
Abstract
Interleukin(IL)-1β, a pro-inflammatory cytokine, was elevated and participates in periodontitis. Not only the link between IL-1β and periodontitis was proved by clinical evidence, but also the increased IL-1β triggers a series of inflammatory reactions and promotes bone resorption. Currently, IL-1β blockage has been therapeutic strategies for autoimmune and autoinflammatory diseases such as rheumatoid arthritis, cryopyrin-associated periodic syndromes, gout and type II diabetes mellitus. It is speculated that IL-1β be a potential therapeutic target for periodontitis. The review focuses on the production, mechanism, present treatments and future potential strategies for IL-1β in periodontitis.
Similar content being viewed by others

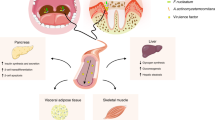

Clinical evidence regarding interleukin(IL)-1β
Periodontitis is an inflammatory disease that infects tooth supporting tissue and eventually induces tooth loss. At present, the aetiology of periodontitis is characterised by both dysbiosis of the host and oral microbiota.1 Various mediators affect the balance in chronic inflammation. The pro-inflammatory cytokines include IL-1β, tumour necrosis factor (TNF)-α, IL-6 and IL-17, which lead to periodontal inflammation and tissue injury. Another group includes regulatory cytokines such as IL-10.2,3 IL-1β is a strong stimulator of periodontal tissue destruction. The properties of IL-1β include promoting bone resorption and inducing the production of tissue-degrading proteinases.4 In addition to periodontitis, pro-inflammatory cytokines such as TNF-α, IL-1β and IL-6 modulate systemic diseases.3 Correspondingly, cytokine-based strategies have the potential to improve both periodontitis and systemic health. This review focuses on new cytokine-based strategies related to IL-1β in periodontitis.
As a pro-inflammatory cytokine, IL-1β participates in inflammation, immune regulation and bone resorption in periodontitis. The clinical findings provide strong evidence about its significance. The biological effects of IL-1β depend on its tissue concentration, which is elevated in periodontitis. Increased levels of IL-1β are frequently detected in the saliva and gingival crevicular fluid (GCF) of patients with periodontitis compared with healthy controls.5,6 Patients with deeper pocket depths and more severe bleeding on probing (BOP) have increased levels of GCF IL-1β.7,8 Moreover, serum IL-1β in patients with chronic periodontitis also reaches a high level, inducing a systemic effect.9,10 This finding demonstrates that IL-1β is a possible causal link between periodontitis and systemic diseases such as cardiovascular diseases.11 The nod-like receptor protein-3 (NLRP3) inflammasome signalling pathway, which contributes to the activation of IL-1β, is involved in periodontal pathogenesis. The mRNA expression of the NLRP3 inflammasome complex was enhanced both in gingivitis and in periodontitis.12
There are significant increases in the salivary levels of NLRP3, apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC), and IL-1β in chronic periodontitis and aggressive periodontitis.13
Additional evidence is that polymorphisms in IL-1 genes might be associated with the risk of periodontitis. Numerous studies have reported that the IL-1β polymorphisms affect the susceptibility to periodontal diseases and their progression. Moreover, the polymorphisms in the IL-1β gene might be associated with increased periodontitis severity.14 The studies focus on the polymorphisms leading to transitions between C and T at three positions, 3954/3 953(C → T, rs1143634), −511(C → T, rs16944), and −31(T → C, rs1143627). IL-1β rs1143634 has long been reported as a risk factor in the progression of periodontitis.15,16,17,18,19 However, some studies do not support such an association,20,21 especially in aggressive periodontitis.22 Recent meta-analyses demonstrated that the IL-1β rs1143634 polymorphism is associated with periodontitis.23,24 Individuals with IL-1β rs1143634 also show high levels of IL-1β in the GCF,25 which adds to the evidence that IL-1β rs1143634 might be a genetic risk factor for periodontitis.
The association between two other polymorphisms and periodontitis is comparably weak. The IL-1β rs16944 polymorphism is associated with susceptibility to periodontitis in the European20 and the Chinese population.26 However, another report showed no significant association between the IL-1β rs16944 polymorphism and CP susceptibility.27 The link between the IL-1β rs1143627 polymorphism and susceptibility to periodontitis is even weaker. The studies show that the IL-1β rs1143627 polymorphism is not related to periodontal disease susceptibility.18,28
Clinical evidence verifies that periodontitis leads to excessive IL-1β production locally (GCF and saliva) and systemically (serum). The mechanism of IL-1β production is important for final control.
The production of IL-1β in periodontitis
IL-1β is mainly expressed by macrophages and dendritic cells. However, gingival fibroblasts, periodontal ligament cells, and osteoblasts can also secrete IL-1β.29 The secretion of IL-1β is unique. The cytokine is first produced as a proprotein, which is subsequently proteolyzed into its active form by caspase-1. The inactive precursor, pro-IL-1β, is produced in response to pathogen-associated molecular patterns [PAMPs, e.g., lipopolysaccharide (LPS)] or damage-associated molecular patterns (DAMPs, e.g., HMGB1, ATP). PAMPs and DAMPs function through pattern recognition receptors (PRRs), e.g., Toll-like receptors (TLRs), to activate the signalling adaptor myeloid differentiation primary response 88 (MyD88). MyD88 activation results in the degradation of IκB as a prerequisite for the release of NF-κB dimers and hence promotes pro-IL-1β expression.30
Pro-IL-1β is produced as biologically inactive forms that must be proteolytically cleaved to acquire functional activity.31 Caspase-1, originally recognised as an IL-1-converting enzyme, is able to cleave pro-IL-1β into active IL-1β, leading to the final secretion. Caspase-1 undergoes activation by a multi-molecular assembly known as the inflammasome. Inflammasomes are composed of the sensor (PRRs including NLRP1, NLRP3, NLRC4, Pyrin and AIM2), the adaptor ASC (apoptosis-associated spec-like protein containing a CARD), and caspase-1.32,33 To date, most investigations have focused on the NLRP3 inflammasome, which is involved in various inflammatory diseases.33,34,35 The NLRP3 inflammasome is activated in response to a variety of PAMPs and DAMPs. In macrophages, NLRP3 inflammasome activation is proposed to be a two-signal pathway. The first signal is priming, which is induced by microbial or endogenous stimuli that promote NLRP3 and pro-IL-1β expression through the activation of NF-κB. The second signal is activation, which is triggered by exogenous ATP (eATP), pore-forming toxins and particulate matter sensors, which activate the NLRP3 inflammasome and caspase-1 and cleave pro-IL-1β into its mature form. The protein gasdermin is another substrate of active caspase-1. Active caspase-1 cleaves gasdermin at the N terminus and C terminus. The N terminus of gasdermin forms pores on cell membranes and subsequently induces pyroptosis.32
In recent decades, the production of IL-1β and pyroptotic processes has been studied in the field of periodontitis. The periodontal pathogens belong to PAMPs that initiate IL-1β production. In macrophages, leucocytes and gingival fibroblasts, Porphyromonas gingivalis (P. gingivalis) is able to activate caspase-1, IL-1β and IL-18. P. gingivalis fimbriae, LPS and DNA act as PAMPs and are recognised by several surface and cytosolic PRRs, e.g., TLRs.36 However, different cell types vary in their priming and activating pathways. Macrophages37,38 and gingival epithelial cells39 participate in two signalling pathways. They first promote NLRP3 and pro-IL-1β expression in response to P. gingivalis. Subsequently, the binding of eATP to the P2X7 receptor, an ion-gated channel, is required for the maturation of IL-1β and final secretion.40 However, human monocytes require only one signal for secreting IL-1β after stimulation with TLR2 or TLR4 receptors due to the endogenous release of ATP and the activation of the P2X7 receptor.41 Human gingival fibroblasts are special in their response to P. gingivalis for the maturation of IL-1β. Supragingival biofilms could enhance caspase 1 activation and the expression of IL-1β and IL-18 in gingival fibroblasts.42 However, subgingival biofilms, including P. gingivalis, downregulate NLRP3 and IL-1β.43 It was suggested that hypoxic conditions were essential in the maturation of IL-1β.44 The diagram is shown in Fig. 1.

