
Abstract
Social reinforcement-based treatments are effective for many, but not all, people with addictions to drugs. We recently developed an operant rat model that mimics features of one such treatment, the community-reinforcement approach. In this model, rats uniformly choose social interaction over methamphetamine or heroin. Abstinence induced by social preference protects against the incubation of drug-seeking that would emerge during forced abstinence. Here, we determined whether these findings generalize to cocaine and whether delaying or increasing effort for social interaction could reveal possibly human-relevant individual differences in responsiveness. We trained male and female rats for social self-administration (6 days) and then for cocaine self-administration, initially for 2-h/day for 4 days, and then for 12-h/day continuously or intermittently for 8 days. We assessed relapse to cocaine seeking after 1 and 15 days. Between tests, the rats underwent either forced abstinence or social-choice-induced abstinence. After establishing stable social preference, we manipulated the delay for both rewards or for social reward alone, or the response requirements (effort) for social reward. Independent of cocaine-access conditions and sex, operant social interaction inhibited cocaine self-administration and prevented incubation of cocaine seeking. Preference for social access was decreased by the delay of both rewards or social reward alone, or by increased response requirements for social reward, with notable individual variability. This choice procedure can identify mechanisms of individual differences in an animal model of cocaine use and could thereby help screen medications for people who are relatively unresponsive to treatments based on rewarding social interaction.
Similar content being viewed by others

Introduction
For people who use potentially addictive drugs, one buffer against transitioning to addiction seems to be embeddedness in social structures, both small (e.g., family) and large (e.g., professional or political communities) [1,2,3,4,5]. For people who have already transitioned to addiction, the effects of social connection can be harnessed in behavioral treatments, where social reinforcers (e.g., participation in support groups and job training) are systematically made more accessible and reliable, contingent on their being chosen over drug use [6,7,8,9]. These socially based treatments, which include the community-reinforcement approach and the therapeutic workplace, are effective in some people, a finding of both heuristic importance (because it shows that addiction does not inevitably obliterate the capacity to choose non-drug rewards) and practical importance (because it offers a means to treat types of addiction for which no effective medications are known). However, social-based behavioral treatments are not effective for every patient, at least not at any practically achievable magnitude or schedule of nondrug reinforcement [10,11,12,13,14]. This might partly reflect the fact social interactions can have negative consequences and promote drug use and relapse [3, 15, 16], but it also suggests that there are considerable individual differences in responsiveness to social rewards, and it points to a need to discover new treatments for people who are less responsive. One step toward treatment discovery is the establishment of an animal model of the condition to be treated.
We recently introduced a rat model designed to mimic a narrower range of positive social interactions that inhibit drug-taking and relapse in operant-based therapeutic situations [17]. In this model, rats can choose between the self-administered drug and interaction with a social peer [17, 18]. When we used this model in rats that had already met criteria for drug addiction in other established models (the escalation model [19], the intermittent-access binge model [20], or a DSM-IV-based model [21, 22]), we found that immediate access to a conspecific prevented their resumption of methamphetamine or heroin self-administration [17, 18, 23]. We also found that when we induced drug abstinence with our socially based procedure [24, 25], we reduced subsequent incubation of heroin craving [23] and prevented subsequent incubation of methamphetamine craving [17]. Incubation of drug craving refers to the time-dependent increase in drug seeking during abstinence [26]. The most salient gap between our results and the clinical situation we were modeling was that rats, unlike humans, chose social reinforcement over drug reinforcement on almost all occasions (unless we significantly delayed or punished the operant response for the social reinforcer). We discussed possible reasons for this, such as differences in the concreteness or abstractness with which humans (versus rats) evaluate the reward value of interacting with peers [17].
In the present study, we first determined whether our prior results with heroin and methamphetamine would generalize to cocaine, under conditions that promote either (1) very high and escalated continuous drug intake for 12 h per day or (2) multiple binge episodes (24 five-min episodes per day over 12 h). For this purpose, we used the escalation and intermittent-access drug self-administration addiction models [19, 20, 27, 28]. Next, we used a discrete trial drug-versus-nondrug choice procedure [17, 29,30,31] to determine whether parametric manipulations of delay and response requirements (effort) could uncover individual differences in social interaction versus cocaine choice.
Our results indicate that even after daily extended access to cocaine that was twice as long as the daily access to methamphetamine and heroin in our previous studies (6-h/day), operant social reward prevented cocaine self-administration and incubation of craving (in both male and female rats). In addition, parametric manipulations sufficed to uncover the rats’ differing propensities to resume cocaine seeking when the social reward was delayed or more effortful. These individual differences might identify a subset of rats comparable to the subset of people who do not respond well to socially based treatments. Such rats might be a suitably stringent testing ground for new pharmacological/biomedical treatments.
Materials and methods
For information on Materials and methods, see the Supplemental Online Materials. Our procedures followed the guidelines outlined in the Guide for the Care and Use of Laboratory Animals (8th edition; http://grants.nih.gov/grants/olaw/Guide-for-the-Care-and-Use-of-Laboratory-Animals.pdf). The study has been approved by the NIDA IRP Animal Care and Use Committee.
