
Abstract
Early-life stress (ELS) leaves signatures upon the brain that persist throughout the lifespan and increase the risk of psychiatric illnesses including mood and anxiety disorders. In humans, myriad forms of ELS—including childhood abuse, bullying, poverty, and trauma—are increasingly prevalent. Understanding the signs of ELS, including those associated with psychiatric illness, will enable improved treatment and prevention. Here, we developed a novel procedure to model human ELS in mice and identify translationally-relevant biomarkers of mood and anxiety disorders. We exposed male mice (C57BL/6 J) to an early-life (juvenile) chronic social defeat stress (jCSDS) and examined social interaction and responsivity to reward during adulthood. As expected, jCSDS-exposed mice showed a socially avoidant phenotype in open-field social interaction tests. However, sucrose preference tests failed to demonstrate ELS-induced reductions in choice for the sweetened solution, suggesting no effect on reward function. To explore whether other tasks might be more sensitive to changes in motivation, we tested the mice in the Probabilistic Reward Task (PRT), a procedure often used in humans to study reward learning deficits associated with depressive illness. In a touchscreen PRT variant that was reverse-translated to maximize alignment with the version used in human subjects, mice exposed to jCSDS displayed significant reductions in the tendency to develop response biases for the more richly-rewarded stimulus, a hallmark sign of anhedonia when observed in humans. Our findings suggest that translationally-relevant procedures that utilize the same endpoints across species may enable the development of improved model systems that more accurately predict outcomes in humans.
Similar content being viewed by others

Introduction
Early life stress (ELS) is a primary risk factor for developing mood and anxiety disorders including Major Depressive Disorder, Generalized Anxiety Disorder, and Post-Traumatic Stress Disorder [1, 2]. In humans, ELS—which encompasses many conditions including child abuse, bullying, neglect, and poverty/low resource environments—affects as many as 1 in 7 children a year and is increasingly associated with alterations in brain connectivity and anatomy during adolescence and adulthood [1,2,3]. Interestingly, studies in laboratory animals indicate that different forms of ELS can produce profoundly different neurobiological alterations. As only one example, early life neglect can result in precocious maturation of prefrontal amygdala circuitry, whereas early life physical abuse instead delays maturation of this same circuitry [4, 5]. Considering that the effects of ELS on the brain seem so dependent on the specific attributes of the stressful experience, it is critical to explore multiple forms of ELS in model systems that are sensitive to key diagnostic features of mood and anxiety disorders, such as social withdrawal and decreased sensitivity to reward (anhedonia).
While numerous rodent models of early life neglect [6, 7] as well as caregiver maltreatment in infants [8, 9] have been developed, there are relatively few reports describing the effects of early life trauma with a physical component [10,11,12] in post-weaning age mice. Chronic social defeat stress (CSDS) in adult rodents is a commonly used model of stress that produces deficits in endpoints including social interaction, motivation, and sleep [13,14,15,16]. Complex behavioral tasks such as intracranial self-stimulation (ICSS) clearly demonstrate that CSDS produces anhedonia [17], but that procedure requires surgical intervention, long periods of training (weeks to months), and a substantial equipment infrastructure while having no complementary paradigm in humans to validate direct translational relevance. Instead, the sucrose preference test (SPT) is widely used to quantify hedonic motivation [18, 19], as it is rapid (days), inexpensive, and produces outcomes that generally align with data using other testing procedures. Indeed, CSDS in mice causes reliable decreases in preference for sucrose solutions in the SPT, a putative reflection of stress-induced anhedonia and depressive-like behavior [18, 19].
Here we describe the development of a novel juvenile chronic social defeat stress (jCSDS) procedure that produces profound and persistent changes in behavior during adulthood as the result of a single epoch of stress (10 days of CSDS) that occurs early in life. Whereas most reports on CSDS involve both stress exposure and testing during adulthood [20], this new approach involves stress exposure in young mice and testing during adulthood. Specifically, 4-week-old C57BL/6 male mice are defeated by novel aggressive adult males of a different strain (Swiss Webster [CFW]) for 10 consecutive days. After each defeat session, the mouse pair is housed together in the same cage but separated by a plastic barrier. We report that, as expected, mice defeated as juveniles subsequently show robust social avoidance in tests conducted during adulthood. Surprisingly, however, behavior in the SPT [18, 21] is unaffected. To explore whether other tasks might be more sensitive to changes in motivation, we tested jCSDS-exposed mice in the Probabilistic Reward Task (PRT), a procedure often used in humans to study reward learning deficits associated with depressive illness. In a touchscreen version of the PRT that was directly reverse-translated from that used in humans and requires a modest training period (days to weeks), we found that jCSDS reduces response bias for the more richly rewarded stimulus, without producing alterations in task acquisition rates, accuracy, or reaction time. While this procedure has been used in rats [21], there are currently no reports in the literature of it being validated in mice. We also explored relationships among behavior in the social interaction tests, SPT, and PRT. Our findings are consistent with RDoC frameworks [22] that emphasize heterogeneity of neuropsychiatric disorders via subdomains that may be differentially affected and dependent on the type of ELS experienced, while also highlighting the interpretational advantages of using translationally-relevant procedures and endpoints.
