
Abstract
Studies in mice and humans have shown that CD8+ T cell immunosurveillance in non-lymphoid tissues is dominated by resident populations. Whether CD4+ T cells use the same strategies to survey peripheral tissues is less clear. Here, examining the turnover of CD4+ T cells in transplanted duodenum in humans, we demonstrate that the majority of CD4+ T cells were still donor-derived one year after transplantation. In contrast to memory CD4+ T cells in peripheral blood, intestinal CD4+ TRM cells expressed CD69 and CD161, but only a minor fraction expressed CD103. Functionally, intestinal CD4+ TRM cells were very potent cytokine producers; the vast majority being polyfunctional TH1 cells, whereas a minor fraction produced IL-17. Interestingly, a fraction of intestinal CD4+ T cells produced granzyme-B and perforin after activation. Together, we show that the intestinal CD4+ T-cell compartment is dominated by resident populations that survive for more than 1 year. This finding is of high relevance for the development of oral vaccines and therapies for diseases in the gut.
Similar content being viewed by others

Introduction
Studies in mouse models of infection have shown that CD8+ T cells remain in peripheral tissues long after pathogen clearance.1 These long-lived CD8+ T cells have limited potential to recirculate and have been termed resident memory T (TRM) cells.2,3,4 Moreover, CD8+ TRM cells show an extraordinary ability to mount rapid and potent in situ responses after infectious re-exposure.5,6,7 The currently most established markers to identify CD8+ TRM cells in barrier tissues are CD69 and CD103.8,9,10 CD69 is rapidly upregulated after arrival into the tissue,11 and plays a key role preventing tissue egress by antagonizing sphingosine 1-phosphate receptor (S1PR1).12 CD103 (also known as αE integrin) is highly expressed on intraepithelial lymphocytes (IELs) and the heterodimer αEβ7 binds E-cadherin on the surface of epithelial cells,13,14 promoting the accumulation of IELs in the epithelium.
Although CD4+ T cells are more abundant than CD8+ T cells in most peripheral tissues,15 studies to understand TRM cell biology have mainly focused on CD8+ T cells. Over the last decade, CD4+ TRM cells have been identified in the lung,16,17 skin18,19 and reproductive tract.20 However, the CD4+ TRM population seems to be more heterogeneous and functionally plastic compared to CD8+ TRM cells,21,22 and whether CD4+ T cells in peripheral tissues are truly resident, non-circulatory cells is still a matter of debate.23
CD103 as well as CD69 are induced by TGF-β, which is constitutively produced by gut epithelial cells.24 At human mucosal sites, most CD4+ T cells express CD69, but few express CD103 compared to CD8+ TRM cells,15 and both CD103- and CD103+ CD4+ T subsets have been described in different tissues, such as lung9,25 and skin.18 Intestinal CD4+ TRM cells have shown to play a critical role in protection against different pathogens, including C. rodentium26 and Listeria27 in mouse models. Although our knowledge about the role of intestinal CD4+ T-cell effector subsets in the pathogenesis of inflammatory bowel disease (IBD)28,29,30 and coeliac disease31,32 have substantially progressed over the last decade, our current understanding of CD4+ T-cell immunosurveillance and long-term persistence in the human intestine remains incomplete.
We have recently reported that the majority of CD8+ T cells persists for years in human small intestine,8 however, it is still unknown whether CD4+ T cells share these features with their CD8+ counterparts. Here, we present a comprehensive study of the longevity and phenotype of intestinal CD4+ T cells in humans. In a unique transplantation setting we followed the persistence of donor-derived CD4+ T cells in grafted duodenum over time and found that the majority of donor CD4+ T cells are maintained for at least one year in the graft. Furthermore, both CD103- and CD103+ CD4+ T cell populations presented very similar turnover rates, suggesting that both constitute TRM populations. Finally, we showed that the vast majority of both CD103- and CD103+ CD4+ TRM cells were polyfunctional TH1 cells and a fraction produced cytotoxic granules after activation.
Results
Human intestinal CD4+ T cells are phenotypically distinct from their circulating counterparts
To identify CD4+ T cells with a TRM-phenotype in the human small intestine (SI) we first studied the CD4+ T-cell compartment under steady state conditions. For this purpose we collected SI specimens from proximal duodenum-jejunum resections of patients undergoing pancreatic cancer surgery (Whipple procedure, n = 35; mean age 63 yr; 16 female), and from donors and recipients during pancreatic-duodenal Tx (baseline samples, donors: n = 52; mean age 31 yr; 24 female; patients: n = 36; mean age 41 yr; 14 female). All tissue samples were evaluated by experienced pathologists and only histologically normal SI was included. Single-cell suspensions from epithelium (intraepithelial, IE) and enzyme-digested lamina propria (LP) were obtained and analyzed by flow cytometry together with peripheral blood mononuclear cells (PBMCs) from the patients. To characterize the phenotypic profile of SI CD4+ T cells, we performed flow-cytometry analysis using a panel of antibodies that we recently implemented to study SI CD8+ TRM cells.8 CD4+ T cells comprised almost 60% of LP T cells (with a CD4+: CD8+ ratio similar to PB), but constituted only 10% of T cells in the epithelium (Supplementary Fig. S1A-B). The relative distribution of T cell subsets was conserved in mucosal biopsies sampled up to 35 cm apart from the same intestinal resection (Supplementary Fig. S1C).