Signal 1 is a priming signal. One of pathogen-associated molecular patterns, e.g., P. gingivalis, activates the transcriptional factor NF-κB and thus increases the production of NLRP3, pro-IL-1β and pro-IL-18. Signal 2 is an activation signal. One of pathogen-associated molecular patterns, such as ATP, activates the NLRP3 inflammasome via the P2X7 receptor. Hypoxia helps to activate the NLRP3 inflammasome. Activated caspase-1 produces the active form of IL-1β and IL-18. Activated caspase-1 also cleaves gasdermin D at the N terminus, thus forming pores and inducing the pyroptosis of cells.
Actinobacillus actinomycetemcomitans (A. actinomycetemcomitans), the pathogen of aggressive periodontitis, upregulates IL-1β expression in human mononuclear leucocytes45 and macrophages.46 Leukotoxin, an important virulence factor that targets leucocytes, rapidly activates caspase-1 and thus induces a massive secretion of IL-1β in human monocytes and macrophages.47,48 The NLRP345,49 and AIM2 inflammasomes,50 reactive oxygen species and cathepsin B51 might be involved in the process. However, the exact signalling pathways are not clear and require further investigation.
The role of IL-1β in periodontitis
After secretion, the accumulated IL-1β triggers a series of inflammatory reactions and participates in the pathology of periodontitis.52 Generally, IL-1β in the inflammatory site accounts for increased local blood flow, leucocyte recruitment and neutrophil infiltration.53 Furthermore, IL-1β acts as a strong stimulator of bone resorption, making it a featured cytokine in periodontitis. The major mechanisms are shown in Fig. 2.

IL-1β promotes the secretion of MMPs, RANKL, PGE2, IL-6, IL-8, etc., which promotes osteoclastogenesis.
IL-1β increases the expression of collagenolytic enzymes, matrix metalloproteinases (MMPs), which contribute to extracellular matrix degradation and in turn lead to bone resorption and tissue destruction.53,54 MMP-9 is an important indicator of the severity and progression of periodontitis.55,56,57,58 IL-1β upregulates MMP-9 expression in various cell types involved in periodontal inflammation, including osteoblasts, osteoclasts, neutrophils, and cementoblasts.59,60 In addition, IL-1β stimulated the production of other MMPs, e.g., MMP-1 and/or MMP-3, in human periodontal ligament cells and gingival fibroblast cells.61,62,63
The receptor activator of nuclear factor kappa-B ligand (RANKL) and osteoprotegerin (OPG) system regulates osteoclastogenesis. Increased RANKL and decreased OPG is a feature of periodontitis.64 IL-1β upregulates RANKL and thus stimulates osteoclastogenesis.65
RANKL is a member of the membrane-associated TNF ligand superfamily and is detected on osteoblasts, bone marrow stromal cells, activated T cells, periodontal ligament fibroblasts, cementoblasts and endothelial cells.66,67,68,69
RANKL binds its receptor, receptor activator of NF-κB (RANK), expressed on the osteoclast precursor cells, resulting in the activation of the osteoclast.70 The binding of RANKL to RANK finally activates the transcription of NFATc1, c-Fos and NF-κB-related genes, which are upregulated in periodontitis71,72 and modulate the proliferation, differentiation and activation of osteoclasts.73,74 IL-1β also regulates the production of OPG, which is a soluble receptor for RANKL that prevents it from binding to RANK.75,76 The RANK/RANKL/OPG system is influenced by IL-1β and has been associated with bone metabolism.77
Other than these direct effects, some indirect effects are influenced by IL-1β. IL-1β increases PGE2 synthesis in fibroblast cells.61,75 PGE2 induces the expression of RANKL as well.78 IL-1β increases the expression of CX3CL1, which mediates the migration of osteoclast precursors under the osteoblast layer and finally leads to osteoclastogenesis.79 In response to IL-1β, the fibroblasts in the periodontium produce pro-inflammatory mediators, e.g., IL-6, IL-8,80,81,82 which are reported to stimulate bone resorption.83
In brief, IL-1β has a long-lasting effect on osteoclastogenesis, which leads to bone resorption.65,67
The influence of current therapies on IL-1β
The treatments for periodontitis consist of scaling and root planing (SRP), surgery and some adjunctive therapies, e.g., antibiotics, laser therapy or antimicrobial photodynamic therapy (aPDT).84 The therapies not only improve clinical parameters but also influence the levels of cytokines. However, conventional SRP and surgery are not efficient in reducing the IL-1 levels. Some adjunctive therapies have unexpected effects.
SRP and surgery
SRP is a cost-effective, minimally invasive and non-surgical treatment to prevent and/or control periodontal diseases. However, the mechanical treatment may be incomplete in eliminating the pathogenic microorganisms.85,86 The levels of pro-inflammatory cytokines may be continuously high after treatment. Previous studies found that SRP did not significantly reduce the IL-1 levels.87,88 Some recent studies also show that the IL-1β level is decreased after SRP treatment.89,90 SRP results in a decrease in the level of IL-1β in GCF or saliva. However, the amounts are still higher than in the healthy group.91,92 Furthermore, an obvious increase in the IL-1β level is detected even after papillary flap debridement, which suggests that trauma and wound healing result in prolonged production of IL-1β.93
Due to the difficult access by SRP, the removal of plaque and infectious cells can be impaired in some sites. Although SRP treatment is effective in reducing clinical parameters, it is insufficient in the anti-inflammatory treatment for increased mediators. Thus, some adjuvant therapies have been analysed for this purpose.
Antibiotics
The local and systemic administration of antibiotics is an adjunctive treatment for periodontitis. Drugs such as minocycline, doxycycline, roxithromycin, amoxicillin and metronidazole are typically used.94 Clinical studies have shown that antibiotics promote clinical parameters. However, its effect on IL-1β is limited.
The local administration of minocycline microspheres in periodontal pockets during periodontal maintenance significantly reduces IL-1β in GCF at 6 months.94 However, there are no changes over 24 months.95 The systemic administration of amoxicillin and metronidazole is an adjunctive treatment for aggressive periodontitis. This treatment reduces the IL-1β level in GCF at 3 months,96 but it has no effect at the 6-month follow-up.97,98 Roxithromycin therapy is applied in cyclosporine-A-induced gingival overgrowth. However, there is no effect on the level of IL-1β.99 Furthermore, antibiotics at or below minimal inhibitory concentrations increase IL-1β secretion in macrophages by inducing the shedding of LPS by P. gingivalis.100
Laser therapy
Lasers could be used as a monotherapy, as an adjunct to SRP, or in a surgery for incision. Laser therapy provided minimally invasive soft tissue and biostimulatory effects on tissue regeneration.101,102 Laser therapy significantly improved clinical parameters, including the plaque index, gingival index, probing depth, clinical attachment level and BOP.89 The biochemical parameters IL-1β and TNF-α were reduced as well.103
For the treatment of periodontal inflammation, Nd:YAG therapy combined with SRP was more effective than SRP alone in reducing GCF IL-1β levels at the 1-week, 3-month and 6-month follow-ups.104,105 Nd:YAG laser therapy was also effective in reducing serum IL-1β in patients with periodontal diseases.106 Er,Cr:YSGG laser therapy decreased IL-1β in both aggressive periodontitis and chronic periodontitis.107 However, SRP plus the adjunctive use of diode laser therapy did not reduce GCF IL-1β levels compared to SRP alone,108 nor did it reduce inflammation in sites with ≥5 mm PD.109 Er:YAG laser irradiation had little effect on GCF IL-1β levels, both in the SRP+laser group and in the laser-only group.110
Different types of lasers are more effective in some specific clinical areas. For example, the Nd:YAG laser could be used to remove gingival tissue and was the only laser for possible periodontal regeneration.111 The present evidence showed that the Nd:YAG laser was also effective in reducing IL-1β. However, more evidence about the types of lasers, different wavelengths and the best exposure protocol is needed to assess laser therapy.