Methods
Subjects
For Experiment 1, 3-week-old male C57BL/6 J (C57) mice were obtained from Jackson Laboratory (Bar Harbor, ME, USA). For Experiment 2, 3-week-old male C57BL/6 J (C57) mice were either obtained from Jackson Laboratory (Bar Harbor, ME, USA) or bred in house. The mice were housed under a 12-h light and 12-h dark schedule temperature (21 ± 2 C and humidity (50 ± 20%), and food and water were available ad libitum until PRT training. Following arrival at McLean Hospital, the mice were given 7 days to habituate to the vivarium prior to starting the experiment. All experimental procedures were approved by the Institutional Animal Care and Use Committee at McLean Hospital and were performed in accordance with the National Institutes of Health’s (NIH) Guide for the Care and Use of Animals.
Juvenile chronic social defeat stress (jCSDS)
Virgin male CFW mice (8 weeks, Charles River) were housed with ovariectomized female CFW (8 weeks, Charles River) mice for at least 1 week and screened for aggression with C57 male mice prior to defeat sessions. Only male CFWs that show an attack latency of less than 30 s for 2 consecutive days are used for defeats. Before the start of the first defeat, female CFW mice were permanently removed from the cage.
Male C57 mice (P29 ± 3 days) were exposed to a traditional 10-day defeat paradigm [20, 23], with slight modifications. Specifically, C57 juveniles were placed into the home cage of an aggressive male CFW mouse. Each session consists of physical interactions that proceed until the aggressor has delivered 30 bites. Defeat sessions end after 5 min maximum if fewer than 30 bites occur (average latency of attack: 11.5 +/− 1.7 s; average duration of attack: 87.9 +/− 5.8 s, n = 230 attacks from 23 10-day defeats). The juvenile mice are then housed side-by-side with the male CFW aggressor overnight using a plastic cage divider that physically separates the mice but enables visual and olfactory contact. This procedure was repeated each day with a novel aggressor for 10 consecutive days. At 24 h after the last defeat, each defeated C57 mice was re-housed with another defeated mouse but separated by plastic divider, to control for potential effects of social isolation in adolescence. Control mice were housed with other control mice but separated by a plastic divider from P29 onward, to ensure consistent housing conditions across conditions.
Open field social interaction (OFSI)
Mice were transported from the animal care facility and habituated to a behavioral testing room for 1 h. After habituation, each C57 mouse was placed in a large square arena containing an empty wire cup for 150 s. The C57 was then removed and a non-aggressive CFW male was placed under the wire cup. The C57 was then placed back in the arena for an additional 150 s. Social interaction ratio was calculated as the amount of time spent in the ‘social interaction zone’ (~2 cm circular zone around the wire cup) when the CFW was inside the cup divided by the amount of time spent in the ‘social interaction zone’ when the cup was empty.
Sucrose preference test (SPT)
Preference for a weak sucrose solution (1% wt/vol) to water was measured using a 3-day sucrose preference test via a two-bottle choice paradigm, as described [18]. First, mice were acclimated for 2 days with access to two bottles containing water. On Day 3, mice were given free access to one bottle containing water and the second containing the sucrose solution. Bottles were weighed every 24 h and alternated sides to avoid a side-bias. Sucrose preference scores were calculated as the ratios of sucrose intake to total volume intake.
Probabilistic reward task (PRT)
PRT training
The mouse version of the PRT is a touchscreen task reverse-engineered from human studies [24] and modified from a variant originally designed for rats to objectively quantify reward responsivity [25, 26]. Three days before PRT training commenced, mice were food-restricted via post-session portions of 2–3 grams of rodent chow, followed by training as described previously [26]. Briefly, mice were trained to rear and touch a 5 × 5 cm blue square on a black background and in various positions on the touchscreen to receive 0.02 mL of a highly palatable 20% sweetened condensed milk reward, which was delivered in a well on the opposite wall of the touchscreen. After reliable responding was observed, mice were then trained during 100-trial sessions to discriminate between a long or short white line (24 × 3 cm or 12 × 3 cm) on a black background by responding on one of two virtual levers (5 × 5 cm blue squares) presented below the line, to the left and right of center. Correct responses were rewarded with sweetened condensed milk paired with a tone and brief, bright yellow screen followed by a 10 s blackout period, whereas incorrect responses resulted in a 20 s timeout. During initial PRT training sessions, a correction procedure [21] was employed, where incorrect trials were repeated until a correct response was made prior to advancing to the next trial. After reaching criterion in this phase (10 or fewer errors for both the long and short lines on two consecutive sessions), mice were tested without correction under otherwise identical contingencies. Once mice reached >80% accuracy during two consecutive sessions without correction, PRT testing commenced.