Applying a dimensionality reduction technique (UMAP, Uniform Manifold Approximation and Projection) on the compiled flow cytometry data we found that all SI CD4+ T cells clustered separate from PB CD4+ T cells (Fig. 1a, left). The vast majority of the SI CD4+ T cells presented a CD45RA− CD45RO+ L-Sel− CCR7− effector memory (TEM) phenotype (Fig. 1a–c). In contrast, PB CD4+ T cells contained a substantial fraction of naïve (TN, CD45RO− CD45RA+ CCR7+ L-Sel+) and central memory (TCM, CD45RO+ CD45RA− CCR7+ L-Sel+) CD4+ T cells (Fig. 1a–c). Virtually all SI CD4+ T cells expressed the TRM marker CD69 whereas all PB CD4+ T cells were CD69-negative (Fig. 1a, c). Intestinal CD4+ T cells (LP and IE) separated into three clusters. The lower cluster was enriched in CD103+KLRG1- cells, whereas the middle and upper clusters contained mainly CD103−KLRG1− and CD103−KLRG1+ cells, respectively (Fig. 1a). Looking at the expression of each individual marker in a larger number of intestinal samples, the population expressing CD103 comprised on average 18% of the LP and 66% of IE CD4+ T cells (Fig. 1c, right), whereas KLRG1 was expressed by 26 and 5% of LP and IE CD4+ T cells, respectively (Fig. 1d). PB CD4+ T cells were completely negative for CD103, however a fraction (mean 19%) of PB CD4+ TEM cells expressed KLRG1 (Fig. 1a, d). PB CD4+ TEM and SI CD4+ T cells showed similar expression of PD1, CD127 (IL-7 receptor- α) and NKG2D. In contrast, CD28 was significantly higher expressed on PB CD4+ TEM cells, whereas CD161 was expressed at higher levels on SI CD4+ T cells (Fig. 1a, d). In addition, the immunomodulatory receptor CD244 (2B4) was expressed higher on the IE subset. In line with other reports,28,33 we also found that CD49a, CXCR6, and the negative regulator CD101 were highly expressed by the SI CD4+ T cells (Supplementary Fig. S1d). Given that one of the SI CD4+ T-cell clusters was enriched in cells expressing the TRM marker CD103 (Fig. 1a), we examined the differential phenotypic profile of CD103+ and CD103− CD4+ T cells in LP and in the epithelium. CD103− CD4+ T cells presented a higher fraction of KLRG1 positive cells in both compartments, while IE CD103+ CD4+ T cells exhibited significantly higher expression of 2B4. For the rest of the markers analyzed, only small differences were found between the CD103+ and CD103- subsets (representative histograms in Fig. 1e and compiled data in Supplementary Fig. S1E).

a UMAP visualization after concatenation of flow-cytometric data from PB (red), LP (gray), and IE (green) CD4+ T cells, as described in ref. 8. Representative of three samples. Map of the clusters and representation of each tissue compartment (left). Overlay of the UMAP clusters and the expression levels for each marker, color-coded based on the median fluorescence intensity values (MFI) (right). b Representative contour plots showing L-selectin and CD45RA expression on PB, LP and IE CD4+ T cells and classification of these cells into Tcm, central memory; Tem, effector memory; TemRA, effector memory re-expressing CD45RA; Tn, naïve. c Phenotypic comparison of total PB CD4+ T cells or d effector memory (EM) PB CD4+ T cells with intestinal LP, and IE CD4+ T cells. Compiled data for each marker is given and black bars indicate mean values. One-way ANOVA with Tukey’s multiple comparisons test. ns, not significant; *P ≤ 0.05; **P ≤ 0.01 ***P ≤ 0.001; ****P ≤ 0.0001. e Representative histograms showing the differential phenotypic profile of intestinal CD103− and CD103+ CD4+ T cells from LP and IE for several TRM-related markers. Mean values and SEM is provided. Compiled data of all the experiments are shown in Figure S1C.
Taken together, these results show that SI CD4+ T cells were clearly different from their blood counterparts, showing a TRM-like surface phenotype (CD69+ CD103+/− CD49a+ CXCR6+ CD101+ CD161+ CD28low).
CD4+ TRM cells persist for >1 yr in the transplanted SI
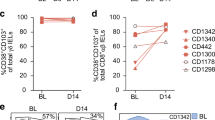
To directly examine the longevity of CD4+ TRM cells in human SI, we assessed the long-term persistence of donor CD4+ T cells in endoscopic biopsies obtained from grafted duodenum at 3, 6 and 52 weeks after pancreatic-duodenal transplantation (Tx) of type I diabetic patients.34 Only patients without histological or clinical signs of rejection were included (n = 32). Most donors and recipients expressed different human leukocyte antigen (HLA) type I molecules rendering it possible to distinguish donor cells from incoming recipient cells in the graft by flow cytometry (Fig. 2a, b). Since CD103 is frequently used as a marker to infer tissue residency, the replacement kinetics of CD103- and CD103+ CD4+ T cells was analyzed independently. At 3 and 6 weeks, LP and IE CD4+ T cells exhibited very low replacement (median >85% donor cells), with no significant differences between the CD103− and CD103+ CD4+ T subsets (Fig. 2b). Importantly, also 1-yr after Tx the majority of SI CD4+ T cells in both the LP and IE compartments were donor-derived, the fraction being slightly higher for LP CD103+ compared to CD103− CD4+ T cells (medians 77 and 60%, respectively). The fact that the majority of CD103− CD4+ T cells were still of donor origin at 1 yr post-Tx suggests that CD103 expression is not absolutely necessary for the persistence of CD4+ TRM cells in human SI. In line with this, the turnover of both IE and LP CD103+ and CD103- cells was highly correlated at 1 yr post-Tx (Fig. 2c). Moreover, at 1 yr post-Tx the CD103- and CD103+ CD4+ T-cell subsets contained a similar (or higher) proportion of donor cells compared to donor CD8+ T cell subsets (Fig. 2d).8 To confirm the persistence of donor CD4+ T cells we performed immunostaining with anti-CD3 and anti-CD4 antibodies combined with fluorescent in situ hybridization probes specific for X/Y-chromosomes on tissue sections where recipients and donors were of different gender and consistently observed donor-derived CD4+ T cells in the graft 1-yr after Tx (Fig. 2e).