Antimicrobial photodynamic therapy (aPDT)
aPDT is an antimicrobial treatment. The therapy is an oxygen-dependent photochemical reaction that occurs upon light-triggered activation of a certain photosensitising compound. The reaction finally leads to the production of cytotoxic reactive oxygen species, predominantly singlet oxygen, which kills microorganisms including viruses, bacteria and fungi.112 Due to its antibacterial effects, aPDT along with SRP is advantageous and effective in the treatment of periodontitis.113 aPDT plus SRP resulted in a significant reduction in probing depth and a suppression of IL-1β and MMP-8 in GCF when compared with SRP alone114 aPDT plus SRP also decreased the amount of periodontal pathogens (red and orange complexes) and lowered the IL-1β/ IL-10 ratio in GCF compared to SRP alone.115 Compared with the systemic use of metronidazole, aPDT significantly lowered the bacterial load and IL-1β level of the gingival sulcus of rats with periodontitis.116 These results suggest that aPDT has an anti-inflammatory effect on IL-1β.
Overall, conventional treatment, including SRP, surgery and antibiotics, has limited effects on IL-1β. Some new adjunctive treatments, including laser therapy and antimicrobial photodynamic therapy, are comparably more efficient than conventional treatment. Some potential adjunctive treatments might be new strategies to target IL-1β.
Therapeutic strategies for the potential use of IL-1 blockage in periodontitis
Other than the current therapies for IL-1β, are there any potential strategies to target IL-1β in periodontitis?
The pathological role of IL-1 is being discovered in a broadening list of diseases in which blocking strategies of IL-1 could be effective. The diseases include all classic autoimmune diseases and autoinflammatory diseases. The classic autoinflammatory diseases are rare, including neonatal-onset multisystem inflammatory disease, Muckle-Wells syndrome, familial cold-induced autoinflammatory syndrome, hyperimmunoglobulin D syndrome, etc. Recently, the benefit of IL-1 blockage has expanded to rare conditions, such as gout, diabetes mellitus and even myeloma. These conditions share the common feature that IL-1 is involved in their pathogenesis.117 Urate crystals promote IL-1β secretion and thus induce joint inflammation in gout.118 Type II diabetes mellitus is a chronic inflammatory disease in which β cells are continuously destroyed by IL-1119 and is improved by treatment with an IL-1 receptor antagonist.120 IL-1β produced by myeloma cells stimulatesim the secretion of IL-6 by adjacent stromal cells. The excess IL-6 in turn promotes the proliferation of the pre-myeloma cells.121
The effect of IL-1β in periodontitis is discussed above. Studies early in 1998 showed that blocking IL-1β resulted in reduced progression of periodontal bone loss and attachment loss in a non-human primate model of periodontitis,122,123 suggesting that IL-1 blockage is potentially effective in periodontitis. With the current development of IL-1 blocking agents, the strategy of blocking IL-1β may have new prospects.
Since the first blocking agent, anakinra (Kineret; Amgen) was introduced in 1993,124 a variety of agents have been produced for IL-1 blockage, including IL-1β/receptor antagonists, inflammasome inhibitors and P2X7 antagonists. These antagonists/antibodies are shown in Fig. 3.

IL-1R1, subunit of IL-1 receptor; IL-1RAcP, IL-1R accessory protein.
IL-1β/receptor antagonists/antibodies
Anakinra (Kineret)
Anakinra is a recombinant homologue of the IL-1 receptor antagonist. It competes with IL-1α and IL-1β to bind to the IL-1 receptor and thus reduces the activity. Anakinra was first approved in the US in 2001 and in Europe in 2001 for the treatment of rheumatoid arthritis. It has been proven to be effective in a variety of inflammatory diseases.124,125 In 2018, the National Health Service (England) published a Clinical Commissioning Policy for anakinra to treat periodic fevers and autoinflammatory diseases at all ages. However, anakinra has a short half-life of 4–6 h; therefore, daily subcutaneous injections were required.126
Rilonacept (Arcalyst)
Rilonacept is a recombinant IL-1 antagonist consisting of the extracellular portion of the human IL-1 receptor and the IL-1 receptor accessory protein fused with the Fc portion of human IgG1. The extracellular portion has a strong affinity for both IL-1α and IL-1β, thereby neutralising their activities and functioning as an “IL-1 trap”.127,128 It was approved by the FDA in 2008 for the treatment of cryopyrin-associated periodic syndromes (CAPS). Presently, a number of clinical trials are being developed for chronic inflammatory diseases, including type I diabetes (NCT00962026), atherosclerosis (NCT00417417), hepatitis (NCT01903798) and chronic kidney disease (NCT01663103).126 Furthermore, rilonacept has a longer half-life of 6–8 days; therefore, the interval of injections can be extended to a week.129
Canakinumab (Ilaris)
Canakinumab is a high-affinity, fully human monoclonal anti-IL-1β antibody designed to exclusively bind and neutralise human IL-1β. Canakinumab was approved by the FDA in 2009 for the treatment of CAPS. Some more clinical trials for diseases such as osteoarthritis (NCT01160822), chronic obstructive pulmonary disease (NCT00581945), type II diabetes (NCT00605475), atherosclerosis (NCT00995930) and rheumatoid arthritis (NCT00504595, NCT00424346) are undergoing. The half-life of canakinumab is as long as 26 days, which ensures that the injection can be administered bimonthly, a substantial advantage over anakinra and rilonacept.126
Inflammasome inhibitors
The NLRP3 inflammasome contributes to the maturation of IL-1β. The inflammasome inhibitors include the targets caspase-1 and NLRP3.
Caspase 1 inhibitor
Belnacasan (VX-765, Vertex Pharmaceuticals) is a selective inhibitor of caspase-1. It is an orally absorbed prodrug that is converted to VRT-043198 under the action of plasma and liver esterase.129 Currently, Belnacasan is used to suppress caspase-1 activity in inflammatory diseases. It inhibits collagen-induced arthritis and lung inflammation in mouse models.130,131 It partly decreases bone resorption in the periapical lesion of rat experimental apical periodontitis.80 In addition to its anti-inflammatory effect, VX-765 also has a protective role against cell death. It combines with a P2Y12 antagonist, cangrelor, to reduce myocardial infarct size and preserve ventricular function.132 VX-765 is well tolerated in phase II clinical trials in patients with epilepsy and133 also possesses neuroprotective activity in multiple system atrophy in a mouse model.134
NLRP3 inflammasome activation inhibitors
MCC950 and β-hydroxybutyrate are two small-molecule inhibitors of the NLRP3 inflammasome. MCC950 is a diarylsulfonylurea-containing compound that selectively inhibits the activation of the NLRP3 inflammasome. MCC950 acts specifically on the NLRP3 inflammasome but not on NLRP1, AIM2 or NLRC4 inflammasomes. MCC950 blocks ASC oligomerization and the activation of caspase-1 and IL-1β in mouse and human macrophages.135 Thus, MCC950 reduces IL-1β production and may serve as a potential target for inflammatory diseases136,137,138 It also provides protection against injury. MCC950 significantly reduces the development of atherosclerotic lesions, cardiac infarction and brain injury after intracerebral haemorrhage in mice.139,140,141
β-Hydroxybutyrate (BHB) is a ketone body that is produced as an adaptive starvation response during a negative energy balance.142 BHB inhibits the NLRP3 inflammasome in macrophages and neutrophils.143,144 Although IL-1β or IL-1 receptor antagonists have been approved by the FDA and have been shown to be effective in some clinical trials, the high costs and potential risk may limit their use. BHB, an endogenous ketone, may function as a regulatory metabolite to regulate inflammation. It has an anti-inflammatory effect on retinal damage.145 BHB prevents NLRP3 inflammasome activation in both mouse and human neutrophils and reduces urate-crystal-induced inflammation in individuals with gout.144
Compared with the currently used IL-1β or IL-1 receptor antagonists, MCC950 and BHB are less expensive to produce. Due to their specificity, MCC950 and BHB will benefit NLRP3-targeted therapies for both acute and chronic inflammatory diseases.146
P2X7 antagonists
The P2X7 receptor is an ionotropic ATP-gated cation channel. DAMPs, such as eATP, which is released from damaged cells, activate the purinergic receptor and function as a second signal for assembly of the NLRP3 inflammasome. The NLRP3 inflammasome in turn initiates and amplifies the innate immune and IL-1β-dependent pro-inflammatory responses.147,148 The P2X7 receptor antagonist has potential in the treatment of chronic pain, inflammation and cancer.149,150 The P2X7 receptor antagonists oATP and A-740003 inhibit eATP-induced IL-1β secretion in P. gingivalis-infected macrophages.38 An antagonist of the P2X7 receptor, AZ106006120, reduces neutrophil infiltration and the secretion of pro-inflammatory cytokines in a mouse model of acute lung injury.151 Several P2X7 antagonists are in clinical trials for inflammatory or pain-related diseases. Unfortunately, AstraZeneca’s AZD-9056 and Pfizer’s CE-224,535 failed in phase II clinical trials. Evotec’s EVT 401 is in phase II clinical trials. GSK’s GSK1482160 and Affectis Pharmaceutical’s AFC-5128 are currently in phase I clinical trials.149
Other potential agents that influence IL-1β in periodontitis
Some in vitro and in vivo experiments showed that some other agents have anti-inflammatory properties against IL-1β. They might be candidates for future adjuvant treatments.