Exp. 1: effect of social reward on cocaine self-administration and incubation of cocaine craving
We tested whether social choice-induced abstinence would protect against incubation of cocaine craving after a history of continuous (Exp. 1A) or intermittent (Exp. 1B) access to cocaine self-administration. We used 2 cohorts (Continuous, Intermittent) with two groups of rats each (11–12 males;11 females) in an experimental design that included the between-subjects factors of Abstinence condition (Forced, Voluntary) and Sex (Male, Female).
Training
We first trained rats to self-administer palatable food (3 sessions, 2-h/session, 1 pellet). Next, we trained them to self-administer social interaction (6 sessions, 60-trials/session, 60-s interaction). Next, we trained them to self-administer cocaine (4 sessions, 2-h/session; 8 sessions, 12-h/session; 0.75 mg/kg/infusion).
Discrete choice tests
We determined social interaction versus cocaine choice for 2 sessions during the training phase and then for 10 sessions (over 14 days) during the voluntary abstinence phase.
Forced abstinence
After training, we returned the rats to their home cage for 14 days and handled them twice/week.
Relapse test
We tested the forced and voluntary abstinence rats for cocaine seeking under extinction conditions on abstinence days 1 and 15. The duration of the test session was 30 min on day 1 to minimize the carryover effect of extinction learning, which may subsequently decrease drug seeking on day 15 (180-min session) testing.
Exp. 2: effect of diminished value of social interaction on social preference
The goal of Exp. 2 was to determine whether individual differences in the choice of social interaction versus cocaine would emerge when we parametrically manipulate the delay and response requirement (effort) to obtain the social reward. Based on results from a recent study [32], we also delayed access to both cocaine and social interaction. We used a male (n = 6) and female (n = 5–6) rats from the voluntary abstinence group of Exp. 1B. After the day 15 relapse test, we assessed baseline choice preference over 3 sessions.
Discrete choice sessions
We determined the rats’ social interaction versus cocaine choice using two delay discounting manipulations (15 choice tests, Exp. 2A—delays for both rewards; 14 choice tests, Exp. 2B—delays for social reward only), and a response requirement (effort) manipulation of increasing the fixed-ratio requirement (10 choice tests, Exp. 2C).
Statistical analysis
We used factorial ANOVA and t-tests using SPSS (IBM, version 27, GLM procedure). When we obtained significant main effects and interaction effects (p < 0.05, two-tailed), we followed them with post-hoc tests (Fisher PLSD). Because our factorial ANOVA yielded multiple main and interaction effects, we only report significant effects that are critical for data interpretation. We indicate the results of the post-hoc analyses in the figures but do not describe them in the “Results”. We indicate p < 0.001 and provide exact p values for results smaller than 0.05 and greater than 0.001. Supplementary Table S1 provides a complete report of the statistical results for the data shown in the figures.
For the choice data, our outcome measure was preference score (number of social rewards/[number of social rewards + number of drug infusions]). We provide our rationale for this outcome measure in the Supplemental Online Section. We also provide, in Table S2, an alternative analysis in which Reward Type (social interaction, cocaine) is treated as a predictor of the number of rewards; the conclusions are the same.
To characterize individual differences across rats, we performed cluster analysis of the choice data shown in Fig. 3 using R [33] with the packages NbClust [34] and kml [35]. Because there are many procedures for identifying the most relevant number of clusters in a data set, but no single procedure is recognized as being superior, the approach taken by NbClust is to obtain a consensus by applying multiple procedures for optimizing the number of clusters.
No statistical methods were used to predetermine sample sizes, but our sample sizes are similar to those reported in previous publications [17, 18, 23].
Results
Effect of social reward on cocaine self-administration and incubation of craving
The goal of Exp. 1 was to determine whether our prior findings with methamphetamine—an inhibitory effect of operant social interaction on extended access (6-h/day) continuous or intermittent methamphetamine self-administration and incubation of methamphetamine craving after social-choice-induced voluntary abstinence [17, 18]—would generalize to cocaine under access conditions (12-h/day) that induce escalated cocaine intake. The experiment (Fig. 1A) consisted of the following: (1) self-administration training, (2) relapse tests 1 day after the last self-administration session, (3) social choice-induced voluntary abstinence or homecage forced abstinence, and (4) relapse test after 15 abstinence days.

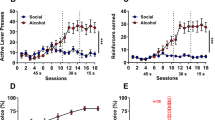
A Timeline of the experiments. B Self-administration training (rewards: social interaction or cocaine infusion). The number of social rewards (60 trials) or cocaine infusions (2 h–4 d; 12 h–12 d) in male (n = 12) and female (n = 11) rats. C Choice during training. Left panel: social rewards and cocaine infusions earned during 2 discrete-choice sessions (15 trials per session). Middle panel: average of preference score (number of social rewards/[number of social rewards + number of drug infusions]). 0 indicates a preference for cocaine; 1 indicates a preference for social reward. Right panel: individual data for male and female rats during the two choice sessions. D Voluntary abstinence. Left panel: Social rewards and cocaine infusions earned during 10 discrete-choice sessions (15 trials per session). Middle panel: average of preference score. 0 indicates a preference for cocaine; 1 indicates a preference for social reward. Right panel: individual data for male and female rats during the ten choice sessions. E Incubation (relapse) test. Active-lever presses during the 30-min test sessions (including individual data) for the forced-abstinence (left panel) and social-choice (right panel) groups. During testing, active-lever presses led to the contingent presentation of the discrete light cue previously paired with cocaine infusions during training, but not cocaine (extinction conditions). *Different from test day 1, p < 0.05. #Different from the social-choice voluntary-abstinence group on test day 15, p < 0.05. Forced-abstinence condition: 6 male/6 female rats; social-choice abstinence condition: 6 male/5 female rats. Data are mean ± SEM. See also Figs. S1, S2, and S3.
