,32]. Translocation of gut bacteria in mice also exacerbates pre-leukemia bone marrow dysplasia, which can lead to precursor B-cell acute lymphoblastic leukemia (pB-ALL). Of a total of 23 mice with pB-ALL defects, 11 developed pB-ALL between 11 and 20 months of age, while none developed in germ-free mice (n = 12) [33]. The effects of the gut microbiota on lymphoid organs may be due to the activation of mitochondrial anti-viral signaling proteins (MAVS) by endogenous ligands (such as viruses, bacteriophages, or bacteria-derived RNA) of RIG-I (a receptor that recognizes abnormal mRNA in cells), which in turn induces protective signaling by IFN-I [34].
Gut microbiota and adaptive immunity
In adaptive immunity, the gut microbiota elicit host-specific T-cell responses in an antigen-presenting manner, in which the microbiota and its metabolites significantly affect the body’s antitumor immune effects [35, 36]. T-cell differentiation can be divided into three effector pathways: Th1, Th2, and Th17 responses. The STAT1 and STAT4 signaling pathways promote the Th1 response, the STAT6 signaling pathway promotes the Th2 response, and the STAT3 signaling pathway promotes the Th17 response. The Th1 response is characterized by the production of IFN-γ, which generally has anticancer effects, although it also plays roles in allergic and inflammatory reactions. However, the toxin secreted by Bacteroides fragilis (BFT) significantly increases colon tumor formation by rapidly, strongly, and selectively activating STAT3 and promoting the Th17 response, a process accompanied by the activation of serine/threonine mitogen-activated protein kinase (MAPK) and NF-kB signaling [37,38,39,40]. In addition, Trimethylamine N-oxide (TMAO), a metabolic derivative produced by gut microbiota that helps the body metabolize choline or trimethylamine foods, has been demonstrated to enhance the antitumor immunity to pancreatic ductal adenocarcinoma in mouse models; researchers have described the mechanism by which TMAO enhances the INF-I signaling pathway and enhances the antitumor effect in an INF-I-dependent manner [41]. Inosine is a purine metabolite that acts as an important modulator of the immune checkpoint blockade therapy response. Bifidobacterium dentium pseudotudes and Lactobacillus johnsonii in the intestine produce inosine in the systemic circulation and induce Th1 differentiation and effector function [42].
Gut microbiota and immune cells
DCs are among the most important antigen-presenting cells in the human body, antigens produced by gut microbiota or their metabolites can be used to activate dc to reverse immune tolerance induced by immature DCs [43]. DCs from the gut-associated lymphoid tissue area sense various gut microbiota antigens, including Bifidobacteria, Bacteroides fragilis, Myxobacterium, Bacillus rodentia, Bacteroides, and their metabolites, and catalyze immune reactions through IFN-I and IL-12 [44,45,46,47]. In a melanoma mouse model, Bifidobacteria activated DCs through the TLR4-mediated signaling pathway, and then DCs amplified the CD8+ T-cell response in the tumor microenvironment. Through ELISPOT and flow cytometry, it was found that the mechanism was to strongly induce peripheral tumor-specific T cells and to increase the accumulation of antigen-specific CD8 + T cells in the tumor [45]. Conversely, it has also been reported that elevated levels of gut microbiota metabolites, such as butyric acid and propionic acid, increase the proportion of Treg cells and decrease DC activation, which, in turn, leads to reduced effector T-cells and IL-2, and even tolerance to cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) blockers [48].
Additionally, Tfh cells are an important member of the adaptive immune family, and are present in mucosal lymphoid tissue and tumour-draining lymph nodes. Apoptosis of ileal crypt intestinal epithelial cells can induce Tfh cells to interfere with proximal colon tumors in an IL-12-dependent manner, inhibit the growth of tumor cells. This immune response depends on the microbiome of the ileum site, such as Bacteroides fragilis [49]. Th9 cells are important immune cells that secrete IL-9 in the intestinal lamina propria. If the expression of IL-4 and TGF-β and the number of Th9 cells in germ-free mice decrease, the probability of melanoma growth increases. After transplantation into germ-free mice, IL-9 production was restored and tumor growth was reduced [50]. Another study showed that Th9 cells exposed to staphylococcal enterotoxin B (ETB) significantly promoted apoptosis in tumor cells. As an antigen, ETB significantly increased the expression levels of STAT5 and HDAC-1 in CD4+ T-cells, resulting in increased IL-9 secretion [51].
DNA damage
Genotoxins released by gut microbiota exhibit DNase activity. Once released near gastrointestinal epithelial cells, these toxins cause double-stranded DNA to break in host epithelial cells, resulting in a brief arrest of the cell cycle. The first known mutagenic effect was observed in Escherichia coli. Colibactin from Escherichia coli can induce double-strand breaks by alkylating adenine residues in DNA, leading to direct mutations that can trigger CRC [52]. Since then, various studies have found that colibactin, cell-lethal bulking toxin, and BFT cause genetic mutations to varying degrees in colorectal, head and neck, urothelial, and other cancers [52,53,54,55] (Fig. 2).

Gut microbiota such as Shigella flexneri (S. flexneri), Escherichia coli (E. coli), Bacteroides fragilis (B. fragilis), Enterococcus faecalis (E. faecalis), Morganella morganii (M. morganii), and Helicobacter pylori (H. pylori) block the normal cell cycle by affecting oxidative environment-dependent DNA damage and disrupting the DNA mismatch-repair process, thus increasing the tendency of epithelial cells to become cancerous. Additionally, toxins secreted by E. coli interfere with histone acetylation, while butyrate and propionic acid, metabolites of Butyrivibrio fibrisolvens (B. fibrisolvens) and Propionibacterium, as inhibitors of deacetylase, can increase the degree of acetylation and have an opposite anticancer effect. E. faecalis releases oxidants through a macrophage-dependent manner, causing chromosomal instability.
Oxidation of the environment is an important cause of DNA damage. For example, analysis of ESR (electron spin resonance) spectra after Enterococcus faecalis colonization showed that E. faecalis produced extracellular superoxide and derived oxygen in a NOX2-dependent manner, which oxidizes the environment when diffused into the host cell and increases the likelihood of DNA mutations in the host cell [56]. Reactive oxygen species (ROS) production may be reduced by antibiotic use or intestinal sterility [28]. Similarly, Helicobacter pylori activate spermine oxidase in the host, producing large amounts of hydrogen peroxide and reactive oxygen species, inducing DNA mutation and carcinogenesis [57, 58]. SMO (spermine oxidase) is a metabolic enzyme induced by inflammatory signals. In colon cancer cells, BFT rapidly induced SMO gene expression, resulting in a 2- to 4-fold increase after 3 or 6 h of exposure, respectively, resulting in the production of SMO-dependent ROS and dysregulation of Gamma-H2AX, which further leads to DNA damage and induces carcinogenesis. Gamma-H2AX is the phosphorylated form of H2AX, involved in DNA repair when DNA breaks and cell cycle abnormalities occur [58] (Fig. 2).
As a tumor suppressive transcription factor, p53 can bind to specific DNA sequences and activate transcription, regulate unbalanced cell cycle, and repair defective genes. Common oncogenic p53 mutations usually occur when mediated by metabolites produced by the gut microbiota [59]. The protein CagA, produced by Helicobacter pylori, was the first bacterial protein to be shown to be associated with human cancer, impairing the repair process of DNA mismatches in gastrointestinal epithelial cells [60, 61]. CagA can interfere with the AKT pathway in host cells thereby promoting the occurrence of gastric cancer, after HCT116 cells were infected with the specified H. pylori strain for 10 h, western blot analysis showed a large degree of p53 degradation [62]. Similarly, Shigella flexneri also induces host cell p53 degradation through the secretases IpgD and VirA, increases the frequency of DNA mutations [63] (Fig. 2).
In addition, a large number of similar studies have assessed the ability of the gut microbiota to influence host DNA integrity. For example, a clinical study found that highly pathogenic mutations in the APC tumor suppressor gene in the intestinal cells of patients were associated with an increase in Fusobacterium mortiferum and a significant decrease in Clostridium geniculate and Bifidobacteria [64]. Morganella morganii produces a novel DNA toxic metabolite, indoleamine, which increases DNA mutation rate and intestinal permeability in mouse models of colon cancer and increases tumor load [65]. In SW480 cells transfected with a fusion gene containing 12 continuous cytosine residues, a significant increase in infection-induced cell point mutations of EPEC (Enteropathogenic E.coli) was observed compared to those for infection. Mechanistically, EPEC consumes host cell DNA mismatch-repair proteins by secreting EspF, which specifically targets the mitochondria of intestinal epithelial cells to induce their degradation [66] (Fig. 2).