Plant-derived substances
Plant-derived substances are largely utilised in immunomodulatory therapy. Some of them have anti-inflammatory effects that modulate the host response in inflammation.152 Resveratrol and curcumin are produced by several plants and belong to polyphenols. They are capable of reducing IL-1β and bone loss in animal models of experimental periodontitis.152 The anti-inflammatory properties of curcumin are similar to those of chlorhexidine-metronidazole.153 Curcumin attenuates the production of IL-1β and TNF-α stimulated by LPS in rat gingival fibroblasts in vitro.154 Piperine isolated from black and long peppers exhibits anti-inflammatory activity. It inhibits alveolar bone loss and downregulates the expression of IL-1β in rat periodontitis.155 Plumbagin is extracted from the roots of Plumbago zeylanica L. It significantly decreases the expression of TNF-α, IL-1β and IL-6 and decelerates bone destruction in rats with chronic periodontitis.156
Compared to the systemic antimicrobial agents or the chemical agent chlorhexidine gluconate, plant-derived substances partly avoid the problems of drug resistance, overdoses and a number of adverse effects.157,158 Plant-derived substances have great potential as adjuvant therapy for periodontal diseases.
Anti-inflammatory agents
Some anti-inflammatory or antioxidant agents are beneficial for reducing IL-1β. Metformin is an agent for the treatment of type II diabetes. Metformin activates AMP-activated protein kinase, which has been shown to exert significant anti-inflammatory and immunosuppressive effects.159,160 Metformin reduces the concentrations of IL-1β and bone loss in a rat model of experimental periodontitis.161 In vitro, metformin inhibits LPS-influenced IL-1β production in human gingival fibroblast cells.162
Vitamin E, a potent antioxidant, is important to the host’s antioxidant defence and immune functions.163 Vitamin E decreases the secretion of IL-1β in human gingival fibroblasts stimulated with P. gingivalis LPS. As a result, vitamin E may have anti-inflammatory effects against P. gingivalis.164
Antibodies or antagonists
There are also some antibodies or antagonists that indirectly influence IL-1β. Infliximab is a monoclonal antibody against TNF-α. It reduces the expression of IL-1β in gingiva and has significant anti-inflammatory and bone-protective effects in Wistar rats with experimental periodontitis.165 Bortezomib, a proteasome inhibitor, is used as an anticancer drug. Bortezomib interrupts the breaking-down process of the proteasome and promotes the death of cancer cells. The anticancer activity is accompanied by an anti-inflammatory effect. It has been reported that bortezomib inhibits the expression of IL-1β and prevents alveolar bone absorption in experimental periodontitis.166
Conclusion
IL-1β is an important pro-inflammatory cytokine and participates in periodontitis. As a strong stimulator of bone resorption, continuous bone loss may be induced by IL-1β. Conventional therapies, SRP, surgery and antibiotics have limited effects on IL-1β. IL-1β blockage by receptor antagonists, antibodies, inhibitors, plant-derived substances and anti-inflammatory agents is beneficial for reducing IL-1β. More investigation is necessary for IL-1β blockage to be used in periodontal treatment or as an adjunctive treatment in the future.
References
Cheng, R., Hu, T. & Bhowmick, N. A. Be resistant to apoptosis: a host factor from gingival fibroblasts. Cell Death Dis. 6, e2009 (2015).
Garlet, G. P. Destructive and protective roles of cytokines in periodontitis: a re-appraisal from host defense and tissue destruction viewpoints. J. Dent. Res. 89, 1349–1363 (2010).
Cardoso, E. M., Reis, C. & Manzanares-Cespedes, M. C. Chronic periodontitis, inflammatory cytokines, and interrelationship with other chronic diseases. Postgrad. Med. 130, 98–104 (2018).
Stashenko, P., Dewhirst, F. E., Peros, W. J., Kent, R. L. & Ago, J. M. Synergistic interactions between interleukin 1, tumor necrosis factor, and lymphotoxin in bone resorption. J. Immunol. 138, 1464–1468 (1987).
Rangbulla, V., Nirola, A., Gupta, M., Batra, P. & Gupta, M. Salivary IgA, interleukin-1beta and MMP-8 as salivary biomarkers in chronic periodontitis patients. Chin. J. Dent. Res. 20, 43–51 (2017).
Kinney, J. S. et al. Crevicular fluid biomarkers and periodontal disease progression. J. Clin. Periodontol. 41, 113–120 (2014).
Offenbacher, S. et al. Periodontal disease at the biofilm-gingival interface. J. Periodontol. 78, 1911–1925 (2007).
Sanchez, G. A., Miozza, V. A., Delgado, A. & Busch, L. Salivary IL-1beta and PGE2 as biomarkers of periodontal status, before and after periodontal treatment. J. Clin. Periodontol. 40, 1112–1117 (2013).
Engebretson, S. P., Grbic, J. T., Singer, R. & Lamster, I. B. GCF IL-1beta profiles in periodontal disease. J. Clin. Periodontol. 29, 48–53 (2002).
Kusuhara, M., Isoda, K. & Ohsuzu, F. Interleukin-1 and occlusive arterial diseases. Cardiovasc. Hematol. Agents Med. Chem. 4, 229–235 (2006).
Zhu, H., Lin, X., Zheng, P. & Chen, H. Inflammatory cytokine levels in patients with periodontitis and/or coronary heart disease. Int. J. Clin. Exp. Med. 8, 2214–2220 (2015).
Bostanci, N. et al. Expression and regulation of the NALP3 inflammasome complex in periodontal diseases. Clin. Exp. Immunol. 157, 415–422 (2009).
Isaza-Guzman, D. M., Medina-Piedrahita, V. M., Gutierrez-Henao, C. & Tobon-Arroyave, S. I. Salivary levels of NLRP3 inflammasome-related proteins as potential biomarkers of periodontal clinical status. J. Periodontol. 88, 1329–1338 (2017).
McDevitt, M. J. et al. Interleukin-1 genetic association with periodontitis in clinical practice. J. Periodontol. 71, 156–163 (2000).
Nikolopoulos, G. K., Dimou, N. L., Hamodrakas, S. J. & Bagos, P. G. Cytokine gene polymorphisms in periodontal disease: a meta-analysis of 53 studies including 4178 cases and 4590 controls. J. Clin. Periodontol. 35, 754–767 (2008).
Deng, J. S., Qin, P., Li, X. X. & Du, Y. H. Association between interleukin-1beta C (3953/4)T polymorphism and chronic periodontitis: evidence from a meta-analysis. Hum. Immunol. 74, 371–378 (2013).
Ayazi, G., Pirayesh, M. & Yari, K. Analysis of interleukin-1beta gene polymorphism and its association with generalized aggressive periodontitis disease. DNA Cell Biol. 32, 409–413 (2013).
Amirisetty, R. et al. Interleukin 1beta (+3954, −511 and −31) polymorphism in chronic periodontitis patients from North India. Acta Odontol. Scand. 73, 343–347 (2015).
Ribeiro, M. S., Pacheco, R. B., Fischer, R. G. & Macedo, J. M. Interaction of IL1B and IL1RN polymorphisms, smoking habit, gender, and ethnicity with aggressive and chronic periodontitis susceptibility. Contemp. Clin. Dent. 7, 349–356 (2016).
Kornman, K. S. et al. The interleukin-1 genotype as a severity factor in adult periodontal disease. J. Clin. Periodontol. 24, 72–77 (1997).