Continuous access cocaine self-administration (Exp. 1A)
Training and choice
During training for food self-administration, food-sated male and female rats increased their food intake over time, and no sex differences were observed (Fig. S1A). During social self-administration, the rats increased their operant response to gain access to a social peer over days and no sex differences were observed (Fig. 1B). The statistical analysis showed a significant effect of Session (F5,105 = 26.6, p < 0.001) but not sex or session × sex interaction (p values > 0.05). During training for cocaine self-administration, the rats escalated their cocaine intake over time and no sex differences were observed (Figs. 1B and S2A). The statistical analysis showed a significant effect of the session (F11,231 = 133.0, p < 0.001) but not sex or session × sex interaction (p values > 0.05). During the two choice sessions, the statistical analysis of the preference score (number of social rewards/[number of social rewards + number of drug infusions]) showed no significant effects of the session, sex, or interaction (p values > 0.05). Figure 1C middle and right panels show average and individual preference scores.
Abstinence phase
During the 14-day abstinence phase, the male and female rats in the voluntary-abstinence groups showed a robust preference for social interaction over cocaine, an effect that was stronger in males than in females. The statistical analysis of the preference score showed significant effects of the session (F9,81 = 4.28, p < 0.001) and sex (F1,9 = 5.8, p = 0.04), but no interaction (p > 0.05). Figure 1D middle and right panels show average and individual preference scores.
Relapse tests
In both sexes, active-lever presses during the tests were higher after 15 abstinence days than after 1 day in the forced-abstinence group but not the voluntary-abstinence group (Fig. 1E). These data demonstrate that social choice-induced voluntary abstinence prevented incubation of cocaine craving. The statistical analysis, which included the between-subjects factors of abstinence condition (forced, voluntary) and sex (male, female), and the within-subjects factor of abstinence day (1, 15) and lever (active, inactive), showed a significant interaction of abstinence condition × abstinence day × lever interaction (F1,19 = 6.2, p = 0.02) but no main effect of sex or interactions with sex (p values > 0.05). We also analyzed the data from the 180-min relapse-test session on day 15 using the factors of abstinence condition, sex, and lever (Fig. S3A). This analysis showed a significant interaction between the abstinence condition and Lever (F1,19 = 6.2, p = 0.02), but no main effect of sex or interactions with sex (p values > 0.05).
Intermittent access cocaine self-administration (Exp. 2A)
Training and choice
During training for food self-administration, the rats increased their food intake over time, and no sex differences were observed (Fig. S1B). During social self-administration, the rats increased their lever presses to gain access to a social peer and no sex differences were observed (Fig. 2B). The statistical analysis showed a significant main effect of the session (F5,100 = 24.6, p < 0.001), but no effect of sex or session × sex interaction (p values > 0.05). During training for cocaine self-administration, the rats escalated their cocaine intake over time and no sex differences were observed (Figs. 2B and S2B). The statistical analysis showed a significant effect of Session (F11,220 = 79.7, p < 0.001) but no effect of sex or session × sex interaction (p values > 0.05). In addition, the statistical analysis showed that male and female rats increased their response rate (infusions/min) within the 5 min of drug availability (Fig. S2C). During the two choice sessions, the statistical analysis of the preference score showed no significant effects of the session, sex, or interaction (p values > 0.05). Figure 2C middle and right panels show average and individual preference scores.

A Timeline of the experiments. B Self-administration training (rewards: social interaction or cocaine infusion). The number of social rewards (60 trials) or cocaine infusions (2 h–4 d; 12 h–12 d) in male (n = 11) and female (n = 11) rats. C Choice during training. Left panel: social rewards and cocaine infusions earned during two discrete-choice sessions (15 trials per session). Middle panel: average of preference score (number of social rewards/[number of social rewards + number of drug infusions]). 0 indicates a preference for cocaine; 1 indicates a preference for social reward. Right panel: individual data for male and female rats during the two choice sessions. D Voluntary abstinence. Left panel: Social rewards and cocaine infusions earned during 10 discrete-choice sessions (15 trials per session). Middle panel: average of preference score. 0 indicates a preference for cocaine; 1 indicates a preference for social reward. Right panel: individual data for male and female rats during the ten choice sessions. E Incubation (relapse) test. Active-lever presses during the 30-min test sessions (including individual data) for the forced-abstinence group (left panel) and social-choice group (right panel). During testing, active-lever presses led to the contingent presentation of the discrete light cue previously paired with cocaine infusions during training, but not cocaine (extinction conditions). *Different from test day 1, p < 0.05. #Different from the social-choice voluntary-abstinence group on test day 15, p < 0.05. Forced-abstinence condition: 5 male/5 female rats; social-choice abstinence condition: 6 male/6 female rats. Data are mean ± SEM. See also Figs. S1, S2, and S3.