a Representative contour plots at 52 weeks (wk) after Tx and b compile data for the fractions of donor-derived CD4+ T cells in LP CD103− (red) and CD103+ (blue) compartments as well as IE CD103− (gray) and CD103+ (green) at 3 (n = 20), 6 (n = 18), and 52 wk after Tx (n = 14) determined by HLA class I expression (as in a). Gray columns indicate median values and lines represent interquartile range. Statistical analysis was performed using two-way ANOVA for repeated measures (RM) with Tukey’s multiple comparisons test. ns, not significant; ***P ≤ 0.001; c Pearson correlation of the percentages of donor-derived cells in the above-mentioned LP and IE CD4+ T cell subsets 1-yr after Tx. Statistics performed using two-tailed P-value (95% confidence interval, n = 14). d Frequencies of persisting donor cells in different subsets of CD4+ and CD8+ T cells from the same biopsies of donor duodenum at 52 wk post-Tx. Data for CD8+ T cells has been previously published.8 Gray bars indicate median values. RM two-way ANOVA. ****P ≤ 0.0001; ns, non-significant. e Representative confocal image of biopsies obtained from donor duodenum (male) of a female patient at one-year post-transplantation. Tissue sections were stained with X/Y chromosome fluorescent in situ hybridization probes (Y, green; X, red) and antibodies against CD4 (red) and CD3 (blue). Hoechst (gray) stains individual nuclei. Scale bars, 50 and 10 µm, respectively.
These results showed that the majority of donor-derived SI CD4+ T cells persisted at least 1 yr (possibly years) in the tissue. However, to exclude effects of the surgical trauma, immunosuppressive treatment and leukocyte chimerism on the SI CD4+ T-cell population, we examined the absolute T-cell counts in SI over time. Serial tissue sections were stained for CD3 and CD8, scanned and counted. The density of CD4+ T cells was determined by subtracting the number of CD8+ cells from the total CD3+ cell count. We found that the overall density of both CD4+ and CD8+ T cells in Tx duodenum was stable throughout the 1-yr follow-up period (Supplementary Fig. S2A, B). Intracellular staining of single cell suspensions from Tx biopsies with the proliferation marker Ki67 showed few Ki67-positive cells among the donor CD4+ T cells (Supplementary Fig. S2C, D). The percentage of Ki67+ CD4+ T cells was similar to that seen in the native duodenum in Tx patients and in steady state controls (Supplementary Fig. S2D), indicating that proliferation did not contribute substantially to the large number of persisting donor CD4+ T cells in transplanted SI. Finally, we confirmed that the CD4+ T cells in the native (recipient) duodenum were exclusively recipient-derived (Supplementary Fig. S3), demonstrating that migration of donor cells out of the graft was not occurring.
In conclusion, these results show that the CD4+ TRM cell population includes both CD103− and CD103+ cells, and that CD4+ TRM cells are at least as persistent as CD103+ CD8+ TRM cells8 in the transplanted SI.
Incoming recipient CD4+ T cells undergo gradual phenotypic changes over time in transplanted duodenum
Transplanted SI gives us a unique opportunity to study the differentiation of recruited incoming CD4+ T cells and whether they acquire a TRM phenotype in SI mucosa. To this end, we compared the expression of TRM associated markers on donor- and recipient- derived LP CD4+ T cells from biopsies of transplanted duodenum over time. Already at 3 wk post-Tx, virtually all recipient LP CD4+ T cells expressed CD69 (Fig. 3a). More than half of recipient CD4+ T cells expressed CD161 at 6 weeks and that was further increased at 1-yr post Tx to similar levels as donor CD4+ T cells (Fig. 3b). CD103 was expressed on a minor subset of recipient-derived CD4+ T cells at both 6 and 52 weeks; slightly lower than that on donor CD4+ T cells (Fig. 3c, e). In contrast, the fraction of KLRG1-positive cells within donor and recipient-derived CD4+ T cells remained almost unchanged (Fig. 3d, e). Similarly to the steady state conditions (Fig. 1a), the majority of the LP CD4+ T cells were CD103− KLRG1− at all the time points regardless of their origin (Fig. 3e). Furthermore, the turnover of donor LP CD103− KLRG1− and CD103− KLRG1+ CD4+ T cells was very similar, evidenced by the high correlation of donor-derived cells within both subsets over time (Fig. 3f).

a–d Representative flow-cytometric analysis for the expression of different TRM associated markers on donor-derived (black) and recipient- derived (red) LP CD4+ T cells from donor duodenum at the indicated weeks (wk) post-Tx. e Distribution of donor (gray) and recipient-derived (red) LP CD4+ T cells isolated from donor duodenum (above) and native duodenum (below, black) according to the expression of CD103 and KLRG1 at the indicated time-points after-Tx. f Pearson correlation of the percentages of donor-derived cells in LP CD103- KLRG1+ and KLRG1- CD4+ T cell subsets over time after Tx. Statistics performed using two-tailed P value (95% confidence interval, n = 32).
Together, we find that recipient CD4+ T cells recruited to the transplanted duodenum rapidly acquire phenotypic features similar to persistent donor CD4+ T cells, suggesting that they gradually differentiate into TRM in situ.