Isaza-Guzman, D. M., Hernandez-Viana, M., Bonilla-Leon, D. M., Hurtado-Cadavid, M. C. & Tobon-Arroyave, S. I. Determination of NLRP3 (rs4612666) and IL-1B (rs1143634) genetic polymorphisms in periodontally diseased and healthy subjects. Arch. Oral Biol. 65, 44–51 (2016).
Chen, Y. J. et al. Interleukin-1beta rs1143634 polymorphism and aggressive periodontitis susceptibility: a meta-analysis. Int. J. Clin. Exp. Med. 8, 2308–2316 (2015).
Yin, W. T., Pan, Y. P. & Lin, L. Association between IL-1alpha rs17561 and IL-1beta rs1143634 polymorphisms and periodontitis: a meta-analysis. Genet. Mol. Res. https://doi.org/10.4238/gmr.15017325 (2016).
da Silva, F. R. P. et al. Association between the rs1143634 polymorphism in interleukin-1B and chronic periodontitis: results from a meta-analysis composed by 54 case/control studies. Gene 668, 97–106 (2018).
Lavu, V. et al. Clinical relevance of cytokines gene polymorphisms and protein levels in gingival cervical fluid from chronic periodontitis patients. Iran. J. Immunol. 14, 51–58 (2017).
Wang, H. F. et al. Association between the interleukin-1beta C-511T polymorphism and periodontitis: a meta-analysis in the Chinese population. Genet. Mol. Res. 16, gmr16019315 (2017).
Zeng, X. T. et al. Meta-analysis of association between interleukin-1beta C-511T polymorphism and chronic periodontitis susceptibility. J. Periodontol. 86, 812–819 (2015).
Huang, W., He, B. Y., Shao, J., Jia, X. W. & Yuan, Y. D. Interleukin-1beta rs1143627 polymorphism with susceptibility to periodontal disease. Oncotarget 8, 31406–31414 (2017).
Liu, Y. C., Lerner, U. H. & Teng, Y. T. Cytokine responses against periodontal infection: protective and destructive roles. Periodontol 2000 52, 163–206 (2010).
Bent, R., Moll, L., Grabbe, S. & Bros, M. Interleukin-1 beta-A friend or foe in malignancies? Int. J. Mol. Sci. 19, E2155 (2018).
Kwak, A., Lee, Y., Kim, H. & Kim, S. Intracellular interleukin (IL)-1 family cytokine processing enzyme. Arch. Pharm. Res. 39, 1556–1564 (2016).
He, Y., Hara, H. & Nunez, G. Mechanism and regulation of NLRP3 inflammasome activation. Trends Biochem. Sci. 41, 1012–1021 (2016).
Lamkanfi, M. & Dixit, V. M. Mechanisms and functions of inflammasomes. Cell 157, 1013–1022 (2014).
Franchi, L., Munoz-Planillo, R. & Nunez, G. Sensing and reacting to microbes through the inflammasomes. Nat. Immunol. 13, 325–332 (2012).
Strowig, T., Henao-Mejia, J., Elinav, E. & Flavell, R. Inflammasomes in health and disease. Nature 481, 278–286 (2012).
Hajishengallis, G. et al. Differential interactions of fimbriae and lipopolysaccharide from Porphyromonas gingivalis with the toll-like receptor 2-centred pattern recognition apparatus. Cell. Microbiol. 8, 1557–1570 (2006).
Park, E. et al. Activation of NLRP3 and AIM2 inflammasomes by Porphyromonas gingivalis infection. Infect. Immun. 82, 112–123 (2014).
Ramos-Junior, E. S. et al. A dual role for P2X7 receptor during Porphyromonas gingivalis infection. J. Dent. Res. 94, 1233–1242 (2015).
Yilmaz, O. et al. ATP-dependent activation of an inflammasome in primary gingival epithelial cells infected by Porphyromonas gingivalis. Cell. Microbiol. 12, 188–198 (2010).
Pelegrin, P., Barroso-Gutierrez, C. & Surprenant, A. P2X7 receptor differentially couples to distinct release pathways for IL-1beta in mouse macrophage. J. Immunol. 180, 7147–7157 (2008).
Netea, M. G. et al. Differential requirement for the activation of the inflammasome for processing and release of IL-1beta in monocytes and macrophages. Blood 113, 2324–2335 (2009).
Bostanci, N., Meier, A., Guggenheim, B. & Belibasakis, G. N. Regulation of NLRP3 and AIM2 inflammasome gene expression levels in gingival fibroblasts by oral biofilms. Cell. Immunol. 270, 88–93 (2011).
Belibasakis, G. N., Guggenheim, B. & Bostanci, N. Downregulation of NLRP3 inflammasome in gingival fibroblasts by subgingival biofilms: involvement of Porphyromonas gingivalis. Innate Immun. 19, 3–9 (2013).
Cheng, R. et al. Porphyromonas gingivalis-derived lipopolysaccharide combines hypoxia to induce caspase-1 activation in periodontitis. Front. Cell. Infect. Microbiol. 7, 474 (2017).
Belibasakis, G. N. & Johansson, A. Aggregatibacter actinomycetemcomitans targets NLRP3 and NLRP6 inflammasome expression in human mononuclear leukocytes. Cytokine 59, 124–130 (2012).
Kelk, P., Claesson, R., Chen, C., Sjostedt, A. & Johansson, A. IL-1beta secretion induced by Aggregatibacter (Actinobacillus) actinomycetemcomitans is mainly caused by the leukotoxin. Int. J. Clin. Exp. Med. 298, 529–541 (2008).
Kelk, P., Johansson, A., Claesson, R., Hanstrom, L. & Kalfas, S. Caspase 1 involvement in human monocyte lysis induced by Actinobacillus actinomycetemcomitans leukotoxin. Infect. Immun. 71, 4448–4455 (2003).
Kelk, P. et al. Abundant secretion of bioactive interleukin-1beta by human macrophages induced by Actinobacillus actinomycetemcomitans leukotoxin. Infect. Immun. 73, 453–458 (2005).
Zhao, P., Liu, J., Pan, C. & Pan, Y. NLRP3 inflammasome is required for apoptosis of Aggregatibacter actinomycetemcomitans-infected human osteoblastic MG63 cells. Acta Histochem. 116, 1119–1124 (2014).
Kim, S., Park, M. H., Song, Y. R., Na, H. S. & Chung, J. Aggregatibacter actinomycetemcomitans-induced AIM2 inflammasome activation is suppressed by xylitol in differentiated THP-1 macrophages. J. Periodontol. 87, e116–e126 (2016).
Okinaga, T., Ariyoshi, W. & Nishihara, T. Aggregatibacter actinomycetemcomitans invasion induces interleukin-1beta production through reactive oxygen species and cathepsin B. J. Interferon Cytokine Res. 35, 431–440 (2015).
Faizuddin, M., Bharathi, S. H. & Rohini, N. V. Estimation of interleukin-1beta levels in the gingival crevicular fluid in health and in inflammatory periodontal disease. J. Periodontal Res. 38, 111–114 (2003).
Schett, G., Dayer, J. M. & Manger, B. Interleukin-1 function and role in rheumatic disease. Nat. Rev. Immunol. 12, 14–24 (2016).
Kobayashi, M. et al. Role of interleukin-1 and tumor necrosis factor alpha in matrix degradation of human osteoarthritic cartilage. Arthritis Rheum. 52, 128–135 (2005).
Makela, M., Salo, T., Uitto, V. J. & Larjava, H. Matrix metalloproteinases (MMP-2 and MMP-9) of the oral cavity: cellular origin and relationship to periodontal status. J. Dent. Res. 73, 1397–1406 (1994).
Salo, T., Makela, M., Kylmaniemi, M., Autio-Harmainen, H. & Larjava, H. Expression of matrix metalloproteinase-2 and -9 during early human wound healing. Lab. Invest. 70, 176–182 (1994).
Rai, B., Kaur, J., Jain, R. & Anand, S. C. Levels of gingival crevicular metalloproteinases-8 and -9 in periodontitis. Saudi Dent. J. 22, 129–131 (2010).
Rai, B., Kharb, S., Jain, R. & Anand, S. C. Biomarkers of periodontitis in oral fluids. J. Oral Sci. 50, 53–56 (2008).