In addition to DNA damage and variation, gut microbiota is also associated with chromosome stability, epigenome change, and microRNA, which we have detailed in the supplementary information (Supplementary information: supplement to the article).
Tumor-related signaling pathways
Sparassis latifolia polysaccharides (SLPs), secreted by Sparassis latifolia (a fungus of the genus Sparassis), has been shown to influence the progression of colorectal cancer in mice through multiple mechanisms, including inhibiting the infiltration of immune cells, reducing the expression of inflammatory cytokines, and improving the metabolic disorders of cancer cells [67]. Thus, in addition to participating in the immune response and DNA damage, communication between the gut microbiota and the host can occur through a variety of mechanisms, including receptor ligand-mediated signaling and regulation of intestinal epithelial barrier function, which often leads to cancer progression. In this section, we seek to explore the role of the gut microbiota in the signaling of tumor-related pathways, both promoting and inhibiting cancer (Fig. 3). In addition, the components and secreted products of intestinal flora, such as LPS and SCFAs, are also involved in the occurrence and development of tumors, which are discussed in the supplement (Supplementary information: supplement to the article).

CagA binds to SHP-2 and activates it to promote Ras/MAPK signaling and trigger the abnormal proliferation of host T-cells. Gingipains is the main virulence factor of Porphyromonas gingival, it up-regulate phosphorylation of MEK and ERK, the core components of the RAS/RAF/MEK/ERK pathway, promoting cancer cell proliferation. E-cadherin is a transmembrane glycoprotein that exists in cell membranes and binds epithelial cells together to maintain their normal morphology and polarity. Normally, the intracellular peptide of E-cadherin is linked to β-catenin to ensure that it does not transmit signals to the nucleus, while the intracellular free β-catenin exists in the form of a complex. CagA and FadA destroy the function of E-cadherin, affect the intercellular connection and the binding force of β-catenin, and switch on the WNT/β-catenin signal when the WNT signal activates the cell membrane receptor FRZ. TLR-4 recognizes bacterial metabolites, such as Bacteroides fragilis toxin (BFT), gingipains, lipopolysaccharides, and butyrate, and activates the NF-kB and NFAT signaling pathways to promote abnormal proliferation of cancer cells.
Inflammasomes
Inflammasomes are a class of multiprotein intracellular complexes expressed in immune and epithelial cells that induce cell death under pathological conditions such as inflammation and stress, and their disorders can lead to a variety of diseases, including autoimmune diseases and cancer. The outcome of inflammasome activation depends on a variety of factors, including its expression pattern and effector molecules. The gut microbiota may also influence the activation of specific inflammasomes [68, 69]. By activating the inflammasome, the gut microbiota interact with the immune and intestinal epithelial cells, producing results that can be both cancer-suppressing and cancer-proactive [70, 71]. In a state of homeostasis, intestinal commensal bacteria activate the NLRP3 inflammasome production of IL-18, regulating adaptive immunity, which is essential for maintaining the integrity of the intestinal barrier and preventing dysbacteriosis [72,73,74,75]. Experiments have shown that inflammasomes weaken the occurrence of colitis and colitis-related tumors in mouse models, the effector factor IL-18 plays a key role in this action [74, 76,77,78]. Conversely, by releasing IL-1β, inflammasome activation promotes the development of lung, skin, breast, and pancreatic cancers, a process in which the gut microbiota also play an important role [79,80,81,82,83].
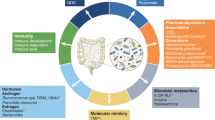
In addition to immune, genetic, cellular pathways, and inflammatory mediators, the gut microbiota also influence the development of hormone-dependent cancers. Patients with prostate cancer (PCa) often develop aggressive castration-resistant PCa because of alternative sources of androgens. Clostridium scindens is a member of the gut microbiota that converts cortisol into 11β-hydroxyl androstenedione, a potent androgen precursor. Cortisol metabolites derived from Clostridium scindens-conditioned medium promote the proliferation and migration of androgen-dependent PCa cells, which indicate that Clostridium scindens promotes PCa progression by activating androgen receptor signaling [84].
Moderating effect of gut microbiota on anticancer therapy
The combination of microbial and anticancer therapy began as early as the 19th century, when the first attempt was made to inject inactivated Streptococcus into human tumor tissues to cure cancer [85]. Previous studies have shown that microbial preparations injected directly into tumor tissues or administered orally can have a direct cytotoxic effect on tumor cells or stimulate the body’s local antitumor immune response. [86,87,88,89]. Recent studies have found that traditional radiotherapy, chemotherapy, and immunotherapy can alter the gut microbiota of patients, and that the composition of the flora can profoundly affect the efficacy and side effects of these treatments, including cancer recurrence, drug resistance, and collateral damage to the body (Table 2), we have analyzed this part of the content in detail and presented it in the supplementary information (Supplementary information: supplement to the article). Probiotics, prebiotics, and fecal microbiota transplantation (FMT) have also achieved some clinical efficacy; we will explain each of them below.
Probiotics
Probiotics are a class of bacteria that exist in the host and are beneficial to the host, and the purpose of administering probiotics to cancer patients is to reactivate the damaged gut microbiota of the patient, thereby reestablishing the level and function of the failed symbiotic microbiome [90, 91] (Table 3). Ingestion of adequate amounts of these microorganisms significantly improves intestinal crypt survival in mice and other animals by promoting the recovery of healthy gut microbiota and reducing apoptosis, in a protective effect that is partly dependent on TLR-2 and COX-2 [92]. The Lactobacillus rhamnosus GG strain (LGG) was the first probiotic studied in the field of oncology. Some previous studies found that LGG can directly regulate the host’s cell proliferation pathways, such as the mTOR or WNT pathways. LGG can also affect the host’s immune system and induce Th1 immune cell polarization through DC recognition, thereby enhancing the antitumor immune response and helping the host to remove newly formed cancer cells early [93,94,95,96,97]. Additionally, when patients were administered a bacterial mixture including two probiotics, Bifidobacterium longum (BB536) and Lactobacillus johnsonii (La1), these microorganisms were found to adhere to the colonic mucosa, reduce the concentration of intestinal pathogens, and regulate the local formation of an anti-cancer immune environment, as shown by significantly reduced proliferation of CD83-123, CD83-11c, and CD83-HLA-DR subsets in subjects receiving probiotics compared to controls [98].
In terms of mitigating the side effects of anti-cancer treatments, a 2015 clinical trial evaluated the preventive effect of a mixture of 10 different probiotic strains on gastrointestinal toxicity in patients with metastatic colorectal cancer receiving chemotherapy with irinotecan [99]. Microbial agents containing Lactobacillus acidophilus and Bifidobacterium longum significantly reduced moderate to severe diarrhea during pelvic radiotherapy [100]. The combination of probiotics in patients after CRC surgery may also relieve irritable bowel syndrome [101]. In a 2019 review, 15 studies showed that the combination of Bifidobacterium longum, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium infantis, and Saccharomycetes reduced the incidence of mucositis in patients who underwent radiation or chemotherapy [102].
Fecal flora transplantation
FMT from healthy donors has been used to repair dysbacteriosis in the gut and was described in TCM 1700 years ago [103]. In recent years, it has been increasingly used to treat various pathological processes [104,105,106,107]. Compared to oral probiotics, it is the most direct and rapid means of manipulating the gut microbiota, and can be administered to patients directly through oral freeze-dried capsules or via gastroscopy or colonoscopy [108,109,110,111]. In recent years, evidence has shown that FMT allows breakthroughs in the field of oncology treatment [112,113,114]. The effectiveness of FMT in reducing local and distant tumorigenesis in the gut was demonstrated in mouse models [115].