PRT testing
Following line-length discrimination training to criteria, subjects were exposed to a 5-session PRT testing protocol using 3:1 probabilistic reinforcement contingencies such that a correct response to one of the line lengths (long or short) was reinforced 60% of the time (rich stimulus), whereas a correct response to the other line length was reinforced 20% of the time (lean stimulus). Incorrect responses were never reinforced. The line length associated with the rich and lean contingency was determined for each subject during their final two line length discrimination training sessions by examining their accuracies and designating the line length with a higher mean accuracy as the stimulus to be rewarded on the lean schedule. This approach was expressly designed to examine the effects of jCSDS on response bias generated by responsivity to asymmetrical probabilistic contingencies, rather than the amplification of a preexisting inherent bias that is a function of uncontrolled variables.
Statistical analyses
PRT
The PRT yields two primary dependent measures: response bias (which reflects reward function) and discriminability (which reflects baseline response capabilities). These values can be quantified using equations derived from signal detection theory [25] by examining the number of Correct and Incorrect responses for Rich and Lean trial types. Response Bias is calculated using the following log b equation:
High bias values are produced by high numbers of correct responses for rich trials and incorrect responses for lean trials. Discriminability is calculated using the following log d equation:
High discriminability values are produced by high numbers of correct responses for both rich and lean trials. (0.5 is added to all parameters in both equations to avoid instances where no errors are made on a given trial type, thus making log transforms impossible.) In addition, reaction time (latency from line presentation to response) was calculated and presented as individual subject values and session-wide group means (±SEM) for rich and lean trials.
OFSI, SPT, and PRT
If the assumptions of homogeneity of variance were determined to be valid by Levene’s test, unpaired t-tests were used to evaluate group differences (control vs. jCSDS) on behavior in each of the test procedures. If the assumptions were violated, Welch’s t-test was used to evaluate group differences (control versus jCSDS) on behavior. A two-way repeated measures ANOVA was used to evaluate differences in main effect between groups for log b and log d collected each day of PRT testing. The criterion for significance was set at p < 0.05. All statistical analyses were conducted using GraphPad Prism 9 Software (San Diego, CA, USA).
Results
Experiment 1
Experiment 1 was designed to characterize the effects of the novel jCSDS procedure on social interaction (using the OFSI test) and reward function (using the SPT) (Fig. 1A). Based on published work describing the effects of adult CSDS [18, 19], we hypothesized that we would observe stress-induced reductions in both metrics following jCSDS.

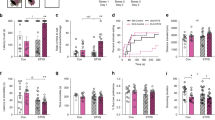
A Schematic of jCSDS and behavioral experiments. B Example heatmaps from open field social interaction test (OFSI). C Social interaction ratio in control (black) versus defeated (purple) adults. D Time spent interacting with non-aggressive CFW. E Total distance traveled in centimeters over OFSI testing.
Adults defeated as juveniles show reduced social interaction
Juvenile male C57BL/6 J mice were subjected to the 10-day jCSDS regimen. Each defeated mouse was then housed in the same cage as another defeated mouse, separated with a plastic barrier that allows for olfactory and visual contact, for the duration of the experiment. Control mice were housed beside other control mice in the same manner to control for housing conditions. Once the mice reached adulthood (P60-70), we examined social approach to a non-familiar non-aggressive male CFW using in the OFSI (Fig. 1B). While control mice showed increased preference for social interaction (i.e., social interaction ratio >1 indicates more time in interaction zone when another mouse is present than when absent), mice exposed to jCSDS avoided social interaction, as evidenced by a low social interaction ratio (unpaired two-tailed t-test, n = 14 controls, 13 defeated, t(25) = 3.350, p = 0.003) (Fig. 1C) as well as a significantly reduced raw social interaction time compared to controls (unpaired two-tailed Welch’s t-test, n = 14 controls, 13 defeated, t(20.64) = 3.161, p = 0.005) (Fig. 1D). There were no differences in distance traveled (unpaired two-tailed t-test, n = 14 controls, 13 defeated, t(25) = 0.696, p = 0.493) (Fig. 1E), suggesting that the change in social interaction was not simply a result of changes in locomotor activity. Control and defeat animals demonstrated minimal aggressive behavior towards the stimulus mouse in OFSI, with no biting or lunging towards the cup observed and only 1 tail rattle observed in a control mouse out of 27 hand-scored interactions. This suggests that our findings are due to a lack of social interaction rather than increases in aggression. These findings suggest that defeated mice retain a long-lasting memory of juvenile trauma into adulthood, creating strong signs of social avoidance behavior.