Abstinence phase
During the 14-day abstinence phase, the rats in the voluntary abstinence groups showed a strong preference for social reward over cocaine. The statistical analysis of the preference score showed a significant effect of Session (F9,90 = 3.3, p = 0.002) but no effect of sex or session × sex interaction (p values > 0.05). Figure 2D middle and right panels show average and individual preference scores.
Relapse tests
In both sexes, active-lever presses during the tests were higher after 15 abstinence days than after 1 day in the forced but not voluntary abstinence group (Fig. 2E). These data demonstrate that social-choice-induced voluntary abstinence prevented incubation of cocaine craving. The statistical analysis, which included the between-subjects factors of abstinence condition and sex, and the within-subjects factor of abstinence day and Lever, showed a significant interaction of abstinence condition × abstinence day × lever (F1,18 = 6.75, p = 0.018) but no main effects of sex or interactions with sex (p values > 0.05). We also analyzed the data from the 180-min relapse-test session on day 15, using the factors of abstinence condition, sex, and lever (Fig. S3B). This analysis showed a significant interaction between the abstinence condition and lever (F1,19 = 6.24, p = 0.02) but no main effect of sex or interactions with sex (p values > 0.05).
Together, the results of Exp. 1 demonstrate that the protective effect of social reward on drug self-administration and incubation of drug craving generalized to cocaine. Additionally, we did not observe sex differences in cocaine self-administration or incubation of cocaine craving. However, the preference for social reward was lower in the female rats than in the male rats after continuous-access (but not intermittent-access) cocaine self-administration.
Effect of diminished value of social interaction on social preference: the emergence of individual differences
The goal of Exp. 2 was to determine whether individual differences in preference for social reward versus cocaine would emerge by increasing the delay to obtain the social reward and cocaine or the social reward alone, or by increasing the response requirements or effort to obtain the social reward.
The discrete choice procedure with the delay of both rewards (Exp. 2A)
During the 15 choice sessions (Fig. 3A), the preference for social interaction decreased while the preference for cocaine increased as delay for both rewards increased (Fig. 3B-I); this effect was sex independent (Fig. 3B-II–V). The statistical analysis of the preference score showed a significant effect of delay (F6,60 = 17.0, p < 0.001) but no effects of sex or interaction (p values > 0.05). When we plotted the individual data (Fig. 3B-II—heatmap: dark red = social preference; light white = cocaine preference), we observed that some rats immediately decreased their social preference after 10–20-s delays while others were more resistant, changing their preference only after 40–50 s delays. To statistically quantify these individual differences, we determined the relevant number of K-means clusters [34, 35] for the choice data (Fig. 3B-III). We found that the most relevant number of clusters (Fig. 3B-II) was six, indicating heterogeneity in social interaction vs. cocaine choice in our sample; some rats showed a sharp decrease in preference for social interaction across delays, while others showed a gradual decrease across delays, while others showed a decrease only at the two longest delays (Fig. 3B-IV, V).

A Timeline of the experiments. B Choice with delay for both rewards (social interaction and cocaine infusion). C Choice with delay for social reward only. D Choice with increased fixed-ratio requirements. For each line: I: mean number of social rewards and cocaine infusions earned during 15 discrete choice sessions (15 trials per session). Male (n = 6) and female (n = 5–6) data are collapsed. Data are shown as the mean ± SEM of two consecutive sessions with the same delay time. Gradations of blue represent decreased preference for social reward (dark blue = strong preference; light blue = no preference); gradations of orange represent increased preference for cocaine reward (dark orange = strong preference; light orange = no preference). II: heatmap of individual social preference scores for male (M) and female (F) rats. Light white indicates a preference for cocaine (score = 0) and dark red indicates a preference for social interaction (score = 1). Colored boxes around the set of rows indicate the K-means clusters. III: Graphical depiction of the choice trajectories of K-mean clusters; each cluster is represented by a specific color that is consistent across the heatmap and trajectory curves. Individual rats are represented by thin lines, and the mean for each cluster is represented by same-color thick lines. IV: average of preference score. 0 indicates a preference for cocaine; 1 indicates a preference for social reward. V: individual data for cocaine infusions earned during the discrete-choice sessions (15 trials per session).
The discrete-choice procedure with the delay of social reward (Exp. 2B)
During the 14 choice sessions (Fig. 3A), the preference for social interaction decreased while the preference for cocaine increased as the delay for social reward increased (Fig. 3C-I); this effect was sex independent (Fig. 3C-II–V). The statistical analysis of the preference score showed a significant effect of delay (F6,54 = 27.7, p < 0.001) but no effect of sex or interaction (p values > 0.05). Although these findings were generally similar to the findings from Exp. 2A, when we plotted the individual data (Fig. 3C-II), we observed that rats were more resistant to changing their preference when we delayed only the social reward relative to when we simultaneously delayed both rewards. We also found individual differences after a delay of social reward only (Fig. 3C-II); the most relevant number of clusters (Fig. 3C-III) was three, with one rat showing no change in preference for social interaction over time, other rats showing a moderate decrease over time, and other rats showing a steeper decrease over time (Fig. 3C-IV, V).