Although the clinical application of FMT is still in the experimental stage, it is effective for patients with acute myeloid leukemia and melanoma [116]. In multiple studies on melanoma patients, FMT has been found to alter the gut microbiota, reprogramming the TME by affecting the local immune system and inflammatory response to overcome PD-1-blocker resistance and improve its antitumor efficacy [117,118,119,120]. Hematopoietic stem cell transplantation (HSCT), as a relatively mature technology for the treatment of benign and malignant diseases of the blood system, often exposes patients to various complications, including recurrence, infection and graft-versa-host disease (GvHD), whish will lead to a great increase in patient mortality [121,122,123]. Several studies have shown that FMT is beneficial for GvHD remission in patients who received allogeneic HSCT. In a study of 15 patients who had received HSCT and developed GvHD, restoration of intestinal microbial diversity via FMT addressed steroid-resistant and steroid-dependent GvHD in the gut, with an increased abundance of beneficial bacteria and resolution of diarrhea [124]. Another eight-patient pilot study showed that GvHD patients who received FMT experienced relief of clinical symptoms, including changes in abdominal pain, diarrhea duration, and stool frequency. Gut microbiota composition was also reconstructed, including an increase in Bacteroidetes, Bacteroidaceae, Ruminococcaeae, and Desulfovibrionaceae [125].
Guiding role of gut microbiota in cancer diagnosis and prognosis
In addition to its important role in traditional cancer treatment, gut microbiota are also valuable in the diagnosis and prognosis of cancers such as CRC and LC. In terms of cancer diagnosis, a study in 2017 presented the metagenomic analysis of the CRC fecal microbiome to identify and validate bacterial biomarkers in different ethnic cohorts. This study included patients with CRC and control samples from China, Denmark, France, and Austria and highlighted the potential of fecal metagenomic biomarkers for early CRC diagnosis [126]. Since then, additional studies have found that fecal microbe DNA markers can be used as new tests to screen for colorectal tumors in asymptomatic subjects, either alone or in combination with fecal immunochemical tests. Furthermore, Zhao et al. collected fecal samples from 41 patients with LC and 40 healthy volunteers and analyzed the gut microbiota using 16 S rRNA gene sequencing. They found that Actinomyces, Veillonella, Megasphaera, Enterococcus, and Clostridioides were more abundant in patients with LC than in healthy individuals. They further demonstrate that gut microbes and their related metabolites as potential biomarkers and therapeutic targets for LC [127]. Gophna et al. examined changes in the gut microbiota and their potential as a biomarker in patients with pancreatic cancer. They compared the microbiomes of pancreatic patients with cancer with precancerous lesions, patients with non-alcoholic fatty liver disease, and healthy controls, and found unique pancreatic cancer-associated gut microbiota signatures. The predominant features were the presence of Clostridiacea, Lachnospiraceae, a lack of Ruminococcaceae, and excessive increases in Veillonellaceae, Akkermansia, and Odoribacter [128]. In conclusion, the gut microbiota profile may become a new effective marker for the early detection of cancer.
In terms of prognosis, many studies have demonstrated that the gut microbiota can be used as a potential prognostic marker of cancer. In a study of a prognostic model of patients with CRC based on age-related genes, Dai et al. found that the risk model was associated with immune status and the gut microbiota in patients with CRC, and that microbiome analysis showed a lower relative abundance of Bacteroidetes and Actinobacteria in high-risk patients than in low-risk patients. Combined with the results of consensus cluster analysis, Bacteroides enrichment in the gut has been associated with a poor prognosis in CRC patients [129]. Colov et al. found that high levels of Fusobacterium nucleatum and Bacteroides fragilis in the gut are associated with poor postoperative outcomes in patients with CRC [130]. The role of Fusobacterium nucleatum as a prognostic marker in patients with CRC has been demonstrated several times. For example, Yamaoka et al. collected 100 CRC tissues and 72 matched normal mucosal tissues and determined that levels of Fusobacterium nucleatum could help to predict clinical outcomes in CRC patients, stage IV CRC patients were found to have higher levels of Fusobacterium nucleatum [131, 132]. Additionally, Chung et al. demonstrated that specific gut microbiota were related to the prognosis of patients with hepatocellular carcinoma treated with nabuliumab. Specifically, the Prevotella/Bacteroides ratio can be used as a prognostic predictor for nivolumab treatment in hepatocellular carcinoma; the higher the ratio, the better the efficacy [133].
Diet, particularly prebiotics, mediates ecological changes in the gut and their association with cancer
The gut microbiota are an important part of the gut microecology. Diet may be the most powerful regulator of the microbiota in terms of its composition and metabolic function [134]. Previous studies have shown that people with diets rich in complex carbohydrates have significantly increased diversity of gut microbiota. Consumption of wheat-based bread improved body mass index and glucose tolerance, which were associated with Prevotella enrichment and increased polysaccharide fermentation capacity. Obese women who ingested a mixture rich in inulin and fructooligosaccharides over a period of time demonstrated enrichment of bacteria producing butyrate that lowered postprandial blood sugar levels. Compared with the high-calorie western diet, the Mediterranean diet is beneficial to the health of the gut microbiota and the host. The latter diet can increase the number of beneficial bacteria, including Lactobacillus and Enterococcus faecalis, promote an anti-inflammatory environment, reduce oxidative stress, particularly against breast cancer, gastric cancer, and upper gastrointestinal and respiratory cancers [135,136,137,138,139,140,141,142]. Yang et al. demonstrated that a high-fat diet often leads to an increased risk of CRC, which is related to ecological disorder of gut microbiota and intestinal barrier dysfunction, specifically manifested by an increase in Alistipessp Marseille P5997 and Alistipessp 5CPEGH6 in the intestine and a decrease in the probiotic Parabacteroides distasonis [143]. Compared with fat, dietary fiber derived from fruits, vegetables, and grains can change the density of gut microbiota, such as Firmicutes, improve antitumor immunity, and is negatively correlated with the risk of cancer [144].
Prebiotics are food components that are not digested and absorbed by the host and can selectively promote the metabolism and proliferation of probiotics in the gut. Common prebiotics include inulin, fructooligosaccharides, galactose, and some algae [145]. Prebiotics also play an important role in cancer development. A recent case–control study by Turati et al., which included 1,953 patients with histologically confirmed CRC and 4,154 controls, found that CRC risk was negatively associated with dietary intake of galactose [146]. Additionally, as the most common prebiotic, inulin increases the richness of Bifidobacteria, Bacteroides, and Akkermansia muciniphila in the intestinal tract of mice, and is associated with antitumor immunity [147]. In a recent study, Boucher et al. observed that a diet rich in inulin changes the gut microbiota, significantly promotes the growth of Bifidobacteria, and based on γδT lymphocyte tumor infiltration processes, also promotes immune control of tumor growth in melanoma, fibrosarcoma, and CRC in a mouse model [148].
Challenges and prospects
We have summarized the ongoing and completed clinical trials on gut microbiota in cancer treatment (Table 4). In the meantime, our results also raise controversial clinical questions such as how drugs (antibiotics) and environmental factors affect the composition and diversity of the gut microbiota, their interventional role in cancer treatment, and whether monitoring these factors during cancer treatment is necessary. Additionally, when it comes to improving the efficacy of cancer treatment responses by modulating the gut microbiota, it is not clear what composition of gut microbiota are the best for promoting antitumor immune responses. Further treatment options need to be carefully tested in clinical trials. It is also worth noting that most of the mechanisms by which the gut microbiota are involved in regulating cancer have been studied in mice, caution should be exercised when extrapolating these results to humans. Firstly, the intestinal structure and environment of mice and humans are different, as is the diversity of their flora. Secondly, the immune systems of mice and humans react differently to cancer. Thirdly, mice and humans have different diets and environmental exposures. Taken together, several factors contribute to the inability of the current knowledge to predict human clinical responses [149]. Therefore, in future studies, it is necessary to explore the safety, duration, dosage, dosage form, route of administration, combination of drugs, and other aspects of cancer treatment dependent on the gut microbiota more fully, to determine the best treatment plan for cancer.
Conclusion
Currently, more and more attention has been paid to the research on the composition and function of gut microbiota. Studies on different cancer types and experimental subjects have emphasized that gut microbiota play a dual role in cancer development. In addition, the use of gut microbiota in combination with traditional anti-tumor treatment strategies, as well as the use of probiotics, FMT, and dietary control, can improve the efficacy of anti-cancer treatment, while reducing the occurrence of side effects and improving prognosis. The value of gut microbiota for diagnosis and prognosis of cancer patients is also gradually being confirmed. In conclusion, the artificial control of gut microbiota can promote the development of cancer treatment system in the desired direction, which will provide a scientific basis for the development of more effective anti-cancer treatment programs and the promotion of precision medicine.