Adults defeated as juveniles show no difference in sucrose preference
As depressive signs during adulthood are often comorbid with early life trauma in humans, we next wanted to examine depressive-like behaviors in adult mice defeated as juveniles to determine if our model of early life trauma could recapitulate the signs of anhedonia. Following the OFSI tests, we tested the mice in the SPT, which has been used extensively in studies of adult CSDS [27]. Surprisingly, defeated mice showed no difference in sucrose preference for a 1% sucrose solution (Fig. 2A) (unpaired two-tailed t-test, n = 14 control, 13 defeated, t(25) = 0.082, p = 0.940). Interestingly, defeated mice showed an increase in total volume of liquid consumed (t(25) = 2.883, p = 0.008) (Fig. 2B) and amount of sucrose consumed per bodyweight (unpaired two-tailed Welch’s t-test, n = 14 control, 13 defeated, t(15.23) = 2.235, p = 0.041) (Fig. 2C). These findings suggest that juvenile exposure to stress does not produce anhedonia—a finding inconsistent with a vast human literature documenting the persistent effects of ELS across the lifespan—or alternatively, that the SPT is not sensitive to the specific types of reward deficits that are produced by jCSDS.

Average percent sucrose preference across 3 day testing period (A). Average total volume of sucrose solution and water consumed per day calculated across 3 day testing period (B). Average sucrose consumed per body weight per day calculated across 3 day testing period (C).
Experiment 2
Experiment 2 was designed to determine if jCSDS effects would be detectable in the PRT, a task frequently used to study reward function in humans with mood and anxiety disorders [24]. For consistency with Experiment 1, new cohorts of mice were first tested in the OFSI, followed by the PRT instead of the SPT (Fig. 3A). After the PRT, the mice were tested in the SPT, to explore the reproducibility of the findings (lack of effect) from Experiment 1. Based on published work describing the performance of humans with depressive illness in the PRT [23, 24], we hypothesized that this task may identify anhedonia-like signs in mice exposed to jCSDS.

A Schematic of jCSDS and behavioral experiments. B Social interaction ratio in control (black) versus defeated (purple) adults. C Time spent interacting with non-aggressive CFW. D Total distance traveled in centimeters over OFSI testing.
Replication: adults defeated as juveniles show reduced social interaction
As was the case in Experiment 1, while control mice showed increased preference for social interaction, mice exposed to jCSDS avoided social interaction, as indicated by a low social interaction ratio (unpaired two-tailed Welch’s t-test, n = 8 controls, 8 defeated, t(7.86) = 3.273, p = 0.012, 1 defeated mouse was excluded from social interaction analysis as an outlier (Grubb’s outlier test, alpha = 0.05) (Fig. 3B) as well as a significantly reduced raw social interaction time compared to controls (unpaired two-tailed t-test, n = 8 controls, 8 defeated, t(14) = 2.889, p = 0.012) (Fig. 3C). There were no changes in distance traveled (t(14) = 0.375, p = 0.713) (Fig. 3D), suggesting that the change in social interaction was not simply a result of changes in locomotor activity. These findings replicate the OFSI findings from Experiment 1 and suggest that the effect is rigorous and reproducible across independent cohorts of mice exposed to jCSDS.