The discrete-choice procedure with increased response requirement for social reward (Exp. 2C)
During the ten choice sessions (Fig. 3A), the preference for social reward decreased while the preference for cocaine increased as the fixed-ratio requirements for social reward increased (Fig. 3D-I); this effect was sex independent (Fig. 3D-II–V). The statistical analysis of the preference score showed a significant effect of fixed-ratio requirement (F4,36 = 18.8, p < 0.001) but no effect of sex or interactions (p values > 0.05). Of note, unlike the delay manipulations, the effort manipulation caused a strong preference shift toward cocaine; most rats preferred cocaine over social interaction when the fixed-ratio requirement was increased to 16 (Fig. 3D-II). We found evidence for individual differences (Fig. 3D-II), with the most relevant number of clusters (Fig. 3C-III) being three; some rats showed only a slight decrease in preference for social interaction, other rats showed a graded decrease as the response requirement increased, and one rat showed a low preference at all FR values higher than one (Fig. 3C-IV, V).
The results of Exp. 2 demonstrate that male and female rats who strongly prefer immediate access to social interaction over immediate cocaine infusions will significantly increase their cocaine choice when the social reward is delayed or requires more effort. Our data also extend a surprising recent observation from Canchy et al. [32]. that delaying access to both cocaine and the alternative nondrug reward increases preference for cocaine. Finally, the choice data indicate that individual differences in propensity toward cocaine seeking (at the expense of social interaction) can emerge when the value of the social reward is diminished by delaying access for social interaction or increasing the effort to obtain it.
Discussion
We showed protective effects of social interaction in rat models of cocaine addiction, along with individual differences in resistance to the loss of that protection. Independent of the model we used (escalation or intermittent access) to establish that the rats were self-administering cocaine in an “addicted” way, we found that their cocaine self-administration significantly decreased (and subsequent incubation of cocaine craving was prevented) when presses on a different lever were immediately reinforced with social interaction. When access to the social peer was delayed or required more effort, the rats showed individual differences in their propensity to choose cocaine over social interaction. These findings did not generally differ by sex.
Protective effect of social reward on cocaine self-administration and incubation
The aspect of our findings that was nearly invariant across rats—that immediate response-contingent operant social reward protected against the continuation of cocaine self-administration—extends results from our previous studies with methamphetamine and heroin, where we found that this protection was independent of sex, drug dose, housing conditions, or the “addiction” score in the animal models we used [17, 23, 25]. In the present study, we trained the rats under conditions that either promoted escalation to very high levels of continuous cocaine intake or under conditions that promoted multiple cocaine-binge episodes (24 five-min episodes/day) over 12 h per day. We chose these procedures to establish that the rats were strongly motivated to take and seek cocaine [19, 20, 27, 28]. Yet even these rats strongly preferred social interaction with a peer, demonstrating the profound effect of immediate and low-effort social reward on drug intake across drug classes and experimental procedures.
Social reward also prevented the incubation of cocaine craving (and this occurred in both male and female rats). Previous studies have shown that incubation of cocaine craving is decreased by the availability of alternative non-drug rewards, mostly in the form of experimenter-imposed enriched environments and aerobic exercise, during periods of forced abstinence [36,37,38]. The forced-abstinence studies did not model the human scenario that was of interest to us, in which lapses to drug use occur after a period of self-imposed abstinence driven by the pending loss of alternative non-drug rewards [24, 25]. Our current findings with incubation of craving for cocaine are consistent with our prior findings with methamphetamine [17, 18], but differ from our prior findings with heroin, for which social-choice-induced voluntary abstinence was only modestly protective against incubation of craving [23]. Any comments on mechanism would be speculative, but one broad implication is that social reward might be more protective against lapses to psychostimulant use than against lapses to opioid use—which would add to evidence that addiction can differ in important ways across drug classes [39,40,41].
Finally, as in our prior studies on voluntary abstinence with heroin and methamphetamine [17, 42, 43], we did not observe sex differences in cocaine self-administration, regardless of access condition, nor did we observe sex differences in the incubation of cocaine craving, regardless of abstinence condition (forced or voluntary). This is in contrast with two studies reporting higher incubation of cocaine craving after forced abstinence in female rats with a history of either intermittent or continuous access [44, 45]. Although comparisons across studies require caution, we suspect that the difference lies in the training conditions. We initially trained all rats, including the forced-abstinence group, for “self-administration” of a social peer. In contrast, Nicolas et al. [44]. and Kerstetter et al. [45]. trained rats for cocaine self-administration only. Based on previous reports showing the protective effect of social reward on drug seeking and craving [36, 38, 46,47,48], the interaction with a social peer during the social self-administration phase might have decreased cocaine craving after forced abstinence in female rats.
In reconciling the differences between our results and those of these previous studies, another important methodological difference is that we did not measure the estrous cycle, which plays a critical role in the incubation of cocaine craving after forced abstinence with higher incubation during estrus vs. non-estrus [43,44,45].