References
Zhang YJ, Li S, Gan RY, Zhou T, Xu DP, Li HB. Impacts of gut bacteria on human health and diseases. Int J Mol Sci. 2015;16:7493–519.
Wu Z, Tian E, Chen Y, Dong Z, Peng Q. Gut microbiota and its roles in the pathogenesis and therapy of endocrine system diseases. Microbiol Res. 2023;268:127291.
Sepich-Poore GD, Zitvogel L, Straussman R, Hasty J, Wargo JA, Knight R. The microbiome and human cancer. Science. 2021;371:eabc4552.
Fulbright LE, Ellermann M, Arthur JC. The microbiome and the hallmarks of cancer. PLoS Pathog. 2017;13:e1006480.
Rajagopala SV, Vashee S, Oldfield LM, Suzuki Y, Venter JC, Telenti A, et al. The human microbiome and cancer. Cancer Prev Res (Philos). 2017;10:226–34.
Knight R, Callewaert C, Marotz C, Hyde ER, Debelius JW, McDonald D, et al. The microbiome and human biology. Annu Rev Genomics Hum Genet. 2017;18:65–86.
Guinney J, Dienstmann R, Wang X, de Reyniès A, Schlicker A, Soneson C, et al. The consensus molecular subtypes of colorectal cancer. Nat Med. 2015;21:1350–6.
Purcell RV, Visnovska M, Biggs PJ, Schmeier S, Frizelle FA. Distinct gut microbiome patterns associate with consensus molecular subtypes of colorectal cancer. Sci Rep. 2017;7:11590.
Schwabe RF, Jobin C. The microbiome and cancer. Nat Rev Cancer. 2013;13:800–12.
Liu Y, Baba Y, Ishimoto T, Gu X, Zhang J, Nomoto D, et al. Gut microbiome in gastrointestinal cancer: a friend or foe? Int J Biol Sci. 2022;18:4101–17.
Wang WJ, Zhou YL, He J, Feng ZQ, Zhang L, Lai XB, et al. Characterizing the composition of intestinal microflora by 16S rRNA gene sequencing. World J Gastroenterol. 2020;26:614–26.
Durazzi F, Sala C, Castellani G, Manfreda G, Remondini D, De Cesare A. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci Rep. 2021;11:3030.
Kim H, Kim S, Jung S. Instruction of microbiome taxonomic profiling based on 16S rRNA sequencing. J Microbiol. 2020;58:193–205.
Laudadio I, Fulci V, Palone F, Stronati L, Cucchiara S, Carissimi C. Quantitative Assessment of Shotgun Metagenomics and 16S rDNA Amplicon Sequencing in the Study of Human Gut Microbiome. Omics. 2018;22:248–54.
Zhao M, Chen X, Yang Z, Yang X, Peng Q. Bacteria and tumor: Understanding the roles of bacteria in tumor genesis and immunology. Microbiol Res. 2022;261:127082.
Kuhn KA, Stappenbeck TS. Peripheral education of the immune system by the colonic microbiota. Semin Immunol. 2013;25:364–9.
Hanahan D. Hallmarks of cancer: new Dimensions. Cancer Discov. 2022;12:31–46.
Shetab Boushehri MA, Lamprecht A. TLR4-based immunotherapeutics in cancer: a review of the achievements and shortcomings. Mol Pharm. 2018;15:4777–800.
Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell. 2014;157:121–41.
Zitvogel L, Ma Y, Raoult D, Kroemer G, Gajewski TF. The microbiome in cancer immunotherapy: Diagnostic tools and therapeutic strategies. Science. 2018;359:1366–70.
Park J, Kim M, Kang SG, Jannasch AH, Cooper B, Patterson J, et al. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal Immunol. 2015;8:80–93.
Hatta MNA, Mohamad Hanif EA, Chin SF, Neoh HM. Pathogens and carcinogenesis: a review. Biology (Basel) 2021;10:533.
Lu Y, Yuan X, Wang M, He Z, Li H, Wang J, et al. Gut microbiota influence immunotherapy responses: mechanisms and therapeutic strategies. J Hematol Oncol. 2022;15:47.
Zheng D, Liwinski T, Elinav E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020;30:492–506.
Gur C, Ibrahim Y, Isaacson B, Yamin R, Abed J, Gamliel M, et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity. 2015;42:344–55.
Kostic AD, Chun E, Robertson L, Glickman JN, Gallini CA, Michaud M, et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe. 2013;14:207–15.
Ma C, Han M, Heinrich B, Fu Q, Zhang Q, Sandhu M, et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science. 2018;360.
Iida N, Dzutsev A, Stewart CA, Smith L, Bouladoux N, Weingarten RA, et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science. 2013;342:967–70.
Peled JU, Gomes ALC, Devlin SM, Littmann ER, Taur Y, Sung AD, et al. Microbiota as Predictor of Mortality in Allogeneic Hematopoietic-Cell Transplantation. N. Engl J Med. 2020;382:822–34.
Khosravi A, Yáñez A, Price JG, Chow A, Merad M, Goodridge HS, et al. Gut microbiota promote hematopoiesis to control bacterial infection. Cell Host Microbe. 2014;15:374–81.
Liu W, Chen Q, Wu S, Xia X, Wu A, Cui F, et al. Radioprotector WR-2721 and mitigating peptidoglycan synergistically promote mouse survival through the amelioration of intestinal and bone marrow damage. J Radiat Res. 2015;56:278–86.
Clarke TB, Davis KM, Lysenko ES, Zhou AY, Yu Y, Weiser JN. Recognition of peptidoglycan from the microbiota by Nod1 enhances systemic innate immunity. Nat Med. 2010;16:228–31.
Vicente-Dueñas C, Janssen S, Oldenburg M, Auer F, González-Herrero I, Casado-García A, et al. An intact gut microbiome protects genetically predisposed mice against leukemia. Blood. 2020;136:2003–17.
Fischer JC, Bscheider M, Eisenkolb G, Lin CC, Wintges A, Otten V, et al. RIG-I/MAVS and STING signaling promote gut integrity during irradiation- and immune-mediated tissue injury. Sci Transl Med. 2017;9:eaag2513.
Zitvogel L, Ayyoub M, Routy B, Kroemer G. Microbiome and Anticancer Immunosurveillance. Cell. 2016;165:276–87.
Ansaldo E, Slayden LC, Ching KL, Koch MA, Wolf NK, Plichta DR, et al. Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science. 2019;364:1179–84.
Weaver CT, Hatton RD, Mangan PR, Harrington LE. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu Rev Immunol. 2007;25:821–52.
Wu S, Rhee KJ, Albesiano E, Rabizadeh S, Wu X, Yen HR, et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat Med. 2009;15:1016–22.
Wu S, Powell J, Mathioudakis N, Kane S, Fernandez E, Sears CL. Bacteroides fragilis enterotoxin induces intestinal epithelial cell secretion of interleukin-8 through mitogen-activated protein kinases and a tyrosine kinase-regulated nuclear factor-kappaB pathway. Infect Immun. 2004;72:5832–9.
Pandey H, Tang DWT, Wong SH, Lal D. Gut microbiota in colorectal cancer: biological role and therapeutic opportunities. Cancers (Basel) 2023;15:866.
Mirji G, Worth A, Bhat SA, El Sayed M, Kannan T, Goldman AR, et al. The microbiome-derived metabolite TMAO drives immune activation and boosts responses to immune checkpoint blockade in pancreatic cancer. Sci Immunol. 2022;7:eabn0704.
Mager LF, Burkhard R, Pett N, Cooke NCA, Brown K, Ramay H, et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science. 2020;369:1481–9.
Uribe-Herranz M, Rafail S, Beghi S, Gil-de-Gómez L, Verginadis I, Bittinger K, et al. Gut microbiota modulate dendritic cell antigen presentation and radiotherapy-induced antitumor immune response. J Clin Invest. 2020;130:466–79.
Vétizou M, Pitt JM, Daillère R, Lepage P, Waldschmitt N, Flament C, et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015;350:1079–84.