Kusano, K. et al. Regulation of matrix metalloproteinases (MMP-2, -3, -9, and -13) by interleukin-1 and interleukin-6 in mouse calvaria: association of MMP induction with bone resorption. Endocrinology 139, 1338–1345 (1998).
Du, M. et al. Effects of IL-1beta on MMP-9 expression in cementoblast-derived cell line and MMP-mediated degradation of type I collagen. Inflammation 42, 413–425 (2019).
Murayama, R., Kobayashi, M., Takeshita, A., Yasui, T. & Yamamoto, M. MAPKs, activator protein-1 and nuclear factor-kappaB mediate production of interleukin-1beta-stimulated cytokines, prostaglandin E(2) and MMP-1 in human periodontal ligament cells. J. Periodontal Res. 46, 568–575 (2011).
Nakaya, H., Oates, T. W., Hoang, A. M., Kamoi, K. & Cochran, D. L. Effects of interleukin-1 beta on matrix metalloproteinase-3 levels in human periodontal ligament cells. J. Periodontol. 68, 517–523 (1997).
Kida, Y. et al. Interleukin-1 stimulates cytokines, prostaglandin E2 and matrix metalloproteinase-1 production via activation of MAPK/AP-1 and NF-kappaB in human gingival fibroblasts. Cytokine 29, 159–168 (2005).
Belibasakis, G. N. & Bostanci, N. The RANKL-OPG system in clinical periodontology. J. Clin. Periodontol. 39, 239–248 (2012).
Huynh, N. C., Everts, V., Pavasant, P. & Ampornaramveth, R. S. Interleukin-1beta induces human cementoblasts to support osteoclastogenesis. Int. J. Oral Sci. 9, e5 (2017).
Nakamura, I. & Jimi, E. Regulation of osteoclast differentiation and function by interleukin-1. Vitam. Horm. 74, 357–370 (2006).
Bloemen, V., Schoenmaker, T., de Vries, T. J. & Everts, V. IL-1beta favors osteoclastogenesis via supporting human periodontal ligament fibroblasts. J. Cell. Biochem. 112, 1890–1897 (2011).
Diercke, K., Kohl, A., Lux, C. J. & Erber, R. IL-1beta and compressive forces lead to a significant induction of RANKL-expression in primary human cementoblasts. J. Orofac. Orthop. 73, 397–412 (2012).
Yang, C. Y. et al. RANKL deletion in periodontal ligament and bone lining cells blocks orthodontic tooth movement. Int. J. Oral Sci. 10, 3 (2018).
Ruscitti, P. et al. The role of IL-1beta in the bone loss during rheumatic diseases. Mediat. Inflamm. 2015, 782382 (2015).
Belibasakis, G. N. et al. Gene expression of transcription factor NFATc1 in periodontal diseases. APMIS 119, 167–172 (2011).
Birkedal-Hansen, H. Role of cytokines and inflammatory mediators in tissue destruction. J. Periodontal Res. 28, 500–510 (1993).
Corrado, A., Neve, A., Maruotti, N. & Cantatore, F. P. Bone effects of biologic drugs in rheumatoid arthritis. Clin. Dev. Immunol. 2013, 945945 (2013).
Walsh, M. C. & Choi, Y. Biology of the RANKL-RANK-OPG system in immunity, bone, and beyond. Front. Immunol. 5, 511 (2014).
Sakata, M. et al. Osteoprotegerin levels increased by interleukin-1beta in human periodontal ligament cells are suppressed through prostaglandin E(2) synthesized de novo. Cytokine 18, 133–139 (2002).
Wada, N., Maeda, H., Yoshimine, Y. & Akamine, A. Lipopolysaccharide stimulates expression of osteoprotegerin and receptor activator of NF-kappa B ligand in periodontal ligament fibroblasts through the induction of interleukin-1 beta and tumor necrosis factor-alpha. Bone 35, 629–635 (2004).
Takegami, N. et al. RANK/RANKL/OPG system in the intervertebral disc. Arthritis Res. Ther. 19, 121 (2017).
Suda, T. et al. Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr. Rev. 20, 345–357 (1999).
Matsuura, T. et al. Involvement of CX3CL1 in the migration of osteoclast precursors across osteoblast layer stimulated by interleukin-1ss. J. Cell. Physiol. 232, 1739–1745 (2017).
Cheng, R. et al. The extent of pyroptosis varies in different stages of apical periodontitis. Biochim. Biophys. Acta 1864, 226–237 (2018).
Nakashyan, V., Tipton, D. A., Karydis, A., Livada, R. & Stein, S. H. Effect of 1,25(OH)2 D3 and 20(OH)D3 on interleukin-1beta-stimulated interleukin-6 and -8 production by human gingival fibroblasts. J. Periodontal Res. 52, 832–841 (2017).
Bhawal, U. K. et al. Micromolar sodium fluoride mediates anti-osteoclastogenesis in Porphyromonas gingivalis-induced alveolar bone loss. Int. J. Oral Sci. 7, 242–249 (2015).
Hienz, S. A., Paliwal, S. & Ivanovski, S. Mechanisms of bone resorption in periodontitis. J. Immunol. Res. 2015, 615486 (2015).
Slots, J. Periodontitis: facts, fallacies and the future. Periodontol 2000 75, 7–23 (2017).
Fleischer, H. C., Mellonig, J. T., Brayer, W. K., Gray, J. L. & Barnett, J. D. Scaling and root planing efficacy in multirooted teeth. J. Periodontol. 60, 402–409 (1989).
Brayer, W. K., Mellonig, J. T., Dunlap, R. M., Marinak, K. W. & Carson, R. E. Scaling and root planing effectiveness: the effect of root surface access and operator experience. J. Periodontol. 60, 67–72 (1989).
Al-Shammari, K. F. et al. Effect of non-surgical periodontal therapy on C-telopeptide pyridinoline cross-links (ICTP) and interleukin-1 levels. J. Periodontol. 72, 1045–1051 (2001).
Aljateeli, M. et al. Surgical periodontal therapy with and without initial scaling and root planing in the management of chronic periodontitis: a randomized clinical trial. J. Clin. Periodontol. 41, 693–700 (2014).
Saglam, M. et al. Combined application of Er:YAG and Nd:YAG lasers in treatment of chronic periodontitis. A split-mouth, single-blind, randomized controlled trial. J. Periodontal Res. 52, 853–862 (2017).
Tsang, Y. C., Corbet, E. F. & Jin, L. J. Subgingival glycine powder air-polishing as an additional approach to nonsurgical periodontal therapy in subjects with untreated chronic periodontitis. J. Periodontal Res. 53, 440–445 (2018).
Konopka, L., Pietrzak, A. & Brzezinska-Blaszczyk, E. Effect of scaling and root planing on interleukin-1beta, interleukin-8 and MMP-8 levels in gingival crevicular fluid from chronic periodontitis patients. J. Periodontal Res. 47, 681–688 (2012).
Kaushik, R., Yeltiwar, R. K. & Pushpanshu, K. Salivary interleukin-1beta levels in patients with chronic periodontitis before and after periodontal phase I therapy and healthy controls: a case-control study. J. Periodontol. 82, 1353–1359 (2011).
Reinhardt, R. A. et al. IL-1 in gingival crevicular fluid following closed root planing and papillary flap debridement. J. Clin. Periodontol. 20, 514–519 (1993).
Killeen, A. C., Harn, J. A., Erickson, L. M., Yu, F. & Reinhardt, R. A. Local minocycline effect on inflammation and clinical attachment during periodontal maintenance: randomized clinical trial. J. Periodontol. 87, 1149–1157 (2016).
Killeen, A. C. et al. Two-year randomized clinical trial of adjunctive minocycline microspheres in periodontal maintenance. J. Dent. Hyg. 92, 51–58 (2018).
Machtei, E. E. & Younis, M. N. The use of 2 antibiotic regimens in aggressive periodontitis: comparison of changes in clinical parameters and gingival crevicular fluid biomarkers. Quintessence Int. 39, 811–819 (2008).
Aral, K., Aral, C. A. & Kapila, Y. Six-month clinical outcomes of non-surgical periodontal treatment with antibiotics on apoptosis markers in aggressive periodontitis. Oral Dis. 25, 839–847 (2019).