Individual differences in propensity to loss of the protective effect of social interaction
A main finding in our study was that rats varied in their propensity to resume cocaine self-administration when we either delayed the social reward or increased the response requirement or effort to obtain it. These results for the delay manipulation extend our prior findings with methamphetamine [17, 25]. Possibly relevant to the effect of delay is a recent suggestion that rats’ preference for food reward over cocaine [29] is due to a pharmacokinetic lag in cocaine’s rewarding effects in the brain relative to those of food [32]. In support of this hypothesis, the same group of authors showed that rats will choose cocaine over food if both rewards are equally delayed in a delay-discounting procedure [49]. Our results extend these findings to rewarding social interaction. We think it is unlikely, however, that the inter-individual variability we saw was attributable to individual differences in the pharmacokinetics of cocaine. Even if it were, this would not explain the inter-individual variability we saw when we increased the response requirement for social reward, leaving the timing unchanged. However, the interpretation of our choice data and their implications to individual differences in cocaine versus social reward choice should be made with caution because of the relatively low number of rats used in Exp. 2 (6 males and 5–6 females).
Finally, our results extend those from previous studies showing that the proximal presence of a social peer, when imposed noncontingently by the experimenter, decreases cocaine self-administration only modestly [50, 51]. The stronger protective effect of operant “self-administration” of a peer was likely due in part to our use of mutually exclusive choice procedure on each trial. We think it is also likely that volitional social interaction is inherently more rewarding than experimenter-imposed social interaction. Support for this interpretation includes our previous finding that socially isolated rats did not shift their preference from social interaction to methamphetamine even after 6 days of experimenter-imposed group housing with social peers [17].
The translational utility of social-based animal models
Like the literature we cited on social reinforcement-based treatments for addiction in humans, our findings have both heuristic and practical implications. Heuristically, they extend our initially unexpected finding that rats meeting criteria for addiction will nonetheless reliably “self-administer” social reinforcement via peer interaction in preference to the drug [17]. As we have previously discussed [17], the near-universality of this preference in rats (when it is not universal in humans) may be understood in terms of a distinction made by social scientists between hedonic and eudaimonic well-being [52]. Hedonic well-being is pleasure as conventionally understood, and it can potentially be derived from almost any peer interaction (in rats or humans). Eudaimonic well-being is a sense of meaning; it derives from living in accordance with one’s values and fulfilling one’s potential—which, for most people, probably requires a feeling of social belongingness [53]. Not every peer interaction can provide that feeling; some might detract from it [54]. This is probably a cross-species gap in the way social reinforcement is experienced, such that interventions specifically intended to increase eudaimonic well-being in humans [55, 56] will not be testable in laboratory animals. But there may be cross-species homology in the traits underlying proneness to forgo social reward (whether hedonic or eudaimonic) in favor of drug reward. This is where our findings of individual differences may have practical implications. The two-part test of those implications will be to find interventions (e.g., medications) that prevent rats from choosing a drug over delayed social reward or with more effort to obtain it, and then to test those interventions in humans who do not respond to social reinforcement-based treatments (or treatments aimed at eudaimonic well-being) for drug addiction.
Concluding remarks
We showed that immediate social reward prevents cocaine self-administration and relapse in rat addiction models, while delaying or increasing the effort to obtain the social reward reveals individual differences in preference for cocaine, potentially modeling the variability in the effectiveness of social-reinforcement-based behavioral treatments for drug-use disorders in humans. From a translational perspective, the subset of rats who are more prone to resume drug self-administration when social interaction is delayed or requires more effort could be the ideal testing ground for new biomedical treatments.
References
Granfield R, Cloud W. Social context and “natural recovery”: the role of social capital in the resolution of drug-associated problems. Subst Use Misuse. 2001;36:1543–70.
Nutt DJ, King LA, Phillips LD.Independent Scientific Committee on Drugs Drug harms in the UK: a multicriteria decision analysis. Lancet. 2010;376:1558–65.
Marlatt GA, Baer JS, Donovan DM, Kivlahan DR. Addictive behaviors: etiology and treatment. Annu Rev Psychol. 1988;39:223–52.
Pickard H. Responsibility without blame for addiction. Neuroethics. 2017;10:169–80.
Heilig M, MacKillop J, Martinez D, Rehm J, Leggio L, Vanderschuren L. Addiction as a brain disease revised: why it still matters, and the need for consilience. Neuropsychopharmacology. 2021;46:1715–23.
Aklin WM, Wong CJ, Hampton J, Svikis DS, Stitzer ML, Bigelow GE, et al. A therapeutic workplace for the long-term treatment of drug addiction and unemployment: eight-year outcomes of a social business intervention. J Subst Abus Treat. 2014;47:329–38.
Hunt WA, Barnett LW, Branch LG. Relapse rates in addiction programs. J Clin Psychol. 1971;27:455–6.
Silverman K, DeFulio A, Sigurdsson SO. Maintenance of reinforcement to address the chronic nature of drug addiction. Prev Med. 2012;55:S46–53. Suppl
Banks ML, Negus SS. Insights from preclinical choice models on treating drug addiction. Trends Pharm Sci. 2017;38:181–94.