Sivan A, Corrales L, Hubert N, Williams JB, Aquino-Michaels K, Earley ZM, et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science. 2015;350:1084–9.
Shi Y, Zheng W, Yang K, Harris KG, Ni K, Xue L, et al. Intratumoral accumulation of gut microbiota facilitates CD47-based immunotherapy via STING signaling. J Exp Med. 2020;217.
Li Y, Tinoco R, Elmén L, Segota I, Xian Y, Fujita Y, et al. Gut microbiota dependent anti-tumor immunity restricts melanoma growth in Rnf5(-/-) mice. Nat Commun. 2019;10:1492.
Nomura M, Nagatomo R, Doi K, Shimizu J, Baba K, Saito T, et al. Association of short-chain fatty acids in the gut microbiome with clinical response to treatment with nivolumab or pembrolizumab in patients with solid cancer tumors. JAMA Netw Open. 2020;3:e202895.
Roberti MP, Yonekura S, Duong CPM, Picard M, Ferrere G, Tidjani Alou M, et al. Chemotherapy-induced ileal crypt apoptosis and the ileal microbiome shape immunosurveillance and prognosis of proximal colon cancer. Nat Med. 2020;26:919–31.
Almeida RR, Vieira RS, Castoldi A, Terra FF, Melo ACL, Canesso MCC, et al. Host dysbiosis negatively impacts IL-9-producing T-cell differentiation and antitumour immunity. Br J Cancer. 2020;123:534–41.
Miao BP, Zhang RS, Sun HJ, Yu YP, Chen T, Li LJ, et al. Inhibition of squamous cancer growth in a mouse model by Staphylococcal enterotoxin B-triggered Th9 cell expansion. Cell Mol Immunol. 2017;14:371–9.
Wilson MR, Jiang Y, Villalta PW, Stornetta A, Boudreau PD, Carrá A, et al. The human gut bacterial genotoxin colibactin alkylates DNA. Science 2019;363:eaar7785.
Dejea CM, Fathi P, Craig JM, Boleij A, Taddese R, Geis AL, et al. Patients with familial adenomatous polyposis harbor colonic biofilms containing tumorigenic bacteria. Science. 2018;359:592–7.
He Z, Gharaibeh RZ, Newsome RC, Pope JL, Dougherty MW, Tomkovich S, et al. Campylobacter jejuni promotes colorectal tumorigenesis through the action of cytolethal distending toxin. Gut. 2019;68:289–300.
Boot A, Ng AWT, Chong FT, Ho SC, Yu W, Tan DSW, et al. Characterization of colibactin-associated mutational signature in an Asian oral squamous cell carcinoma and in other mucosal tumor types. Genome Res. 2020;30:803–13.
Huycke MM, Moore D, Joyce W, Wise P, Shepard L, Kotake Y, et al. Extracellular superoxide production by Enterococcus faecalis requires demethylmenaquinone and is attenuated by functional terminal quinol oxidases. Mol Microbiol. 2001;42:729–40.
Chaturvedi R, Asim M, Romero-Gallo J, Barry DP, Hoge S, de Sablet T, et al. Spermine oxidase mediates the gastric cancer risk associated with Helicobacter pylori CagA. Gastroenterology. 2011;141:1696–708.e1-2.
Goodwin AC, Destefano Shields CE, Wu S, Huso DL, Wu X, Murray-Stewart TR, et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proc Natl Acad Sci USA. 2011;108:15354–9.
Kadosh E, Snir-Alkalay I, Venkatachalam A, May S, Lasry A, Elyada E, et al. The gut microbiome switches mutant p53 from tumour-suppressive to oncogenic. Nature. 2020;586:133–8.
Hatakeyama M. Structure and function of Helicobacter pylori CagA, the first-identified bacterial protein involved in human cancer. Proc Jpn Acad Ser B Phys Biol Sci. 2017;93:196–219.
Kim JJ, Tao H, Carloni E, Leung WK, Graham DY, Sepulveda AR. Helicobacter pylori impairs DNA mismatch repair in gastric epithelial cells. Gastroenterology. 2002;123:542–53.
Buti L, Spooner E, Van der Veen AG, Rappuoli R, Covacci A, Ploegh HL. Helicobacter pylori cytotoxin-associated gene A (CagA) subverts the apoptosis-stimulating protein of p53 (ASPP2) tumor suppressor pathway of the host. Proc Natl Acad Sci USA. 2011;108:9238–43.
Bergounioux J, Elisee R, Prunier AL, Donnadieu F, Sperandio B, Sansonetti P, et al. Calpain activation by the Shigella flexneri effector VirA regulates key steps in the formation and life of the bacterium’s epithelial niche. Cell Host Microbe. 2012;11:240–52.
Liang S, Mao Y, Liao M, Xu Y, Chen Y, Huang X, et al. Gut microbiome associated with APC gene mutation in patients with intestinal adenomatous polyps. Int J Biol Sci. 2020;16:135–46.
Cao Y, Oh J, Xue M, Huh WJ, Wang J, Gonzalez-Hernandez JA, et al. Commensal microbiota from patients with inflammatory bowel disease produce genotoxic metabolites. Science. 2022;378:eabm3233.
Maddocks OD, Scanlon KM, Donnenberg MS. An Escherichia coli effector protein promotes host mutation via depletion of DNA mismatch repair proteins. mBio. 2013;4:e00152–13.
Wei X, Cheng F, Liu J, Cheng Y, Yun S, Meng J, et al. Sparassis latifolia polysaccharides inhibit colon cancer in mice by modulating gut microbiota and metabolism. Int J Biol Macromol. 2023;232:123299.
Levy M, Thaiss CA, Katz MN, Suez J, Elinav E. Inflammasomes and the microbiota-partners in the preservation of mucosal homeostasis. Semin Immunopathol. 2015;37:39–46.
Lara-Reyna S, Caseley EA, Topping J, Rodrigues F, Jimenez Macias J, Lawler SE, et al. Inflammasome activation: from molecular mechanisms to autoinflammation. Clin Transl Immunol. 2022;11:e1404.
Zitvogel L, Kepp O, Galluzzi L, Kroemer G. Inflammasomes in carcinogenesis and anticancer immune responses. Nat Immunol. 2012;13:343–51.
Kantono M, Guo B. Inflammasomes and Cancer: The Dynamic Role of the Inflammasome in Tumor Development. Front Immunol. 2017;8:1132.
Brunetto E, De Monte L, Balzano G, Camisa B, Laino V, Riba M, et al. The IL-1/IL-1 receptor axis and tumor cell released inflammasome adaptor ASC are key regulators of TSLP secretion by cancer associated fibroblasts in pancreatic cancer. J Immunother Cancer. 2019;7:45.
Ershaid N, Sharon Y, Doron H, Raz Y, Shani O, Cohen N, et al. NLRP3 inflammasome in fibroblasts links tissue damage with inflammation in breast cancer progression and metastasis. Nat Commun. 2019;10:4375.
Wen J, Xuan B, Liu Y, Wang L, He L, Meng X, et al. NLRP3 inflammasome-induced pyroptosis in digestive system tumors. Front Immunol. 2023;14:1074606.
Zhang Z, Li X, Wang Y, Wei Y, Wei X. Involvement of inflammasomes in tumor microenvironment and tumor therapies. J Hematol Oncol. 2023;16:24.
Allen IC, TeKippe EM, Woodford RM, Uronis JM, Holl EK, Rogers AB, et al. The NLRP3 inflammasome functions as a negative regulator of tumorigenesis during colitis-associated cancer. J Exp Med. 2010;207:1045–56.
Chen GY, Liu M, Wang F, Bertin J, Núñez G. A functional role for Nlrp6 in intestinal inflammation and tumorigenesis. J Immunol. 2011;186:7187–94.
Williams TM, Leeth RA, Rothschild DE, Coutermarsh-Ott SL, McDaniel DK, Simmons AE, et al. The NLRP1 inflammasome attenuates colitis and colitis-associated tumorigenesis. J Immunol. 2015;194:3369–80.
McLoed AG, Sherrill TP, Cheng DS, Han W, Saxon JA, Gleaves LA, et al. Neutrophil-Derived IL-1β Impairs the Efficacy of NF-κB Inhibitors against Lung Cancer. Cell Rep. 2016;16:120–32.