Adults defeated as juveniles show blunted response bias in the PRT
PRT procedures were initiated at P90-120. Defeated mice showed no difference in learning rates in any of the training stages of PRT compared to control mice (Supplementary Fig. 1B–D). After reaching line-length discrimination training criterion (Fig. 4A), mice were tested on the PRT for 5 daily sessions. Compared to controls, mice exposed to jCSDS showed profoundly blunted response biases towards more richly rewarded stimulus (log b; unpaired two-tailed t-test, n = 8 controls, 9 defeated, t(15) = 2.978, p = 0.009) (Fig. 4B) and, critically, this could not be accounted for by differences in levels of task discriminability (log d; t(15) = 1.382, p = 0.187) (Fig. 4C). This blunted response bias could also not be accounted for by differences in reaction time across rich and lean trial types (main effect of defeat status: F(1,30) = 0.689, p = 0.413) (Fig. 4E), body weight (t(15) = 0.051, p = 0.960) (Supplementary Fig. 1A), or total number of training sessions (t(15) = 0.016, p = 0.988) (Supplementaty Fig. 1E). Rather, although there was only a statistical trend detected for accuracy by rich and lean trial type (main effect of defeat status: F(1,30) = 0.059, p = 0.810), as expected, control subjects had on average higher accuracy during rich trial types and lower accuracy during lean trial types relative to the jCSDS group (Fig. 4D) which, in addition to the log b metric, is another way to characterize their increased response bias for the rich stimulus. Whereas both defeated and control mice showed steady increases in response bias over the course of the 5-day PRT regimen, it was consistently lower in the jCSDS-exposed mice (2-way repeated measures ANOVA, the main effect of defeat status: F(1,15) = 9.050, p = 0.009) (Fig. 5A). In contrast, whereas both defeated and control mice showed comparable increases in discriminability over the same period, there were no group differences at any time point (main effect of defeat status: F(1,15) = 1.939, p = 0.184) (Fig. 5B). These findings suggest that jCSDS in mice causes anhedonic-like phenotypes that persist into adulthood, and that the PRT is more sensitive to this outcome than the SPT.

A Schematic of Probabilistic Reward Task (PRT). B Averaged response bias (log b) over 5 days of PRT testing. C Averaged discriminability (log d) in control (black, n = 8) and adults defeated as juveniles (jCSDS, purple, n = 9) over 5 days of PRT testing. Average accuracy (D) and reaction time (E) over 5 days of PRT testing for Rich and Lean trials.

Response bias (log b, A) and discriminability (log d, B) and on each of the 5 days of PRT testing.
Replication: adults defeated as juveniles show no differences in sucrose preference
After PRT testing, mice were taken off food restriction and given food and water ad libitum for 1 to 2 weeks before the SPT (P150-180). Despite the different order of testing and an intervening period of food restriction, as was the case in Experiment 1, defeated mice showed no difference in sucrose preference for a 1% sucrose solution (Supplementary Fig. 2A) (unpaired two-tailed t-test, n = 8 control, 9 defeated, t(15) = 0.636, p = 0.534). Defeated mice also showed no changes in total volume of liquid consumed (t(15) = 0.735, p = 0.474) (Supplementary Fig. 2B) or amount of sucrose consumed per bodyweight (t(15) = 0.358, p = 0.726) (Supplementary Fig. 2C). These findings replicate the SPT findings from Experiment 1, albeit under slightly different order of testing procedures, and suggest that the lack of effect in this behavioral paradigm is rigorous and reproducible across independent cohorts of mice exposed to jCSDS.
Associations among behaviors in the OFSI, SPT, and PRT
Using the data from mice tested in Experiment 2, we next examined if the behavioral endpoints in any of the tests correlated with one another. Interestingly, percentage sucrose preference was unrelated to either social interaction ratio (Pearson correlation coefficient, R = 0.108, p = 0.680) (Supplementary Fig. 3A) or response bias (Pearson correlation coefficient, R = −0.067, p = 0.805) (Supplementary Fig. 3B). Social interaction ratio and response bias showed a nominal, though not statistically significant, correlation (Pearson correlation coefficient, R = 0.388, p = 0.138) (Supplementary Fig. 3C). These data are consistent with the conclusion that these endpoints reflect separate behavioral domains that are not necessarily coupled to one another.
Discussion
Here we demonstrate that jCSDS, a novel variant of prototypical CSDS procedures in which juvenile (instead of adult) mice are exposed to stress and tested during adulthood, produces robust and persistent changes in behavior. We focused on social interaction and reward function, which are two RDoC-related domains (social processes, positive valence) [22] that are often dysregulated in mood and anxiety disorders [28]. First, jCSDS causes robust decreases in social behavior (i.e., social avoidance) that are unrelated to changes in locomotor activity. Additionally, jCSDS produces a robust blunted response bias to richly rewarded stimulus in PRT, an outcome reflecting anhedonia in humans [23] and rodents [29]. Surprisingly, however, jCSDS failed to alter sucrose preference, an assay commonly used to quantify hedonic behavior in rodents. The observation that jCSDS causes social avoidance but no change in sucrose preference was replicated in two independent cohorts of mice, indicating that it is rigorous and reproducible. Importantly, the SPT was conducted at two different times in the cohorts—immediately after OFSI tests in the first cohort and after OFSI and PRT testing in the second cohort—making it seem unlikely that the failure to observe anhedonia is related to not allowing sufficient time for the phenotype to develop. This pattern of results suggests that the PRT is more sensitive than the SPT to stress-induced changes in reward function, or alternatively, that it may reveal additional facets of anhedonia that are not captured by the SPT alone. Regardless, this work in mice is consistent with a large literature indicating that early life physical and emotional trauma in humans can cause profound changes in the behavior and brain biology that persist across the lifespan [2, 3] and thus establishes a novel methodology that may be useful for modeling ELS-related psychiatric illness.