Azrin NH. Improvements in the community-reinforcement approach to alcoholism. Behav Res. Ther. 1976;14:339–48.
Azrin NH, Acierno R, Kogan ES, Donohue B, Besalel VA, McMahon PT. Follow-up results of supportive versus behavioral therapy for illicit drug use. Behav Res. Ther. 1996;34:41–6.
Stitzer ML, Jones HE, Tuten M, Wong C Community reinforcement approach and contingency management interventions for substance abuse. In: Cox WM, Klinger E, editors. Handbook of motivational counseling: goal-based approaches to assessment and intervention with addiction and other problems. Chichester, UK: John Wiley & Sons, Ltd.; 2011.
Lash SJ, Burden JL, Monteleone BR, Lehmann LP. Social reinforcement of substance abuse treatment aftercare participation: impact on outcome. Addict Behav. 2004;29:337–42.
Regier PS, Redish AD. Contingency management and deliberative decision-making processes. Front Psychiatry. 2015;6:76.
Heilig M, Epstein DH, Nader MA, Shaham Y. Time to connect: bringing social context into addiction neuroscience. Nat Rev Neurosci. 2016;17:592–9.
Havassy BE, Hall SM, Wasserman DA. Social support and relapse: commonalities among alcoholics, opiate users, and cigarette smokers. Addict Behav. 1991;16:235–46.
Venniro M, Zhang M, Caprioli D, Hoots JK, Golden SA, Heins C, et al. Volitional social interaction prevents drug addiction in rat models. Nat Neurosci. 2018;21:1520–29.
Venniro M, Russell TI, Ramsey LA, Richie CT, Lesscher HM, Giovanetti SM, et al. Abstinence-dependent dissociable central amygdala microcircuits control drug craving. Proc Natl Acad Sci USA. 2020;117:8126–34.
Ahmed SH, Koob GF. Transition from moderate to excessive drug intake: change in hedonic set point. Science. 1998;282:298–300.
Zimmer BA, Oleson EB, Roberts DC. The motivation to self-administer is increased after a history of spiking brain levels of cocaine. Neuropsychopharmacology. 2012;37:1901–10.
Deroche-Gamonet V, Belin D, Piazza PV. Evidence for addiction-like behavior in the rat. Science. 2004;305:1014–17.
Piazza PV, Deroche-Gamonet V. A multistep general theory of transition to addiction. Psychopharmacology. 2013;229:387–413.
Venniro M, Russell TI, Zhang M, Shaham Y. Operant social reward decreases incubation of heroin craving in male and female rats. Biol Psychiatry. 2019;86:848–56.
Fredriksson I, Venniro M, Reiner DJ, Chow JJ, Bossert JM, Shaham Y. Animal models of drug relapse and craving after voluntary abstinence: a review. Pharm Rev. 2021;73:1050–83.
Venniro M, Shaham Y. An operant social self-administration and choice model in rats. Nat Protoc. 2020;15:1542–59.
Grimm J, Hope B, Wise R, Shaham Y. Neuroadaptation—incubation of cocaine craving after withdrawal. Nature. 2001;412:141–42.
Ahmed SH, Koob GF. Long-lasting increase in the set point for cocaine self-administration after escalation in rats. Psychopharmacology. 1999;146:303–12.
Kawa AB, Valenta AC, Kennedy RT, Robinson TE. Incentive and dopamine sensitization produced by intermittent but not long access cocaine self-administration. Eur J Neurosci. 2019;50:2663–82.
Lenoir M, Serre F, Cantin L, Ahmed SH. Intense sweetness surpasses cocaine reward. PLoS ONE. 2007;2:e698.
Ahmed SH. Trying to make sense of rodents’ drug choice behavior. Prog Neuropsychopharmacol Biol Psychiatry. 2018;87:3–10. Pt A
Caprioli D, Venniro M, Zeric T, Li X, Adhikary S, Madangopal R, et al. Effect of the novel positive allosteric modulator of metabotropic glutamate receptor 2 AZD8529 on incubation of methamphetamine craving after prolonged voluntary abstinence in a rat model. Biol Psychiatry. 2015;78:463–73.
Canchy L, Girardeau P, Durand A, Vouillac-Mendoza C, Ahmed SH. Pharmacokinetics trumps pharmacodynamics during cocaine choice: a reconciliation with the dopamine hypothesis of addiction. Neuropsychopharmacology. 2021;46:288–96.
Core TR. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2018.
Charrad M, Ghazzali N, Boiteau V, Niknafs A. An R package for determining the relevant number of clusters in a data set. J Stat Softw. 2014;61:1–36.
Genolini C, Alacoque X, Sentenac M, Arnaud C. kml and kml3d: R packages to cluster longitudinal data. J Stat Softw. 2015;65:34.
Solinas M, Chauvet C, Thiriet N, El Rawas R, Jaber M. Reversal of cocaine addiction by environmental enrichment. Proc Natl Acad Sci USA. 2008;105:17145–50.
Zlebnik NE, Carroll ME. Prevention of the incubation of cocaine seeking by aerobic exercise in female rats. Psychopharmacology. 2015;232:3507–13.