Okamoto M, Liu W, Luo Y, Tanaka A, Cai X, Norris DA, et al. Constitutively active inflammasome in human melanoma cells mediating autoinflammation via caspase-1 processing and secretion of interleukin-1beta. J Biol Chem. 2010;285:6477–88.
Zhong FL, Mamaï O, Sborgi L, Boussofara L, Hopkins R, Robinson K, et al. Germline NLRP1 Mutations Cause Skin Inflammatory and Cancer Susceptibility Syndromes via Inflammasome Activation. Cell. 2016;167:187–202.e17.
Kolb R, Phan L, Borcherding N, Liu Y, Yuan F, Janowski AM, et al. Obesity-associated NLRC4 inflammasome activation drives breast cancer progression. Nat Commun. 2016;7:13007.
Daley D, Mani VR, Mohan N, Akkad N, Pandian G, Savadkar S, et al. NLRP3 signaling drives macrophage-induced adaptive immune suppression in pancreatic carcinoma. J Exp Med. 2017;214:1711–24.
Bui NN, Li CY, Wang LY, Chen YA, Kao WH, Chou LF, et al. Clostridium scindens metabolites trigger prostate cancer progression through androgen receptor signaling. J Microbiol Immunol Infect. 2023;56:246–56.
Nauts HC, Swift WE, Coley BL. The treatment of malignant tumors by bacterial toxins as developed by the late William B. Coley, M.D., reviewed in the light of modern research. Cancer Res. 1946;6:205–16.
Zbar B, Bernstein I, Tanaka T, Rapp HJ. Tumor immunity produced by the intradermal inoculation of living tumor cells and living Mycobacterium bovis (strain BCG). Science. 1970;170:1217–8.
Aso Y, Akazan H. Prophylactic effect of a Lactobacillus casei preparation on the recurrence of superficial bladder cancer. BLP Study Group. Urol Int. 1992;49:125–9.
Hoesl CE, Altwein JE. The probiotic approach: an alternative treatment option in urology. Eur Urol. 2005;47:288–96.
Stebbing J, Dalgleish A, Gifford-Moore A, Martin A, Gleeson C, Wilson G, et al. An intra-patient placebo-controlled phase I trial to evaluate the safety and tolerability of intradermal IMM-101 in melanoma. Ann Oncol. 2012;23:1314–9.
Kim SK, Guevarra RB, Kim YT, Kwon J, Kim H, Cho JH, et al. Role of Probiotics in Human Gut Microbiome-Associated Diseases. J Microbiol Biotechnol. 2019;29:1335–40.
Wilkins T, Sequoia J. Probiotics for Gastrointestinal Conditions: A Summary of the Evidence. Am Fam Physician. 2017;96:170–8.
Ciorba MA, Riehl TE, Rao MS, Moon C, Ee X, Nava GM, et al. Lactobacillus probiotic protects intestinal epithelium from radiation injury in a TLR-2/cyclo-oxygenase-2-dependent manner. Gut. 2012;61:829–38.
Khailova L, Baird CH, Rush AA, Barnes C, Wischmeyer PE. Lactobacillus rhamnosus GG treatment improves intestinal permeability and modulates inflammatory response and homeostasis of spleen and colon in experimental model of Pseudomonas aeruginosa pneumonia. Clin Nutr. 2017;36:1549–57.
Banna GL, Torino F, Marletta F, Santagati M, Salemi R, Cannarozzo E, et al. Lactobacillus rhamnosus GG: An Overview to Explore the Rationale of Its Use in Cancer. Front Pharm. 2017;8:603.
Taherian-Esfahani Z, Abedin-Do A, Nouri Z, Mirfakhraie R, Ghafouri-Fard S, Motevaseli E. Lactobacilli Differentially Modulate mTOR and Wnt/ β-Catenin Pathways in Different Cancer Cell Lines. Iran J Cancer Prev. 2016;9:e5369.
Cai S, Kandasamy M, Rahmat JN, Tham SM, Bay BH, Lee YK, et al. Lactobacillus rhamnosus GG Activation of Dendritic Cells and Neutrophils Depends on the Dose and Time of Exposure. J Immunol Res. 2016;2016:7402760.
Salemi R, Vivarelli S, Ricci D, Scillato M, Santagati M, Gattuso G, et al. Lactobacillus rhamnosus GG cell-free supernatant as a novel anti-cancer adjuvant. J Transl Med. 2023;21:195.
Gianotti L, Morelli L, Galbiati F, Rocchetti S, Coppola S, Beneduce A, et al. A randomized double-blind trial on perioperative administration of probiotics in colorectal cancer patients. World J Gastroenterol. 2010;16:167–75.
Mego M, Chovanec J, Vochyanova-Andrezalova I, Konkolovsky P, Mikulova M, Reckova M, et al. Prevention of irinotecan induced diarrhea by probiotics: A randomized double blind, placebo controlled pilot study. Complement Ther Med. 2015;23:356–62.
Demers M, Dagnault A, Desjardins J. A randomized double-blind controlled trial: impact of probiotics on diarrhea in patients treated with pelvic radiation. Clin Nutr. 2014;33:761–7.
Theodoropoulos GE, Memos NA, Peitsidou K, Karantanos T, Spyropoulos BG, Zografos G. Synbiotics and gastrointestinal function-related quality of life after elective colorectal cancer resection. Ann Gastroenterol. 2016;29:56–62.
Picó-Monllor JA, Mingot-Ascencao JM. Search and selection of probiotics that improve mucositis symptoms in oncologic patients. A Syst Rev Nutrients. 2019;11:2322.
Zhang F, Luo W, Shi Y, Fan Z, Ji G. Should we standardize the 1,700-year-old fecal microbiota transplantation? Am J Gastroenterol. 2012;107:1755.
Baunwall SMD, Terveer EM, Dahlerup JF, Erikstrup C, Arkkila P, Vehreschild MJ, et al. The use of Faecal Microbiota Transplantation (FMT) in Europe: A Europe-wide survey. Lancet Reg Health Eur. 2021;9:100181.
El-Salhy M, Hatlebakk JG, Gilja OH, Bråthen Kristoffersen A, Hausken T. Efficacy of faecal microbiota transplantation for patients with irritable bowel syndrome in a randomised, double-blind, placebo-controlled study. Gut. 2020;69:859–67.
Ghani R, Mullish BH, McDonald JAK, Ghazy A, Williams HRT, Brannigan ET, et al. Disease Prevention Not Decolonization: A Model for Fecal Microbiota Transplantation in Patients Colonized With Multidrug-resistant Organisms. Clin Infect Dis. 2021;72:1444–7.
Makkawi S, Camara-Lemarroy C, Metz L. Fecal microbiota transplantation associated with 10 years of stability in a patient with SPMS. Neurol Neuroimmunol Neuroinflamm. 2018;5:e459.
Chin SM, Sauk J, Mahabamunuge J, Kaplan JL, Hohmann EL, Khalili H. Fecal Microbiota Transplantation for Recurrent Clostridium difficile Infection in Patients With Inflammatory Bowel Disease: A Single-Center Experience. Clin Gastroenterol Hepatol. 2017;15:597–9.
van Nood E, Vrieze A, Nieuwdorp M, Fuentes S, Zoetendal EG, de Vos WM, et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N. Engl J Med. 2013;368:407–15.
Orenstein R, Dubberke E, Hardi R, Ray A, Mullane K, Pardi DS, et al. Safety and Durability of RBX2660 (Microbiota Suspension) for Recurrent Clostridium difficile Infection: Results of the PUNCH CD Study. Clin Infect Dis. 2016;62:596–602.
Kao D, Roach B, Silva M, Beck P, Rioux K, Kaplan GG, et al. Effect of Oral Capsule- vs Colonoscopy-Delivered Fecal Microbiota Transplantation on Recurrent Clostridium difficile Infection: A Randomized Clinical Trial. Jama. 2017;318:1985–93.
Xu H, Cao C, Ren Y, Weng S, Liu L, Guo C, et al. Antitumor effects of fecal microbiota transplantation: Implications for microbiome modulation in cancer treatment. Front Immunol. 2022;13:949490.
Baruch EN, Youngster I, Ben-Betzalel G, Ortenberg R, Lahat A, Katz L, et al. Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients. Science. 2021;371:602–9.