The majority of SI CD4+ T cells exhibits a polyfunctional TH1 profile
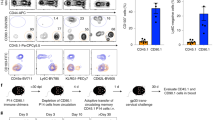
To examine the functional properties of SI CD4+ T cells we studied their cytokine expression profile and ability to produce cytotoxic granules. First, LP CD4+ T cells isolated from histologically normal SI were short-term stimulated with PMA and ionomycin and intracellular staining was performed with antibodies targeting specific cytokines (Supplementary Table S1). By flow-cytometric analysis we found that the majority of the LP CD4+ T cells, both CD103- and CD103+, produced IFN-γ, IL-2 and TNF-α (Fig. 4a). Almost half of the cells produced all these three cytokines simultaneously (Fig. 4b, c), and we did not find significant differences between CD103− KLRG1+ and KLRG1− cells (Supplementary Fig. S4A). In contrast, triple-producing cells constituted only 4% of the memory CD4+ T cells in PB (Fig. 4b, c). Comparing the LP CD103− and CD103+ subsets, we found significantly higher fraction of IL-17 and MIP1-β-producing cells within the CD103+ subset compared to CD103− CD4+ T cell subset (Fig. 4a). Furthermore, CD103+ CD4+ T cells contained a higher fraction of IFN-γ+ IL-17+ double producing cells (Fig. 4d). In contrast, CD103− CD4+ T cells presented higher numbers of IL-13-producing cells than their CD103+ counterparts, whereas comparable expression of IL-10 and IL-22 was found in the two subsets (Fig. 4a).

a Representative contour plots (left) and compiled data (right) showing PMA/ionomycin induced cytokine production by CD103− compared to CD103+ LP CD4+ T cells. P-values of paired t-test are displayed. b IFN-γ, IL-2 and TNF-α production by PB TEM CD4+ T cells and intestinal LP CD103−, LP CD103+ CD4+ T cells. The bars indicate mean values. Statistics performed using one-way ANOVA for each combination of cytokines. c Relative fractions of each cytokine combination indicated in b by PB TEM CD4+ T cells (n = 3), and intestinal LP CD103− (n = 6) and LP CD103+ (n = 6) CD4+ T cells, represented on pie charts with color codes. Mean values of indicated experiments. d Representative contour plots (left) and compiled data (right) showing simultaneous IFN-γ and IL17 expression by LP CD103- and CD103+ CD4+ T cells. Paired t-test.
Murine CD4+ TRM cells exhibit upregulation of granzyme-B upon reactivation with their cognate antigen.35 We therefore analyzed the capacity of SI CD4+ T cells to produce granzyme-B and perforin at the steady state or after stimulation with anti-CD3/CD28 beads. In the absence of stimulation, very few cells expressed these cytolytic proteins (Fig. 5a), however, both LP CD103− and CD103+ subsets increased their expression of granzyme-B and perforin after activation (Fig. 5b, c). We found a significantly higher proportion of granzyme-B producing cells within the LP CD103+ subset as compared to the CD103− CD4+ T cell subset (Fig. 5c). On the other hand, no significant differences were found in the activation-induced production of perforin between either subset (Fig. 5c). Comparing the KLRG1+ and KLRG1− cells in the LP CD103− compartment, we found higher basal levels of granzyme-B among the KLRG1+ cells (Supplementary Fig. S4B), but similar levels of granzyme-B and perforin after stimulation (Supplementary Fig. S4B, C).

Representative flow-cytometric dot plots a, b and compiled data c for the intracellular expression of granzyme-B and perforin in LP CD103− and CD103+ CD4+ T cell subsets without and after stimulation with anti-CD3/CD28 beads for 21 h. Red lines indicate median values. Student’s t-test was applied to compare the expression of cytotoxic mediators by both subsets (black horizontal lines), and by unstimulated versus 21 h stimulated cells (red vertical lines and asterisks). **P ≤ 0.01; ns, not significant.
These data show that the majority of the SI CD4+ TRM cells are polyfunctional TH1 cells, with a large fraction co-producing IFN-γ, IL-2 and TNF-α. A subset of CD4+ TRM cells also produces the cytotoxic proteins granzyme-B and perforin after stimulation.
Discussion
Over the last years it has been demonstrated that immunosurveillance by memory CD8+ T cell in barrier tissues is largely mediated by durable, resident cell populations. However, whether memory CD4+ T cells use similar surveillance strategies is less clear.9,18,23,36 Here, we show that the majority of CD4+ T cells are persistent for at least 1 yr in the human SI mucosa, where they exhibit a polyfunctional TH1 cytokine profile.
There is conflicting evidence with regards to the long-term residency of memory CD4+ T cells in barrier tissues. Studies of CD4+ TRM cells using parabiotic mice have suggested that CD4+ T-cell surveillance in the skin was dependent on continuous recirculation rather than permanent residency.37,38 However, evidence of CD4+ TRM cells persistence has been reported in other peripheral tissues, such as the reproductive mucosa and lung.16,20 Similarly, Beura et al. recently demonstrated that residency is the dominant mechanism of memory CD4+ T-cell immunosurveillance in non-lymphoid tissues, but they did not evaluate the longevity.35 Moreover, in a recent study Klicznik and colleagues discovered a population of skin CD103+ CD69+ CD4+ T cells that were able to downregulate CD69 expression and enter the circulation, indicating that some CD4+ TRM cells may retain migratory potential.39
In mouse models of infection, the number of antigen-specific memory CD4+ T cells in lymphoid and non-lymphoid tissues seem to decline faster than CD8+ T cells,36,40 suggesting that memory CD4+ T cells are less durable. In line with these results, donor CD4+ T cells in lung transplanted patients were more rapidly lost than CD8+ T cells.9 Here, we found that donor CD4+ T cells were maintained in duodenal grafts at equal or even higher numbers than CD103+ CD8+ T cells. At 6 weeks post-transplantation more than 80% of CD4+ T cells were of donor origin and at 1 yr the average of donor CD4+ T cells was still over 60%, but with high variation between grafts. This variation was not likely caused by an increased influx of recipient CD4+ T cells outnumbering persisting donor cells in the graft, as the overall density of CD4+ T cells was unchanged at 1 yr post-transplantation (Figure S2).