Thiel KJ, Painter MR, Pentkowski NS, Mitroi D, Crawford CA, Neisewander JL. Environmental enrichment counters cocaine abstinence-induced stress and brain reactivity to cocaine cues but fails to prevent the incubation effect. Addict Biol. 2012;17:365–77.
Badiani A, Belin D, Epstein D, Calu D, Shaham Y. Opiate versus psychostimulant addiction: the differences do matter. Nat Rev Neurosci. 2011;12:685–700.
De Luca MT, Montanari C, Meringolo M, Contu L, Celentano M, Badiani A. Heroin versus cocaine: opposite choice as a function of context but not of drug history in the rat. Psychopharmacology. 2019;236:787–98.
De Pirro S, Galati G, Pizzamiglio L, Badiani A. The affective and neural correlates of heroin versus cocaine use in addiction are influenced by environmental setting but in opposite directions. J Neurosci. 2018;38:5182–95.
Venniro M, Zhang M, Shaham Y, Caprioli D. Incubation of methamphetamine but not heroin craving after voluntary abstinence in male and female rats. Neuropsychopharmacology. 2017;42:1126–35.
Nicolas C, Zlebnik NE, Farokhnia M, Leggio L, Ikemoto S, Shaham Y. Sex differences in opioid and psychostimulant craving and relapse: a critical review. Pharmacol Rev. 2021; Accepted pending revisions.
Nicolas C, Russell TI, Pierce AF, Maldera S, Holley A, You ZB, et al. Incubation of cocaine craving after intermittent-access self-administration: sex differences and estrous cycle. Biol Psychiatry. 2019;85:915–24.
Kerstetter KA, Aguilar VR, Parrish AB, Kippin TE. Protracted time-dependent increases in cocaine-seeking behavior during cocaine withdrawal in female relative to male rats. Psychopharmacology. 2008;198:63–75.
Campbell EJ, Jin S, Lawrence AJ. Environmental enrichment reduces the propensity to relapse following punishment-imposed abstinence of alcohol seeking. Physiol Behav. 2019;210:112638.
Chauvet C, Goldberg SR, Jaber M, Solinas M. Effects of environmental enrichment on the incubation of cocaine craving. Neuropharmacology 2012;63:635–41.
Ewing S, Ranaldi R. Environmental enrichment facilitates cocaine abstinence in an animal conflict model. Pharm Biochem Behav. 2018;166:35–41.
Mazur JE. An adjusting procedure for studying delayed reinforcement. Quantitative analysis of behavior: the effect of delay and of intervening events on reinforcement value. Hillsdale, NJ: Erlbaum; 1987. p. 55–73.
Smith MA. Peer influences on drug self-administration: social facilitation and social inhibition of cocaine intake in male rats. Psychopharmacology. 2012;224:81–90.
Smith MA, Pitts EG. Social preference and drug self-administration: a preclinical model of social choice within peer groups. Drug Alcohol Depend. 2014;135:140–5.
Ryan RM, Deci EL. On happiness and human potentials: a review of research on hedonic and eudaimonic well-being. Annu Rev Psychol. 2001;52:141–66.
Baumeister RF, Leary MR. The need to belong: desire for interpersonal attachments as a fundamental human motivation. Psychol Bull. 1995;117:497–529.
Knapp KS, Brick TR, Bunce SC, Deneke E, Cleveland HH. Daily meaningfulness among patients with opioid use disorder: examining the role of social experiences during residential treatment and links with post-treatment relapse. Addict Behav. 2021;119:106914.
van Agteren J, Bartholomaeus J, Steains E, Lo L, Gerace A. Using a technology-based meaning and purpose intervention to improve well-being: a randomised controlled study. J Happiness Stud. 2021:1–21.
Lubarsky KE, Avery JD. Dignity therapy for alcohol use disorder. Am J Psychiatry. 2016;173:90.
Funding
The authors declare that they do not have any conflicts of interest (financial or otherwise) related to the text of the paper. The research was supported by the Intramural Research Program of NIDA, a grant from NIDA [DA047976] (MV), and the NARSAD Distinguished Investigator Grant Award (YS).
Author information
Authors and Affiliations
Contributions
MV, LVP, DHE and YS contributed to different aspects of the study, including the design and performance of the research, the data analysis, and the write-up of the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Venniro, M., Panlilio, L.V., Epstein, D.H. et al. The protective effect of operant social reward on cocaine self-administration, choice, and relapse is dependent on delay and effort for the social reward. Neuropsychopharmacol. 46, 2350–2357 (2021). https://doi.org/10.1038/s41386-021-01148-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-021-01148-6
This article is cited by
-
Social odor choice buffers drug craving
Neuropsychopharmacology (2024)
-
Acquisition of cocaine reinforcement using fixed-ratio and concurrent choice schedules in socially housed female and male monkeys
Psychopharmacology (2024)
-
A contextualized reinforcer pathology approach to addiction
Nature Reviews Psychology (2023)
-
Wistar rats choose alcohol over social interaction in a discrete-choice model
Neuropsychopharmacology (2023)
-
An operant social self-administration and choice model in mice
Nature Protocols (2023)