It is important to note that anhedonia is a construct characterized by substantial heterogeneity and subdomains, and it is unlikely that either the SPT or the PRT captures its full spectrum. Indeed, these tests are expressly designed to evaluate different aspects of reward responsiveness (reward consumption versus reward learning, respectively). The lack of concordance or correlation between assay outcomes, which has been observed previously [29], is both not surprising and consistent with an RDoC-like framework that requires a diversity of tasks to investigate these multidimensional psychological systems. While it is conceivable that subtle changes in behavior in the SPT may have been detectable if a full range of sucrose concentrations had been examined—analogous to dose-finding in drug self-administration studies—a null effect is more easily interpreted than reductions in intake (which could reflect either leftwards or rightward shifts in functions represented by an inverted U-shaped curve). The lack of correlations among the behavioral endpoints in the OFSI, SPT, and PRT suggest that social processes and positive valence can be uncoupled in mice, as is the case in humans, where certain signs are frequently co-morbid but there is considerable heterogeneity among patients and myriad combinations among conditions [28].
The approaches used here may help to promote improvements in the face validity and translational relevance of preclinical studies of human conditions the enhance risk of developing mental health disorders. While ELS can take many forms in humans, jCSDS may most closely approximate bullying, considering that the subordinate mice are younger, smaller, and not genetically related to the aggressors. The mouse version of the PRT was reverse-translated from a version used in humans and involves a digital element (responses delivered via touchscreen), which together enhance the translational relevance of the work. In humans, significant reductions in the tendency to develop response biases for more richly-rewarded stimuli are frequently seen with depressive illness and considered a hallmark sign of anhedonia [24]. The fact that the PRT has been so thoroughly validated in humans and the mouse version shares conceptual, procedural, and analytical elements highlights the interpretational advantages of using translationally-relevant procedures and endpoints. Widespread adoption of these principles may enable the development of improved animal models that more accurately predict outcomes in humans [30].
One limitation of the current experiments is that only male mice were studied. It is well-established that biological sex affects resiliency and susceptibility to stress. Indeed, the diagnoses of mood disorders in which anhedonia is prominent are more prevalent in women [31]. While our recent work with adult CSDS demonstrates that, with at least some stress biomarkers, effects in male mice accurately predicts outcomes in both traumatized men and women [32], it is known that ELS in rodents can produce sex-dependent effects on reward [33]. Application of jCSDS to female mice is hypothetically feasible and currently under development in our labs, with efforts to develop a single standardized protocol that can be used for both sexes. Upon validation, future experiments will investigate sex differences in response to early life physical trauma.
Additionally, while the current study extensively characterizes behavioral phenotypes resulting from early life trauma, the neural mechanisms underlying the anhedonic phenotypes produced by jCSDS are yet to be explored. Previous work from adult social defeat studies highlights a role for the dopaminergic pathway from VTA to nucleus accumbens in socially avoidant behavior in defeated mice [34, 35], with BDNF release playing a critical factor [19, 36]. The prefrontal cortex and its projection to the nucleus accumbens is also implicated in both socially avoidant and decreased sucrose prefence phenotypes produced by adult CSDS [37, 38]. As our defeat targets a window in development in which the prefrontal cortex is not yet fully developed, it will be critical to understand how jCSDS may affect prefrontal development and subsequent innervation of downstream targets.
In summary, we have demonstrated that the jCSDS model of ELS can recapitulate key symptoms of childhood trauma and comorbid psychiatric conditions. In male mice tested during adulthood, jCSDS produces a distinct socially avoidant and anhedonic phenotype. These findings provide the behavioral basis for investigations into the neural circuit alterations as well as more holistic (e.g., peripheral) biomarkers of early life trauma and abuse that may enable improvements in the diagnosis, treatment, and prevention of psychiatric illness.
References
Lange I, Goossens L, Bakker J, Michielse S, van Winkel R, Lissek S, et al. Neurobehavioural mechanisms of threat generalization moderate the link between childhood maltreatment and psychopathology in emerging adulthood. J Psychiatry Neurosci. 2019;44:185–94.
Colich NL, Rosen ML, Williams ES, McLaughlin KA. Biological aging in childhood and adolescence following experiences of threat and deprivation: a systematic review and meta-analysis. Psychol Bull. 2020;146:721–64.
McLaughlin KA, Weissman D, Bitrán D. Childhood adversity and neural development: a systematic review. Annu Rev Dev Psychol. 2019;1:277–312.