Several reports on organ transplantation, including intestinal transplantation,9,41,42 have shown that rejection episodes dramatically increase the replacement kinetics of immune cells. Although we only included patients without histological and/or clinical signs of cellular and antibody-mediated rejection, the patients were not routinely examined by a clinician between 6 and 52 weeks. Therefore, a likely explanation for the large variation in persisting donor CD4+ T cells after 1 yr is that some patients have had undiagnosed intermittent rejection episodes (or other clinical problems) between 6 and 52 weeks after transplantation. Consequently, the grafts with low replacement might more closely represent the steady-state situation in healthy, non-transplanted gut. Together, these results indicate that most CD4+ T cells in human SI under normal conditions are non-circulating, resident cells that most likely perpetuate for years.
Similar to intestinal CD8+ TRM cells,8 we found that virtually all the SI CD4+ T cells expressed CD69 and CD161. However, unlike CD8+ T cells, only a minor fraction of LP CD4+ T cells expressed the αE integrin, CD103. While CD103− CD8+ T cells very rapidly turned over in transplanted duodenum (Fig. 2d and ref. 8), both CD103− and CD103+ donor CD4+ T cells were maintained at high numbers one year after transplantation, with CD103+ cells displaying slightly higher persistence. These results suggest that both CD103− and CD103+ CD4+ T cells constitute resident populations and that retention mechanisms independent of CD103 exist, in line with previous reports of intestinal CD4+ T cells in mice models.27
Like murine CD4+ TRM cells,20,27 the vast majority of LP CD4+ TRM cells exhibited a polyfunctional TH1 profile, producing high amounts of IFN-γ, IL-2 and TNF-α. The fraction of polyfunctional TH1 cells among SI CD4+ T cells was much higher than among memory CD4+ T cells in blood. Furthermore, >40% of the CD4+ TRM cells expressed granzyme-B after stimulation. These results show that SI CD4+ TRM cells, like CD8+ TRM cells, undergo tissue-specific changes that make them poised to provide robust TH1 immunity in response to reinfections.35 In addition to protection against pathogens,27 long-lived CD4+ T cell responses to commensal bacteria have been found during acute gastrointestinal infection with T. gondii.43 Moreover, microbiota-specific CD4+ T cells have been identified in blood and intestinal biopsies from healthy humans,44 indicating that CD4+ TRM cells may actively contribute to intestinal homeostasis through interactions with the microbiota.
We found that a fraction of CD4+ TRM cells produced IL-17. TH17 cells play an important role in intestinal inflammatory disorders,28,29,45 however IL-17 is also critical for maintaining mucosal barrier integrity.45,46 Recently it was reported that, in contrast to inflammatory TH17 cells elicited by pathogens, gut commensal bacteria elicited tissue-resident homeostatic TH17 cells, which showed limited capacity to produce inflammatory cytokines.47 In our study only a very small percentage of TH17 cells co-produced the inflammatory cytokine IFN-γ, suggesting that the majority of SI TH17 cells during homeostasis are non-inflammatory cells that support barrier integrity. However, further studies are needed to understand the role of SI TH17 cells under homeostatic and inflammatory conditions.
Finally, we found, although marginally, that CD103+ TRM cells contained higher fractions of IL-17 single- and IL-17/IFN-γ double-producing cells than their CD103− counterparts. Moreover, CD103- and CD103+ CD4+ T cells also showed subtle phenotypic differences regarding their expression of KLRG1, CD28 and 2B4. However, to what extent the CD103+ and CD103- subsets represent distinct functional compartments needs further investigation.
In conclusion, here we provide evidence that the majority of memory CD4+ T cells in the human SI are resident and may persist in the tissue for >1 year. This indicates that tissue residency represents a major mechanism for CD4+ memory T cell immunosurveillance in the human SI and has several important clinical implications. Effective vaccines depend on durable adaptive immune responses. We have previously shown that plasma cells48 and CD8+ T cells8 are long-lived in the SI. The finding that the intestinal immune system fosters also persisting CD4+ memory T cells is important knowledge in the search for strategies to develop long-lasting oral vaccines. Persisting CD4+ T cells may also play an important role in the chronicity of CD4+ T-cell mediated inflammatory diseases, such as celiac disease31 and inflammatory bowel disease.28 Furthermore, we have recently shown that intestinal CD4+ T cells survive conditioning regimens in allogeneic stem cell transplantation and are present in intestinal lesions of graft versus host disease (GVHD) (Divito et al. J Clin Invest, in press), suggesting that persistent host CD4+ T cells may play a role in GVHD pathology. Longevity of intestinal CD4+ T cells must therefore be taken into consideration in therapeutic interventions targeting pathogenic CD4+ T cells to treat immune-mediated disorders.
Methods
Patient samples
Small intestinal samples were either obtained during pancreatic cancer surgery (Whipple procedure, n = 35; mean age 63 yr; range 40–81 yr; 16 female), or from donors and/or patients during pancreas-duodenum transplantation (donors: n = 52; mean age 31 yr; range 5–55 yr; 24 female; patients: n = 36; mean age 41 yr; range 25–60 yr; 14 female) as described previously.8 Cancer patients receiving neoadjuvant chemotherapy were excluded from the study. Endoscopic biopsies from donor and patient duodenum were collected at 3, 6 and 52 weeks after transplantation. All tissue specimens were evaluated blindly by experienced pathologists, and only material with normal histology was included.49 All transplanted patients received a standard immunosuppressive regimen,34 and patients showing clinical or histological signs of rejection or other complications, as well as patients presenting pre-transplant or de novo donor specific antibodies (DSA) were excluded from the study. Blood samples were collected at the time of the surgery and buffy coats obtained from healthy donors (Oslo University Hospital). All participants gave their written informed consent. The study was approved by the Regional Committee for Medical Research Ethics in Southeast Norway and complies with the Declaration of Helsinki.