Casarin, R. C. et al. The combination of amoxicillin and metronidazole improves clinical and microbiologic results of one-stage, full-mouth, ultrasonic debridement in aggressive periodontitis treatment. J. Periodontol. 83, 988–998 (2012).
Gong, Y., Lu, J., Ding, X. & Yu, Y. Effect of adjunctive roxithromycin therapy on interleukin-1beta, transforming growth factor-beta1 and vascular endothelial growth factor in gingival crevicular fluid of cyclosporine a-treated patients with gingival overgrowth. J. Periodontal Res. 49, 448–457 (2014).
Tanabe, S., Yoshioka, M., Hinode, D. & Grenier, D. Subinhibitory concentrations of tetracyclines induce lipopolysaccharide shedding by Porphyromonas gingivalis and modulate the host inflammatory response. J. Periodontal Res. 49, 603–608 (2014).
Cobb, C. M. Lasers and the treatment of periodontitis: the essence and the noise. Periodontol 2000 75, 205–295 (2017).
Ishikawa, I. et al. Application of lasers in periodontics: true innovation or myth? Periodontol 2000 50, 90–126 (2009).
Cheng, Y. et al. Efficacy of adjunctive laser in non-surgical periodontal treatment: a systematic review and meta-analysis. Lasers Med. Sci. 31, 151–163 (2016).
Qadri, T., Poddani, P., Javed, F., Tuner, J. & Gustafsson, A. A short-term evaluation of Nd:YAG laser as an adjunct to scaling and root planing in the treatment of periodontal inflammation. J. Periodontol. 81, 1161–1166 (2010).
Abduljabbar, T., Vohra, F., Kellesarian, S. V. & Javed, F. Efficacy of scaling and root planning with and without adjunct Nd:YAG laser therapy on clinical periodontal parameters and gingival crevicular fluid interleukin 1-beta and tumor necrosis factor-alpha levels among patients with periodontal disease: a prospective randomized split-mouth clinical study. J. Photochem. Photobiol. B 169, 70–74 (2017).
Javed, F. et al. Effect of Nd:YAG laser-assisted non-surgical periodontal therapy on clinical periodontal and serum biomarkers in patients with and without coronary artery disease: a short-term pilot study. Lasers Surg. Med. 48, 929–935 (2016).
Ertugrul, A. S., Tekin, Y. & Talmac, A. C. Comparing the efficiency of Er,Cr:YSGG laser and diode laser on human beta-defensin-1 and IL-1beta levels during the treatment of generalized aggressive periodontitis and chronic periodontitis. J. Cosmet. Laser Ther. 19, 409–417 (2017).
Saglam, M., Kantarci, A., Dundar, N. & Hakki, S. S. Clinical and biochemical effects of diode laser as an adjunct to nonsurgical treatment of chronic periodontitis: a randomized, controlled clinical trial. Lasers Med. Sci. 29, 37–46 (2014).
Nguyen, N. T. et al. Adjunctive non-surgical therapy of inflamed periodontal pockets during maintenance therapy using diode laser: a randomized clinical trial. J. Periodontol. 86, 1133–1140 (2015).
Lopes, B. M., Marcantonio, R. A., Thompson, G. M., Neves, L. H. & Theodoro, L. H. Short-term clinical and immunologic effects of scaling and root planing with Er:YAG laser in chronic periodontitis. J. Periodontol. 79, 1158–1167 (2008).
Ting, M., Huynh, B. H., Devine, S. M., Braid, S. M. & Suzuki, J. B. Laser treatment of periodontal disease: a systematic review of histological outcomes. EC Dent. Sci. 178, 1344–1367 (2018).
Rajesh, S., Koshi, E., Philip, K. & Mohan, A. Antimicrobial photodynamic therapy: an overview. J. Indian Soc. Periodontol. 15, 323–327 (2011).
Joseph, B., Janam, P., Narayanan, S. & Anil, S. Is antimicrobial photodynamic therapy effective as an adjunct to scaling and root planing in patients with chronic periodontitis? A systematic review. Biomolecules 7, E79 (2017).
Queiroz, A. C. et al. Adjunctive effect of antimicrobial photodynamic therapy to nonsurgical periodontal treatment in smokers: a randomized clinical trial. Lasers Med. Sci. 30, 617–625 (2015).
Moreira, A. L. et al. Antimicrobial photodynamic therapy as an adjunct to non-surgical treatment of aggressive periodontitis: a split-mouth randomized controlled trial. J. Periodontol. 86, 376–386 (2015).
Zhang, J. et al. Photodynamic therapy versus systemic antibiotic for the treatment of periodontitis in a rat model. J. Periodontol. 90, 798–807 (2019).
Dinarello, C. A. An expanding role for interleukin-1 blockade from gout to cancer. Mol. Med. 20, S43–S58 (2014).
Joosten, L. A. et al. Engagement of fatty acids with toll-like receptor 2 drives interleukin-1beta production via the ASC/caspase 1 pathway in monosodium urate monohydrate crystal-induced gouty arthritis. Arthritis Rheum. 62, 3237–3248 (2010).
Donath, M. Y. & Shoelson, S. E. Type 2 diabetes as an inflammatory disease. Nat. Rev. Immunol. 11, 98–107 (2011).
Wu, Y. Y., Xiao, E. & Graves, D. T. Diabetes mellitus related bone metabolism and periodontal disease. Int. J. Oral Sci. 7, 63–72 (2015).
Lust, J. A. & Donovan, K. A. The role of interleukin-1 beta in the pathogenesis of multiple myeloma. Hematol. Oncol. Clin. N. Am. 13, 1117–1125 (1999).
Assuma, R., Oates, T., Cochran, D., Amar, S. & Graves, D. T. IL-1 and TNF antagonists inhibit the inflammatory response and bone loss in experimental periodontitis. J. Immunol. 160, 403–409 (1998).
Graves, D. T. et al. Interleukin-1 and tumor necrosis factor antagonists inhibit the progression of inflammatory cell infiltration toward alveolar bone in experimental periodontitis. J. Periodontol. 69, 1419–1425 (1998).
Dinarello, C. A., Simon, A. & van der Meer, J. W. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat. Rev. Drug Discov. 11, 633–652 (2012).
Dinarello, C. A. & van der Meer, J. W. Treating inflammation by blocking interleukin-1 in humans. Semin. Immunol. 25, 469–484 (2013).
Arranz, L., Arriero, M. D. M. & Villatoro, A. Interleukin-1beta as emerging therapeutic target in hematological malignancies and potentially in their complications. Blood Rev. 31, 306–317 (2017).
Grom, A. A., Horne, A. & De Benedetti, F. Macrophage activation syndrome in the era of biologic therapy. Nat. Rev. Immunol. 12, 259–268 (2016).
Ilowite, N. T. et al. Randomized, double-blind, placebo-controlled trial of the efficacy and safety of rilonacept in the treatment of systemic juvenile idiopathic arthritis. Arthritis Rheumatol. 66, 2570–2579 (2014).
Fenini, G., Contassot, E. & French, L. E. Potential of IL-1, IL-18 and inflammasome inhibition for the treatment of inflammatory skin diseases. Front. Pharmacol. 8, 278 (2017).
Zhang, Y. & Zheng, Y. Effects and mechanisms of potent caspase-1 inhibitor VX765 treatment on collagen-induced arthritis in mice. Clin. Exp. Immunol. 34, 111–118 (2016).
Li, F. et al. Roles of mitochondrial ROS and NLRP3 inflammasome in multiple ozone-induced lung inflammation and emphysema. Respir. Res. 19, 230 (2018).
Audia, J. P. et al. Caspase-1 inhibition by VX-765 administered at reperfusion in P2Y12 receptor antagonist-treated rats provides long-term reduction in myocardial infarct size and preservation of ventricular function. Basic Res. Cardiol. 113, 32 (2018).
Vertex. Vertex announces completion of phase 2 study of VX-765 in people with epilepsy who did not respond to previous treatment. https://investors.vrtx.com/news-releases/news-release-details/vertex-announces-completion-phase-2-study-vx-765-people-epilepsy (2011).