McLaughlin KA, Sheridan MA, Lambert HK. Childhood adversity and neural development: deprivation and threat as distinct dimensions of early experience. Neurosci Biobehav Rev. 2014;47:578–91.
Keding TJ, Heyn SA, Russell JD, Zhu X, Cisler J, McLaughlin KA, et al. Differential patterns of delayed emotion circuit maturation in abused girls with and without internalizing psychopathology. Am J Psychiatry. 2021;178:1026–36.
Waters RC, Gould E. Early life adversity and neuropsychiatric disease: differential outcomes and translational relevance of rodent models. Front Syst Neurosci. 2022;16:860847.
Tractenberg SG, Levandowski ML, de Azeredo LA, Orso R, Roithmann LG, Hoffmann ES, et al. An overview of maternal separation effects on behavioural outcomes in mice: evidence from a four-stage methodological systematic review. Neurosci Biobehav Rev. 2016;68:489–503.
Gallo M, Shleifer DG, Godoy LD, Ofray D, Olaniyan A, Campbell T, et al. Limited bedding and nesting induces maternal behavior resembling both hypervigilance and abuse. Front Behav Neurosci. 2019;13:167.
Naeem N, Zanca RM, Weinstein S, Urquieta A, Sosa A, Yu B, et al. The neurobiology of infant attachment-trauma and disruption of parent–infant interactions. Front Behav Neurosci. 2022;16:882464.
Hasegawa S, Miyake Y, Yoshimi A, Mouri A, Hida H, Yamada K, et al. Dysfunction of serotonergic and dopaminergic neuronal systems in the antidepressant-resistant impairment of social behaviors induced by social defeat stress exposure as juveniles. Int J Neuropsychopharmacol. 2018;21:837–46.
Mouri A, Ukai M, Uchida M, Hasegawa S, Taniguchi M, Ito T, et al. Juvenile social defeat stress exposure persistently impairs social behaviors and neurogenesis. Neuropharmacology. 2018;133:23–37.
Coppens CM, Siripornmongcolchai T, Wibrand K, Alme MN, Buwalda B, de Boer SF, et al. Social defeat during adolescence and adulthood differentially induce BDNF-regulated immediate early genes. Front Behav Neurosci. 2011;5:72.
McCullough KM, Missig G, Robble MA, Foilb AR, Wells AM, Hartmann J, et al. Nucleus accumbens medium spiny neuron subtypes differentially regulate stress-associated alterations in sleep architecture. Biol Psychiatry. 2021;89:1138–49.
Wells AM, Ridener E, Bourbonais CA, Kim W, Pantazopoulos H, Carroll FI, et al. Effects of chronic social defeat stress on sleep and circadian rhythms are mitigated by kappa-opioid receptor antagonism. J Neurosci. 2017;37:7656–68.
Donahue RJ, Landino SM, Golden SA, Carroll FI, Russo SJ, Carlezon WA Jr. Effects of acute and chronic social defeat stress are differentially mediated by the dynorphin/kappa-opioid receptor system. Behav. Pharmacol. 2015;26:654–63.
Donahue RJ, Muschamp JW, Russo SJ, Nestler EJ, Carlezon WA. Effects of striatal ΔFosB overexpression and ketamine on social defeat stress–induced anhedonia in mice. Biol Psychiatry. 2014;76:550–8.
Carlezon WA, Chartoff EH. Intracranial self-stimulation (ICSS) in rodents to study the neurobiology of motivation. Nat Protoc. 2007;2:2987–95.
Krishnan V, Han MH, Graham DL, Berton O, Renthal W, Russo SJ, et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell. 2007;131:391–404.
Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science (1979). 2006;311:864–8.
Golden SA, Covington HE, Berton O, Russo SJ. A standardized protocol for repeated social defeat stress in mice. Nat Protoc. 2011;6:1183–91.
Kangas BD, Branch MN. Empirical validation of a procedure to correct position and stimulus biases in matching‐to‐sample. J Exp Anal Behav. 2008;90:103–12.
Insel T, Cuthbert B, Garvey M, Heinssen R, Pine DS, Quinn K, et al. Research domain criteria (RDoC): toward a new classification framework for research on mental disorders. Am J Psychiatry. 2010;167:748–51.
Kudryavtseva NN, Bakshtanovskaya IV, Koryakina LA. Social model of depression in mice of C57BL/6J strain. Pharm Biochem Behav. 1991;38:315–20.
Pizzagalli DA, Jahn AL, O’Shea JP. Toward an objective characterization of an anhedonic phenotype: a signal-detection approach. Biol Psychiatry. 2005;57:319–27.