Preparation of intestinal and peripheral blood single-cell suspensions
Intestinal resections were opened longitudinally and rinsed with PBS, and mucosa was dissected in strips off the submucosa. For microscopy, small mucosal pieces were fixed in 4% formalin and embedded in paraffin according to standard protocols. Intestinal mucosa was washed 3 times in PBS containing 2 mM EDTA and 1% FCS at 37 °C with shaking for 20 min and filtered through nylon 100-µm mesh to remove epithelial cells. Epithelial fractions in each washing step were pooled and filtered through 100-µm cell strainers (BD, Falcon). Epithelial cells in the EDTA fraction were depleted by incubation with anti-human epithelial antigen antibody (clone Ber-EP4, Dako) followed by anti-mouse IgG dynabeads (ThermoFisher) according to the manufacture’s protocol. De-epithelialized LP was minced and digested in complete Roswell Park Memorial Institute (RPMI) medium (supplemented with 1% Pen/Strep) containing 0.25 mg/mL Liberase TL and 20 U/mL DNase I (both from Roche), stirring at 37 °C for 1 h. Digested tissue was filtered twice through 100-µm cell strainers and washed tree times in PBS. Purity of both IE and LP fractions was checked by flow-cytometry.8 Intestinal biopsies from transplanted patients were processed in the same way. PBMCs were isolated by Ficoll-based density gradient centrifugation (Lymphoprep™, Axis-Shield).
Flow cytometry
Single cell suspensions of intestinal LP and IE fractions and PBMCs were stained using different multicolor combinations of directly conjugated monoclonal antibodies (Supplementary Table S1). To assess the expression of L-Selectin on digested tissue, cells were rested for 12 h at 37 °C before the immunostaining. Replacement of donor cells in duodenal biopsies of HLA mismatched transplanted patients was assessed using different HLA type I allotype-specific antibodies targeting donor- and/or recipient-derived cells, and stroma cells were used as a control of specific staining. Dead cells were excluded based on propidium iodide staining (Molecular Probes, Life Technologies). For analysis of cytokine production, LP and IE cell suspensions were stimulated for 4 h with control complete medium (RPMI supplemented with 10% FCS, 1% Pen/Strep) or phorbol-12-myristate-13-acetate PMA (1.5 ng/mL) and ionomycin (1 µg/mL; both from Sigma-Aldrich) in the presence of monensin (Golgi Stop, BD Biosciences) added after 1 h of stimulation to allow intracellular accumulation of cytokines. Cells were stained using the BD Cytofix/Cytoperm kit (BD Biosciences) according to the manufacturer’s instructions and stained with antibodies against several cytokines (Supplementary Table S1). For detection of cytotoxic granules, LP and IE cells were activated for 21 h with anti-CD3/CD28 beads (Dynabeads, ThermoFisher) or control complete medium. For detection of intranuclear Ki67 expression the FoxP3/transcription factor staining buffer set was used according to the manufacturer’s instructions. eFluor-450 or eFluor-780 fixable viability dyes (eBioscience) were used prior any intracellular/intranuclear staining procedure. All samples were acquired on LSR Fortessa flow cytometer (BD Biosciences), using FACSDiva software (BD Biosciences). Single stained controls were prepared for compensation (UltraComp eBeads™, eBioscience), and gates were adjusted by comparison with FMO controls or matched isotype controls. Flow cytometry data were analyzed using FlowJo 10.4.2 (Tree Star). For Fig. 1a, the expression of 16 phenotypic markers was analyzed at the single cell-level and compared for CD4+ T cells in PB, LP and IE (n = 3) using the merge and calculation functions of Infinicyt software (Cytognos). The population within the CD4+ T-cell gate was down-sampled for each compartment and exported to a new file as in,8 and then concatenated and subjected to UAMP analysis using the plugin integrated in FlowJo 10.5.3. All experiments were performed at the Flow Cytometry Core Facility, Oslo University Hospital.
Microscopy
Analysis of chimerism was performed as described previously.48 Briefly, formalin-fixed 4-μm sections were washed sequentially in xylene, ethanol, and PBS. Heat-induced epitope retrieval was performed by boiling sections for 20 min in Dako buffer. Sections were incubated with CEP X SpectrumOrange/Y SpectrumGreen DNA Probes (Abbott Molecular Inc.) for 12 h at 37 °C before immunostaining according to standard protocol with anti-CD3 (Polyclonal; Dako), anti-CD4 (clone 1F6, Leica Biosystems) and secondary antibodies targeting rabbit IgG or mouse IgG2b conjugated to Alexa Fluor 647 and 555, respectively. Laser scanning confocal microscopy was performed on an Olympus FV1000 (BX61WI) system. Image z stacks were acquired at 1-μm intervals and combined using the Z project max intensity function in Image J (National Institutes of Health), and all microscopy images were assembled in Photoshop and Illustrator CC (Adobe).
CD8 and CD3 immunoenzymatic staining was performed on formalin-fixed 4-μm sections, dewaxed in xylene and rehydrated in ethanol, and prepared with Vulcan Fast red kit (Biocare Medical) following standard protocols. In brief, heat-induced antigen retrieval was performed in Tris/EDTA pH9 buffer (EnVision FLEX Dako kit, K8010), followed by staining with primary antibody (CD8 clone 4B11, Novocastra or CD3, polyclonal, Dako), secondary anti- mouse AP-conjugated antibody and incubation with substrate (Fast red chromogen, Biocare Medical). Slides were counterstained with hematoxylin and excess of dye was removed by bluing in ammoniac water. Tissue sections were scanned using Pannoramic Midi slide scanner (3DHISTECH) and counts generated with QuPath software.50
Statistical analysis
Statistical analyses were performed in Prism 8 (GraphPad Software). To assess statistical significance among SI CD4+ T cell subsets, data were analyzed by one-way ANOVA (standard or repeated measures, RM-ANOVA) followed by Tukey’s multiple comparison tests. Replacement data and distribution of CD4+ T cell subsets at different time points were analyzed by two-way ANOVA matching across subsets followed by Tukey’s multiple comparison tests. Correlations between replacement kinetics of different CD4+ T cell subsets were calculated using Pearson correlation with two-tailed P-value (95% confidence interval). P-values of <0.05 were considered significant.