Bassil, F. et al. Reducing C-terminal truncation mitigates synucleinopathy and neurodegeneration in a transgenic model of multiple system atrophy. Proc. Natl Acad. Sci. USA 113, 9593–9598 (2016).
Coll, R. C. et al. A small-molecule inhibitor of the NLRP3 inflammasome for the treatment of inflammatory diseases. Nat. Med. 21, 248–255 (2015).
Perera, A. P., Kunde, D. & Eri, R. NLRP3 inhibitors as potential therapeutic agents for treatment of inflammatory bowel disease. Curr. Pharm. Des. 23, 2321–2327 (2017).
Dolunay, A. et al. Inhibition of NLRP3 inflammasome prevents LPS-induced inflammatory hyperalgesia in mice: contribution of NF-kappaB, caspase-1/11, ASC, NOX, and NOS isoforms. Inflammation 40, 366–386 (2017).
Mridha, A. R. et al. NLRP3 inflammasome blockade reduces liver inflammation and fibrosis in experimental NASH in mice. J. Hepatol. 66, 1037–1046 (2017).
van der Heijden, T. et al. NLRP3 inflammasome inhibition by MCC950 reduces atherosclerotic lesion development in apolipoprotein E-deficient mice-brief report. Arterioscler. Thromb. Vasc. Biol. 37, 1457–1461 (2017).
van Hout, G. P. et al. The selective NLRP3-inflammasome inhibitor MCC950 reduces infarct size and preserves cardiac function in a pig model of myocardial infarction. Eur. Heart J. 38, 828–836 (2017).
Ren, H. et al. Selective NLRP3 (pyrin domain-containing protein 3) inflammasome inhibitor reduces brain injury after intracerebral hemorrhage. Stroke 49, 184–192 (2018).
Deora, V., Albornoz, E. A., Zhu, K., Woodruff, T. M. & Gordon, R. The ketone body beta-hydroxybutyrate does not inhibit synuclein mediated inflammasome activation in microglia. J. Neuroimmune Pharmacol. 12, 568–574 (2017).
Youm, Y. H. et al. The ketone metabolite beta-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med. 21, 263–269 (2015).
Goldberg, E. L. et al. β-hydroxybutyrate deactivates neutrophil NLRP3 inflammasome to relieve gout flares. Cell Rep. 18, 2077–2087 (2017).
Trotta, M. C. et al. The activation of retinal HCA2 receptors by systemic beta-hydroxybutyrate inhibits diabetic retinal damage through reduction of endoplasmic reticulum stress and the NLRP3 inflammasome. PLoS ONE 14, e0211005 (2019).
Gross, C. J. & Gross, O. The Nlrp3 inflammasome admits defeat. Trends Immunol. 36, 323–324 (2015).
Mariathasan, S. et al. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 440, 228–232 (2006).
Giuliani, A. L., Sarti, A. C., Falzoni, S. & Di Virgilio, F. The P2X7 receptor-interleukin-1 liaison. Front. Pharmacol. 8, 123 (2017).
Park, J. H. & Kim, Y. C. P2X7 receptor antagonists: a patent review (2010–2015). Expert Opin. Ther. Pat. 27, 257–267 (2017).
Burnstock, G. & Knight, G. E. The potential of P2X7 receptors as a therapeutic target, including inflammation and tumour progression. Purinergic Signal. 14, 1–18 (2018).
Mishra, A. et al. A critical role for P2X7 receptor-induced VCAM-1 shedding and neutrophil infiltration during acute lung injury. J. Immunol. 197, 2828–2837 (2016).
Correa, M. G. et al. Systemic treatment with resveratrol and/or curcumin reduces the progression of experimental periodontitis in rats. J. Periodontal Res. 52, 201–209 (2017).
Pulikkotil, S. J. & Nath, S. Effects of curcumin on crevicular levels of IL-1beta and CCL28 in experimental gingivitis. Aust. Dent. J. 60, 317–327 (2015).
Xiao, C. J., Yu, X. J., Xie, J. L., Liu, S. & Li, S. Protective effect and related mechanisms of curcumin in rat experimental periodontitis. Head Face Med. 14, 12 (2018).
Dong, Y., Huihui, Z. & Li, C. Piperine inhibit inflammation, alveolar bone loss and collagen fibers breakdown in a rat periodontitis model. J. Periodontal Res. 50, 758–765 (2015).
Zheng, X. Y. et al. Plumbagin suppresses chronic periodontitis in rats via down-regulation of TNF-alpha, IL-1beta and IL-6 expression. Acta Pharmacol. Sin. 38, 1150–1160 (2017).
Saquib, S. A. et al. Evaluation and comparison of antibacterial efficacy of herbal extracts in combination with antibiotics on periodontal pathobionts: an in vitro microbiological study. Antibiotics 8, E89 (2019).
Koychev, S., Dommisch, H., Chen, H. & Pischon, N. Antimicrobial effects of mastic extract against oral and periodontal pathogens. J. Periodontol. 88, 511–517 (2017).
Hawley, S. A., Gadalla, A. E., Olsen, G. S. & Hardie, D. G. The antidiabetic drug metformin activates the AMP-activated protein kinase cascade via an adenine nucleotide-independent mechanism. Diabetes 51, 2420–2425 (2002).
Salt, I. P. & Palmer, T. M. Exploiting the anti-inflammatory effects of AMP-activated protein kinase activation. Expert Opin. Investig. Drugs 21, 1155–1167 (2012).
Araujo, A. A. et al. Effects of metformin on inflammation, oxidative stress, and bone loss in a rat model of periodontitis. PLoS ONE 12, e0183506 (2017).
Kang, W. et al. Metformin inhibits Porphyromonas gingivalis lipopolysaccharide-influenced inflammatory response in human gingival fibroblasts via regulating activating transcription factor-3 expression. J. Periodontol. 88, e169–e178 (2017).
Puertollano, M. A., Puertollano, E., de Cienfuegos, G. A. & de Pablo, M. A. Dietary antioxidants: immunity and host defense. Curr. Top. Med. Chem. 11, 1752–1766 (2011).
Derradjia, A. et al. alpha-tocopherol decreases interleukin-1beta and -6 and increases human beta-defensin-1 and -2 secretion in human gingival fibroblasts stimulated with Porphyromonas gingivalis lipopolysaccharide. J. Periodontal Res. 51, 295–303 (2016).
Goncalves, D. C. et al. Infliximab attenuates inflammatory osteolysis in a model of periodontitis in wistar rats. Exp. Biol. Med. 239, 442–453 (2014).
Jiang, L. et al. The proteasome inhibitor bortezomib inhibits inflammatory response of periodontal ligament cells and ameliorates experimental periodontitis in rats. J. Periodontol. 88, 473–483 (2017).
Acknowledgements
This research was funded by the National Natural Science Foundation of China (Grant No. 81970948), the Innovation Spark Project of Sichuan University (2018SCUH0054) and the Sichuan Provincial Natural Science Foundation of China (2018SZ0139).
Author information
Authors and Affiliations
Contributions
T.H. and R.C. jointly designed the theme and content structure. R.C. and Z.W. wrote the manuscript. M.L. contributed to the figures. M.S. contributed to the revision. T.H. conducted the editing and critical revision.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cheng, R., Wu, Z., Li, M. et al. Interleukin-1β is a potential therapeutic target for periodontitis: a narrative review. Int J Oral Sci 12, 2 (2020). https://doi.org/10.1038/s41368-019-0068-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41368-019-0068-8
This article is cited by
-
OralExplorer: a web server for exploring the mechanisms of oral inflammatory diseases
Journal of Translational Medicine (2024)
-
New insights into nanotherapeutics for periodontitis: a triple concerto of antimicrobial activity, immunomodulation and periodontium regeneration
Journal of Nanobiotechnology (2024)
-
A Robust Strategy Against Multi-Resistant Pathogens in Oral Health: Harnessing the Potency of Antimicrobial Peptides in Nanofiber-Mediated Therapies
International Journal of Peptide Research and Therapeutics (2024)
-
MicroRNA‐155 (miR-155) as an accurate biomarker of periodontal status and coronary heart disease severity: a case–control study
BMC Oral Health (2023)
-
Macrophages immunomodulation induced by Porphyromonas gingivalis and oral antimicrobial peptides
Odontology (2023)