Hindson J. FMT for immunotherapy-refractory melanoma. Nat Rev Gastroenterol Hepatol. 2021;18:82.
Bel S, Elkis Y, Elifantz H, Koren O, Ben-Hamo R, Lerer-Goldshtein T, et al. Reprogrammed and transmissible intestinal microbiota confer diminished susceptibility to induced colitis in TMF-/- mice. Proc Natl Acad Sci USA. 2014;111:4964–9.
Gopalakrishnan V, Helmink BA, Spencer CN, Reuben A, Wargo JA. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell. 2018;33:570–80.
Davar D, Dzutsev AK, McCulloch JA, Rodrigues RR, Chauvin JM, Morrison RM, et al. Fecal microbiota transplant overcomes resistance to anti-PD-1 therapy in melanoma patients. Science. 2021;371:595–602.
Matson V, Fessler J, Bao R, Chongsuwat T, Zha Y, Alegre ML, et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science. 2018;359:104–8.
Gopalakrishnan V, Spencer CN, Nezi L, Reuben A, Andrews MC, Karpinets TV, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science. 2018;359:97–103.
Routy B, Le Chatelier E, Derosa L, Duong CPM, Alou MT, Daillère R, et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. 2018;359:91–7.
Galgano L, Hutt D. HSCT: How Does It Work?, in The European Blood and Marrow Transplantation Textbook for Nurses: Under the Auspices of EBMT, M Kenyon and A Babic, Editors. (2018), Springer Copyright 2018, EBMT and the Author(s). Cham (CH). 23–36.
Koyama M, Kuns RD, Olver SD, Raffelt NC, Wilson YA, Don AL, et al. Recipient nonhematopoietic antigen-presenting cells are sufficient to induce lethal acute graft-versus-host disease. Nat Med. 2011;18:135–42.
Ciernikova S, Kasperova B, Drgona L, Smolkova B, Stevurkova V, Mego M. Targeting the gut microbiome: An emerging trend in hematopoietic stem cell transplantation. Blood Rev. 2021;48:100790.
van Lier YF, Davids M, Haverkate NJE, de Groot PF, Donker ML, Meijer E, et al. Donor fecal microbiota transplantation ameliorates intestinal graft-versus-host disease in allogeneic hematopoietic cell transplant recipients. Sci Transl Med. 2020;12:eaaz8926.
Qi X, Li X, Zhao Y, Wu X, Chen F, Ma X, et al. Treating Steroid Refractory Intestinal Acute Graft-vs.-Host Disease With Fecal Microbiota Transplantation: A Pilot Study. Front Immunol. 2018;9:2195.
Yu J, Feng Q, Wong SH, Zhang D, Liang QY, Qin Y, et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut. 2017;66:70–8.
Zhao F, An R, Wang L, Shan J, Wang X. Specific Gut Microbiome and Serum Metabolome Changes in Lung Cancer Patients. Front Cell Infect Microbiol. 2021;11:725284.
Half E, Keren N, Reshef L, Dorfman T, Lachter I, Kluger Y, et al. Fecal microbiome signatures of pancreatic cancer patients. Sci Rep. 2019;9:16801.
Dai JJ, Fu YY, Zhong XQ, Cen W, Ye MF, Chen XH, et al. Identification of Senescence-Related Subtypes, the Development of a Prognosis Model, and Characterization of Immune Infiltration and Gut Microbiota in Colorectal Cancer. Front Med (Lausanne). 2022;9:916565.
Colov EP, Degett TH, Raskov H, Gögenur I. The impact of the gut microbiota on prognosis after surgery for colorectal cancer—a systematic review and meta-analysis. Apmis. 2020;128:162–76.
Yamaoka Y, Suehiro Y, Hashimoto S, Hoshida T, Fujimoto M, Watanabe M, et al. Fusobacterium nucleatum as a prognostic marker of colorectal cancer in a Japanese population. J Gastroenterol. 2018;53:517–24.
Gethings-Behncke C, Coleman HG, Jordao HWT, Longley DB, Crawford N, Murray LJ, et al. Fusobacterium nucleatum in the Colorectum and Its Association with Cancer Risk and Survival: A Systematic Review and Meta-analysis. Cancer Epidemiol Biomark Prev. 2020;29:539–48.
Chung MW, Kim MJ, Won EJ, Lee YJ, Yun YW, Cho SB, et al. Gut microbiome composition can predict the response to nivolumab in advanced hepatocellular carcinoma patients. World J Gastroenterol. 2021;27:7340–9.
Wastyk HC, Fragiadakis GK, Perelman D, Dahan D, Merrill BD, Yu FB, et al. Gut-microbiota-targeted diets modulate human immune status. Cell. 2021;184:4137–4153.e14.
Schnorr SL, Candela M, Rampelli S, Centanni M, Consolandi C, Basaglia G, et al. Gut microbiome of the Hadza hunter-gatherers. Nat Commun. 2014;5:3654.
Koh A, De Vadder F, Kovatcheva-Datchary P, Bäckhed F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell. 2016;165:1332–45.
Sonnenburg ED, Smits SA, Tikhonov M, Higginbottom SK, Wingreen NS, Sonnenburg JL. Diet-induced extinctions in the gut microbiota compound over generations. Nature. 2016;529:212–5.
Kovatcheva-Datchary P, Nilsson A, Akrami R, Lee YS, De Vadder F, Arora T, et al. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015;22:971–82.
Arpaia N, Campbell C, Fan X, Dikiy S, van der Veeken J, deRoos P, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature. 2013;504:451–5.
Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia. 2007;50:2374–83.
Nagpal R, Shively CA, Register TC, Craft S, Yadav H. Gut microbiome-Mediterranean diet interactions in improving host health. F1000Res. 2019;8:699.
Del Chierico F, Vernocchi P, Dallapiccola B, Putignani L. Mediterranean diet and health: food effects on gut microbiota and disease control. Int J Mol Sci. 2014;15:11678–99.
Yang J, Wei H, Zhou Y, Szeto CH, Li C, Lin Y, et al. High-Fat Diet Promotes Colorectal Tumorigenesis Through Modulating Gut Microbiota and Metabolites. Gastroenterology. 2022;162:135–49.e2.
Yang J, Yu J. The association of diet, gut microbiota and colorectal cancer: what we eat may imply what we get. Protein Cell. 2018;9:474–87.
Martyniak A, Medyńska-Przęczek A, Wędrychowicz A, Skoczeń S and Tomasik PJ, Prebiotics, Probiotics, Synbiotics, Paraprobiotics and Postbiotic Compounds in IBD. Biomolecules 2021;11:1903.
Turati F, Concina F, Rossi M, Fiori F, Parpinel M, Taborelli M, et al. Association of prebiotic fiber intake with colorectal cancer risk: the PrebiotiCa study. Eur J Nutr. 2023;62:455–64.
Li Y, Elmén L, Segota I, Xian Y, Tinoco R, Feng Y, et al. Prebiotic-Induced Anti-tumor Immunity Attenuates Tumor Growth. Cell Rep. 2020;30:1753–66.e6.
Boucher E, Plazy C, Richard ML, Suau A, Mangin I, Cornet M, et al. Inulin prebiotic reinforces host cancer immunosurveillance via ɣδ T cell activation. Front Immunol. 2023;14:1104224.
Fernandes MR, Aggarwal P, Costa RGF, Cole AM, Trinchieri G. Targeting the gut microbiota for cancer therapy. Nat Rev Cancer. 2022;22:703–22.
Rubinstein MR, Wang X, Liu W, Hao Y, Cai G, Han YW. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe. 2013;14:195–206.
Nomoto D, Baba Y, Liu Y, Tsutsuki H, Okadome K, Harada K, et al. Fusobacterium nucleatum promotes esophageal squamous cell carcinoma progression via the NOD1/RIPK2/NF-κB pathway. Cancer Lett. 2022;530:59–67.
Udayasuryan B, Ahmad RN, Nguyen TTD, Umaña A, Monét Roberts L, Sobol P, et al. Fusobacterium nucleatum induces proliferation and migration in pancreatic cancer cells through host autocrine and paracrine signaling. Sci Signal. 2022;15:eabn4948.