References
Masopust, D., Vezys, V., Marzo, A. L. & Lefrancois, L. Preferential localization of effector memory cells in nonlymphoid tissue. Science 291, 2413–2417 (2001).
Masopust, D. & Soerens, A. G. Tissue-resident T cells and other resident leukocytes. Annu. Rev. Immunol. 37, 521–546 (2019).
Mueller, S. N. & Mackay, L. K. Tissue-resident memory T cells: local specialists in immune defence. Nat. Rev. Immunol. 16, 79–89 (2016).
Szabo, P. A., Miron, M. & Farber, D. L. Location, location, location: tissue resident memory T cells in mice and humans. Sci. Immunol. 4, 9673 (2019).
Schenkel, J. M., Fraser, K. A., Vezys, V. & Masopust, D. Sensing and alarm function of resident memory CD8(+) T cells. Nat. Immunol. 14, 509–513 (2013).
Park, S. L. et al. Local proliferation maintains a stable pool of tissue-resident memory T cells after antiviral recall responses. Nat. Immunol. 19, 183–191 (2018).
Beura, L. K. et al. Intravital mucosal imaging of CD8(+) resident memory T cells shows tissue-autonomous recall responses that amplify secondary memory. Nat. Immunol. 19, 173–182 (2018).
Bartolome-Casado, R., et al. Resident memory CD8 T cells persist for years in human small intestine. J. Exp. Med. 216, 2412–2426 (2019).
Snyder, M. E., et al. Generation and persistence of human tissue-resident memory T cells in lung transplantation. Sci. Immunol. 4, 5581 (2019).
Mackay, L. K. et al. The developmental pathway for CD103(+)CD8+ tissue-resident memory T cells of skin. Nat. Immunol. 14, 1294–1301 (2013).
Klonowski, K. D. et al. Dynamics of blood-borne CD8 memory T cell migration in vivo. Immunity 20, 551–562 (2004).
Skon, C. N. et al. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8(+) T cells. Nat. Immunol. 14, 1285 (2013).
Cepek, K. L. et al. Adhesion between epithelial cells and T lymphocytes mediated by E-cadherin and the alpha E beta 7 integrin. Nature 372, 190–193 (1994).
Schon, M. P. et al. Mucosal T lymphocyte numbers are selectively reduced in integrin alpha E (CD103)-deficient mice. J. Immunol. 162, 6641–6649 (1999).
Sathaliyawala, T. et al. Distribution and compartmentalization of human circulating and tissue-resident memory T cell subsets. Immunity 38, 187–197 (2013).
Teijaro, J. R. et al. Cutting edge: tissue-retentive lung memory CD4 T cells mediate optimal protection to respiratory virus infection. J. Immunol. 187, 5510–5514 (2011).
Hondowicz, B. D. et al. Interleukin-2-dependent allergen-specific tissue-resident memory cells drive asthma. Immunity 44, 155–166 (2016).
Watanabe, R. et al. Human skin is protected by four functionally and phenotypically discrete populations of resident and recirculating memory T cells. Sci. Transl. Med. 7, 279ra239 (2015).
Glennie, N. D. et al. Skin-resident memory CD4+ T cells enhance protection against Leishmania major infection. J. Exp. Med. 212, 1405–1414 (2015).
Iijima, N. & Iwasaki, A. T cell memory. A local macrophage chemokine network sustains protective tissue-resident memory CD4 T cells. Science 346, 93–98 (2014).
Becattini, S. et al. T cell immunity. Functional heterogeneity of human memory CD4(+) T cell clones primed by pathogens or vaccines. Science 347, 400–406 (2015).
Brucklacher-Waldert, V., Carr, E. J., Linterman, M. A. & Veldhoen, M. Cellular plasticity of CD4+ T cells in the intestine. Front. Immunol. 5, 488 (2014).
Carbone, F. R. & Gebhardt, T. Should I stay or should I go-reconciling clashing perspectives on CD4(+) tissue-resident memory T cells. Sci. Immunol. 4, 5595 (2019).
Zhang, N. & Bevan, M. J. Transforming growth factor-beta signaling controls the formation and maintenance of gut-resident memory T cells by regulating migration and retention. Immunity 39, 687–696 (2013).
Oja, A. E. et al. Trigger-happy resident memory CD4(+) T cells inhabit the human lungs. Mucosal Immunol. 11, 654–667 (2018).
Bishu, S., et al. Citrobacter rodentium induces tissue-resident memory CD4(+) T cells. Infect. Immun. 87, e00295-19 (2019).
Romagnoli, P. A. et al. Differentiation of distinct long-lived memory CD4 T cells in intestinal tissues after oral Listeria monocytogenes infection. Mucosal Immunol. 10, 520–530 (2017).
Zundler, S., et al. Hobit- and Blimp-1-driven CD4(+) tissue-resident memory T cells control chronic intestinal inflammation. Nat. Immunol. 20, 288–300 (2019).
Kleinschek, M. A. et al. Circulating and gut-resident human Th17 cells express CD161 and promote intestinal inflammation. J. Exp. Med. 206, 525–534 (2009).