Luc OT, Pizzagalli DA, Kangas BD. Toward a quantification of anhedonia: unified matching law and signal detection for clinical assessment and drug development. Perspect Behav Sci. 2021;44:517–40.
Kangas BD, Wooldridge LM, Luc OT, Bergman J, Pizzagalli DA. Empirical validation of a touchscreen probabilistic reward task in rats. Transl Psychiatry. 2020;10:285.
Heshmati M, Russo SJ. Anhedonia and the brain reward circuitry in depression. Curr Behav Neurosci Rep. 2015;2:146–53.
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. (American Psychiatric Association Publishing, 2022). https://doi.org/10.1176/appi.books.9780890425787.
Kangas BD, Short AK, Luc OT, Stern HS, Baram TZ, Pizzagalli DA. A cross-species assay demonstrates that reward responsiveness is enduringly impacted by adverse, unpredictable early-life experiences. Neuropsychopharmacology. 2022;47:767–75.
Bale TL, Epperson CN. Sex as a biological variable: who, what, when, why, and how. Neuropsychopharmacology. 2017;42:386–96.
Hyde JS, Mezulis AH. Gender differences in depression: biological, affective, cognitive, and sociocultural factors. Harv Rev Psychiatry. 2020;28:4–13.
McCullough KM, Katrinli S, Hartmann J, Lori A, Klengel C, Missig G, et al. Blood levels of T-Cell Receptor Excision Circles (TRECs) provide an index of exposure to traumatic stress in mice and humans. Transl Psychiatry. 2022;12:423.
Levis SC, Bentzley BS, Molet J, Bolton JL, Perrone CR, Baram TZ, et al. On the early life origins of vulnerability to opioid addiction. Mol Psychiatry. 2021;26:4409–16.
Chaudhury D, Walsh JJ, Friedman AK, Juarez B, Ku SM, Koo JW, et al. Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nature. 2013;493:532–6.
Wook Koo J, Labonté B, Engmann O, Calipari ES, Juarez B, Lorsch Z, et al. Essential role of mesolimbic brain-derived neurotrophic factor in chronic social stress–induced depressive behaviors. Biol Psychiatry. 2016;80:469–78.
Walsh JJ, Friedman AK, Sun H, Heller EA, Ku SM, Juarez B, et al. Stress and CRF gate neural activation of BDNF in the mesolimbic reward pathway. Nat Neurosci. 2014;17:27–9.
Covington HE, Lobo MK, Maze I, Vialou V, Hyman JM, Zaman S, et al. Antidepressant effect of optogenetic stimulation of the medial prefrontal cortex. J Neurosci. 2010;30:16082–90.
Vialou V, Bagot RC, Cahill ME, Ferguson D, Robison AJ, Dietz DM, et al. Prefrontal cortical circuit for depression- and anxiety-related behaviors mediated by cholecystokinin: role of ΔFosB. J Neurosci. 2014;34:3878–87.
Acknowledgements
We thank the Tommy Fuss Foundation for providing support that enabled us to initiate the line of work described in this article, and Dr. Dominika Burek for assistance with the probabilistic reward task experiments.
Funding
This work was supported primarily by a grant from the Tommy Fuss Foundation (to WAC). Additional resources were provided by P50MH115874 (WAC & KJR), R01MH063266 (to WAC), and R01DA047575 (to BDK). Part of EH’s time spent preparing this article for publication was supported by training grant T32MH125786 (to WAC & KJR).
Author information
Authors and Affiliations
Contributions
EH, ELN, KJR, BDK, and WAC conceived and designed the experiments. EH and EF conducted the experiments, and EH and BDK analyzed the data. EH, EF, BDK, and WAC wrote sections of the manuscript and all authors provided feedback on the version submitted for publication.
Corresponding author
Ethics declarations
Competing interests
WAC and KJR are members of the NPP editorial board. WAC is a consultant for Psy Therapeutics and has sponsored research agreements with Cerevel Therapeutics and Delix Therapeutics. BDK has sponsored research agreements with Compass Pathways, Delix Therapeutics, Engrail Therapeutics, Neurocrine Biosciences, Psy Therapeutics, and Takeda. KJR has performed scientific consultation for Bioxcel, Bionomics, Acer, Takeda, and Jazz Pharma; serves on Scientific Advisory Boards for Sage and the Brain Research Foundation, and he has received sponsored research support from Takeda, Brainsway, and Alto Neuroscience. EH, EF, and ELN report no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hisey, E.E., Fritsch, E.L., Newman, E.L. et al. Early life stress in male mice blunts responsiveness in a translationally-relevant reward task. Neuropsychopharmacol. 48, 1752–1759 (2023). https://doi.org/10.1038/s41386-023-01610-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-023-01610-7