Kim JM, Cho SJ, Oh YK, Jung HY, Kim YJ, Kim N. Nuclear factor-kappa B activation pathway in intestinal epithelial cells is a major regulator of chemokine gene expression and neutrophil migration induced by Bacteroides fragilis enterotoxin. Clin Exp Immunol. 2002;130:59–66.
Wang L, Tang L, Feng Y, Zhao S, Han M, Zhang C, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurised bacterium blunts colitis associated tumourigenesis by modulation of CD8(+) T cells in mice. Gut. 2020;69:1988–97.
Takagi A, Ikemura H, Matsuzaki T, Sato M, Nomoto K, Morotomi M, et al. Relationship between the in vitro response of dendritic cells to Lactobacillus and prevention of tumorigenesis in the mouse. J Gastroenterol. 2008;43:661–9.
Lenoir M, Del Carmen S, Cortes-Perez NG, Lozano-Ojalvo D, Muñoz-Provencio D, Chain F, et al. Lactobacillus casei BL23 regulates Treg and Th17 T-cell populations and reduces DMH-associated colorectal cancer. J Gastroenterol. 2016;51:862–73.
Taieb F, Nougayrède JP, Watrin C, Samba-Louaka A, Oswald E. Escherichia coli cyclomodulin Cif induces G2 arrest of the host cell cycle without activation of the DNA-damage checkpoint-signalling pathway. Cell Microbiol. 2006;8:1910–21.
Cuevas-Ramos G, Petit CR, Marcq I, Boury M, Oswald E, Nougayrède JP. Escherichia coli induces DNA damage in vivo and triggers genomic instability in mammalian cells. Proc Natl Acad Sci USA. 2010;107:11537–42.
Murata-Kamiya N, Kurashima Y, Teishikata Y, Yamahashi Y, Saito Y, Higashi H, et al. Helicobacter pylori CagA interacts with E-cadherin and deregulates the beta-catenin signal that promotes intestinal transdifferentiation in gastric epithelial cells. Oncogene. 2007;26:4617–26.
Chu FF, Esworthy RS, Chu PG, Longmate JA, Huycke MM, Wilczynski S, et al. Bacteria-induced intestinal cancer in mice with disrupted Gpx1 and Gpx2 genes. Cancer Res. 2004;64:962–8.
Huycke MM, Abrams V, Moore DR. Enterococcus faecalis produces extracellular superoxide and hydrogen peroxide that damages colonic epithelial cell DNA. Carcinogenesis. 2002;23:529–36.
Donohoe DR, Holley D, Collins LB, Montgomery SA, Whitmore AC, Hillhouse A, et al. A gnotobiotic mouse model demonstrates that dietary fiber protects against colorectal tumorigenesis in a microbiota- and butyrate-dependent manner. Cancer Discov. 2014;4:1387–97.
Mu W, Jia Y, Chen X, Li H, Wang Z, Cheng B. Intracellular Porphyromonas gingivalis Promotes the Proliferation of Colorectal Cancer Cells via the MAPK/ERK Signaling Pathway. Front Cell Infect Microbiol. 2020;10:584798.
Kim OY, Park HT, Dinh NTH, Choi SJ, Lee J, Kim JH, et al. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat Commun. 2017;8:626.
Konishi H, Fujiya M, Tanaka H, Ueno N, Moriichi K, Sasajima J, et al. Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis. Nat Commun. 2016;7:12365.
Riaz Rajoka MS, Zhao H, Lu Y, Lian Z, Li N, Hussain N, et al. Anticancer potential against cervix cancer (HeLa) cell line of probiotic Lactobacillus casei and Lactobacillus paracasei strains isolated from human breast milk. Food Funct. 2018;9:2705–15.
Zununi Vahed S, Barzegari A, Rahbar Saadat Y, Goreyshi A, Omidi Y. Leuconostoc mesenteroides-derived anticancer pharmaceuticals hinder inflammation and cell survival in colon cancer cells by modulating NF-κB/AKT/PTEN/MAPK pathways. Biomed Pharmacother. 2017;94:1094–100.
Behzadi E, Mahmoodzadeh Hosseini H, Imani Fooladi AA. The inhibitory impacts of Lactobacillus rhamnosus GG-derived extracellular vesicles on the growth of hepatic cancer cells. Micro Pathog. 2017;110:1–6.
Cheng Z, Xu H, Wang X, Liu Z. Lactobacillus raises in vitro anticancer effect of geniposide in HSC-3 human oral squamous cell carcinoma cells. Exp Ther Med. 2017;14:4586–94.
Zhao BB, Meng J, Zhang QX, Kang TT, Lu RR. Protective effect of surface layer proteins isolated from four Lactobacillus strains on hydrogen-peroxide-induced HT-29 cells oxidative stress. Int J Biol Macromol. 2017;102:76–83.
Viaud S, Saccheri F, Mignot G, Yamazaki T, Daillère R, Hannani D, et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science. 2013;342:971–6.
Gui QF, Lu HF, Zhang CX, Xu ZR, Yang YH. Well-balanced commensal microbiota contributes to anti-cancer response in a lung cancer mouse model. Genet Mol Res. 2015;14:5642–51.
An J, Ha EM. Combination Therapy of Lactobacillus plantarum Supernatant and 5-Fluouracil Increases Chemosensitivity in Colorectal Cancer Cells. J Microbiol Biotechnol. 2016;26:1490–503.
Nomoto K, Yokokura T, Mitsuyama M, Yoshikai Y, Nomoto K. Prevention of indigenous infection of mice with Escherichia coli by nonspecific immunostimulation. Antimicrob Agents Chemother. 1992;36:361–7.
Yu T, Guo F, Yu Y, Sun T, Ma D, Han J, et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell. 2017;170:548–63.e16.
Geller LT, Barzily-Rokni M, Danino T, Jonas OH, Shental N, Nejman D, et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science. 2017;357:1156–60.
Chaput N, Lepage P, Coutzac C, Soularue E, Le Roux K, Monot C, et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann Oncol. 2017;28:1368–79.
Lam KC, Araya RE, Huang A, Chen Q, Di Modica M, Rodrigues RR, et al. Microbiota triggers STING-type I IFN-dependent monocyte reprogramming of the tumor microenvironment. Cell. 2021;184:5338–56.e21.
Chitapanarux I, Chitapanarux T, Traisathit P, Kudumpee S, Tharavichitkul E, Lorvidhaya V. Randomized controlled trial of live lactobacillus acidophilus plus bifidobacterium bifidum in prophylaxis of diarrhea during radiotherapy in cervical cancer patients. Radiat Oncol. 2010;5:31.
Hibberd AA, Lyra A, Ouwehand AC, Rolny P, Lindegren H, Cedgård L, et al. Intestinal microbiota is altered in patients with colon cancer and modified by probiotic intervention. BMJ Open Gastroenterol. 2017;4:e000145.
Yamamoto ML, Maier I, Dang AT, Berry D, Liu J, Ruegger PM, et al. Intestinal bacteria modify lymphoma incidence and latency by affecting systemic inflammatory state, oxidative stress, and leukocyte genotoxicity. Cancer Res. 2013;73:4222–32.
de Moreno de LeBlanc A, Matar C, Thériault C, Perdigón G. Effects of milk fermented by Lactobacillus helveticus R389 on immune cells associated to mammary glands in normal and a breast cancer model. Immunobiology. 2005;210:349–58.
Jacouton E, Chain F, Sokol H, Langella P, Bermúdez-Humarán LG. Probiotic Strain Lactobacillus casei BL23 Prevents Colitis-Associated Colorectal Cancer. Front Immunol. 2017;8:1553.
Acknowledgements
We would like to thank Editage (www.editage.cn) for English language editing.
Funding
This project is supported by Dalian Young Stars of Science and Technology Fund (No. 2022-RQ-010).
Author information
Authors and Affiliations
Contributions
SJ was the first author, to collect data, drafting and revising the manuscript. CF and WG design research direction provide writing guidance and participate in the manuscript revision and supplement. All authors have agreed to the version of the manuscript for this release and have agreed to work on their respective aspects.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sun, J., Chen, F. & Wu, G. Potential effects of gut microbiota on host cancers: focus on immunity, DNA damage, cellular pathways, and anticancer therapy. ISME J 17, 1535–1551 (2023). https://doi.org/10.1038/s41396-023-01483-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41396-023-01483-0