Lamb, C. A. et al. alphaEbeta7 integrin identifies subsets of pro-inflammatory colonic CD4+ T lymphocytes in ulcerative colitis. J. Crohns Colitis 11, 610–620 (2017).
Risnes, L. F. et al. Disease-driving CD4+ T cell clonotypes persist for decades in celiac disease. J. Clin. Invest. 128, 2642–2650 (2018).
Christophersen, A. et al. Distinct phenotype of CD4(+) T cells driving celiac disease identified in multiple autoimmune conditions. Nat. Med. 25, 734–737 (2019).
Kumar, B. V. et al. Human tissue-resident memory T cells are defined by core transcriptional and functional signatures in lymphoid and mucosal sites. Cell Rep. 20, 2921–2934 (2017).
Horneland, R. et al. Pancreas transplantation with enteroanastomosis to native duodenum poses technical challenges–but offers improved endoscopic access for scheduled biopsies and therapeutic interventions. Am. J. Transplant. 15, 242–250 (2015).
Beura, L. K., et al. CD4+ resident memory T cells dominate immunosurveillance and orchestrate local recall responses. J. Exp. Med. 216, 1214–1229 (2019).
Homann, D., Teyton, L. & Oldstone, M. B. Differential regulation of antiviral T-cell immunity results in stable CD8+ but declining CD4+ T-cell memory. Nat. Med. 7, 913–919 (2001).
Gebhardt, T. et al. Different patterns of peripheral migration by memory CD4+ and CD8+ T cells. Nature 477, 216–219 (2011).
Collins, N. et al. Skin CD4(+) memory T cells exhibit combined cluster-mediated retention and equilibration with the circulation. Nat. Commun. 7, 11514 (2016).
Klicznik, M. M., et al. Human CD4+CD103+ cutaneous resident memory T cells are found in the circulation of healthy individuals. Sci. Immunol. 4, 8995 (2019).
Cauley, L. S. et al. Cutting edge: virus-specific CD4+ memory T cells in nonlymphoid tissues express a highly activated phenotype. J. Immunol. 169, 6655–6658 (2002).
Zuber, J., et al. Bidirectional intragraft alloreactivity drives the repopulation of human intestinal allografts and correlates with clinical outcome. Sci. Immunol. 1, 3732 (2016).
Eguiluz-Gracia, I. et al. Long-term persistence of human donor alveolar macrophages in lung transplant recipients. Thorax 71, 1006–1011 (2016).
Hand, T. W. et al. Acute gastrointestinal infection induces long-lived microbiota-specific T cell responses. Science 337, 1553–1556 (2012).
Hegazy, A. N. et al. Circulating and tissue-resident CD4(+) T cells with reactivity to intestinal microbiota are abundant in healthy individuals and function is altered during inflammation. Gastroenterology 153, 1320–1337 e1316 (2017).
Ouyang, W., Kolls, J. K. & Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 28, 454–467 (2008).
Martinez-Lopez, M. et al. Microbiota sensing by mincle-Syk axis in dendritic cells regulates interleukin-17 and -22 production and promotes intestinal barrier integrity. Immunity 50, 446–461 e449 (2019).
Omenetti, S., et al. The intestine harbors functionally distinct homeostatic tissue-resident and inflammatory Th17 cells. Immunity 51, 77–89 (2019).
Landsverk, O. J. et al. Antibody-secreting plasma cells persist for decades in human intestine. J. Exp. Med. 214, 309–317 (2017).
Ruiz, P. et al. International grading scheme for acute cellular rejection in small-bowel transplantation: single-center experience. Transplant. Proc. 42, 47–53 (2010).
Bankhead, P. et al. QuPath: open source software for digital pathology image analysis. Sci. Rep. 7, 16878 (2017).
Acknowledgements
We are grateful to the staff at the Endoscopy Unit and the surgical staff; Christian Naper, Institute of Immunology, for providing HLA typing; the Confocal Microscopy and Flow Cytometry Core Facilities; all at Oslo University Hospital, Rikshospitalet. This work was partly supported by the Research Council of Norway through its Centres of Excellence funding scheme (project number 179573/V40) and by grant from the South Eastern Norway Regional Health Authority (project number 2015002).
Author information
Authors and Affiliations
Contributions
R.B.-C., O.J.B.L., E.S.B., and F.L.J. conceived the project. R.B.-C., O.J.B.L., and S.K.C. processed samples, designed and performed experiments, and analyzed data. R.B.-C. prepared figures. F.S. and K.T.H. assisted with experiments and data analysis. S.Y. and R.H. coordinated recruitment of patients and collection of biopsies. S.Y., R.H., O.Ø., and E.M.A. performed surgery and provided samples. L.A. performed endoscopy and provided endoscopic biopsies. R.B.-C. and F.L.J. wrote the manuscript. O.J.B.L., F.S., and E.S.B. contributed to writing the manuscript. E.S.B. and F.L.J. supervised the study.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Bartolomé-Casado, R., Landsverk, O.J.B., Chauhan, S.K. et al. CD4+ T cells persist for years in the human small intestine and display a TH1 cytokine profile. Mucosal Immunol 14, 402–410 (2021). https://doi.org/10.1038/s41385-020-0315-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41385-020-0315-5
This article is cited by
-
Tissue-resident memory T cells: decoding intra-organ diversity with a gut perspective
Inflammation and Regeneration (2024)
-
Tolerance-inducing therapies in coeliac disease — mechanisms, progress and future directions
Nature Reviews Gastroenterology & Hepatology (2024)
-
Tango of B cells with T cells in the making of secretory antibodies to gut bacteria
Nature Reviews Gastroenterology & Hepatology (2023)
-
CD4+ T cell memory
Nature Immunology (2023)
-
Mucosal vaccines — fortifying the frontiers
Nature Reviews Immunology (2022)
