
Abstract
Hair follicles, which are connected to sebaceous glands in the skin, undergo cyclic periods of regeneration, degeneration, and rest throughout adult life in mammals. The crucial function of hair follicle stem cells is to maintain these hair growth cycles. Another vital aspect is the activity of melanocyte stem cells, which differentiate into melanin-producing melanocytes, contributing to skin and hair pigmentation. Sebaceous gland stem cells also have a pivotal role in maintaining the skin barrier by regenerating mature sebocytes. These stem cells are maintained in a specialized microenvironment or niche and are regulated by internal and external signals, determining their dynamic behaviors in homeostasis and hair follicle regeneration. The activity of these stem cells is tightly controlled by various factors secreted by the niche components around the hair follicles, as well as immune-mediated damage signals, aging, metabolic status, and stress. In this study, we review these diverse stem cell regulatory and related molecular mechanisms of hair regeneration and disease conditions. Molecular insights would provide new perspectives on the disease mechanisms as well as hair and skin disorder treatment.
Similar content being viewed by others
Introduction
Hair follicles (HFs) are complex structures within the skin with crucial roles in various functions, such as thermoregulation and sensory input detection1. They comprise actively dividing progenitor cells in the hair matrix, which proliferate and develop into differentiated progeny, and hair follicle stem cells (HFSCs), which undergo dynamic molecular changes necessary for hair growth promotion and wound healing promotion in the skin2,3,4. HFSCs are regulated by internal and external signals, which determine their behaviors3,5. Transcription factors and key signaling pathways, such as the BMP, FOXC1, NFATC1, Shh, and Wnt pathways, are critical for regulating HFSC behaviors2,6,7,8,9,10. The dermal papilla (DP), comprising mesenchymal cells, regulates HF growth, and epithelial-mesenchymal interactions are pivotal for maintaining morphogenesis and the hair growth cycle11. While progress has been made in developing approaches to target the regulatory mechanisms of the DP and various HFSC niches in the promotion of HF growth12,13, further research is needed for an in-depth understanding of their potential in complete HF regeneration.
The immune privilege (IP) of anagen HFs is a unique characteristic that protects follicles from immune attacks14. Several inflammatory hair loss disorders have been linked to HF-IP dysfunction and loss15,16. Hormones, such as androgens, estrogens, and prolactin, are also central to the regulation of the hair growth cycle17,18,19. Increased androgen or prolactin levels, as well as hormonal imbalances, are closely linked to baldness, also known as alopecia20,21. Exploring the immune response regulatory mechanisms within the HF microenvironment and the hormonal effects in regulating hair growth is vital to addressing these conditions22,23,24.
Melanocytes in the skin play a significant role in determining skin and hair color as well as safeguarding against ultraviolet (UV)-induced DNA damage. Understanding the gene expression and signaling pathway regulation in melanocyte stem cells (MeSCs) in niches is thus essential for developing effective strategies to enhance skin health and prevent skin pigment disorders25. Furthermore, environmental factors, such as UV radiation, can also affect melanocyte behavior and alter gene expression, leading to pigmentation-related changes and an increased risk of skin cancer26. Sebaceous glands (SGs) are HF-associated sebum lipid-producing essential epidermal structures that contribute to barrier function and microbiome composition in the skin27. Excess and reduced sebum production result in problems such as oily skin and eczematous diseases, respectively28. Moreover, androgenic alopecia (AGA), a common hair loss condition, is characterized by SG hypertrophy29. Therefore, further research is needed to fully understand the regulatory mechanism of MeSC and SG stem cell dynamics in the hair growth cycle and identify novel targets for therapeutic interventions.
This review delves into the complex molecular and cellular processes involved in niche-related dynamic skin stem cell behaviors in HFs. Specifically, we summarized HFSC, MeSC, and SG stem cell functions and activities during hair growth cycles. We aimed to provide a comprehensive analysis of the immune-mediated damage process in HFs as well as the effects of aging, metabolic changes, and stress on HF regeneration. A detailed overview of these aspects could encourage future research and provide new insights into hair regeneration.
Stem cell dynamics in the hair follicle
The HF is a unique miniature organ in mammalian skin that undergoes continuous regeneration cycles comprising the anagen, catagen, and telogen phases1,30. The anagen phase in the human scalp frequently lasts for 2–6 years, whereas in mouse dorsal skin, it normally lasts for approximately 2 weeks31. Due to its characteristics, the mouse HF is a useful model for studying stem cell quiescence and activation32,33. During the telogen phase, a quiescent HFSC population resides in the outer layer of the bulge region, an anatomical niche, while primed HFSCs are located in the hair germ2,3. HFSCs remain quiescent for most of the hair growth cycle, except for the early anagen phase when they become active. During the telogen-to-anagen transition, primed HFSCs in the hair germ are the first cells to become activated before bulge HFSC activation3. The anagen phase begins when the quiescent signals from the inner bulge layer and other HFSC niches are overwhelmed by a threshold level of the combined BMP inhibitory and Wnt activating signals3,6,34,35,36. These primed HFSCs generate transit-amplifying cells (TACs), rapidly proliferating progenitor cells located in the hair matrix2. The DP regulates the lineage choices of TACs throughout the progression of the anagen phase37. Following terminal differentiation, TACs give rise to downstream HF lineages, including the hair shaft, companion layer, and inner root sheath38. On the other hand, bulge HFSCs mainly give rise to the downward-growing outer root sheath34. During the catagen phase, the lower portion of the HF regresses with a gradually shortening epithelial strand before entering the upcoming telogen phase39.
MeSCs are undifferentiated cells that originate from the neural crest and reside in the bulge and hair germ area of the HF40. During the telogen phase, MeSCs remain quiescent until anagen onset. In the early anagen phase, activated MeSCs generate proliferating progenitors that migrate to the hair bulb and differentiate into melanin-producing melanocytes, which are responsible for the color of the newly formed HFs41. During the catagen phase, differentiated melanocytes undergo apoptosis within the hair matrix42. However, when exposed to UV radiation, MeSCs can migrate toward the epidermis and differentiate into melanocytes, contributing to epidermal pigmentation as a protective function43.
The SG maintains its connection to the HF structure, located in the upper permanent HF region, specifically in the junctional zone where Lrig1+ SG stem cells reside44. SCD1+ proliferative SG progenitor cells are continuously produced within the SG outer layer. These cells subsequently differentiate into mature sebocytes filled with lipids in the inner layer45. Mature sebocytes undergo a unique form of programmed cell death during the holocrine secretion of lipid-containing sebum, in which their cellular organelles are degraded46. Sebum is released through the sebaceous secretory duct of the SG into the HF canal, ultimately reaching the skin surface28,47. However, disrupted SG homeostasis, such as that seen in sebaceous hyperplasia, could affect HF–SG communication48.
Stem cell regulatory mechanisms in the hair follicle niche
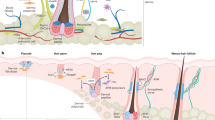
HFSCs are sustained by niches, specialized microenvironments that regulate their functions13,49,50. The niches are composed of neighboring cells that produce various factors, impacting HFSCs both directly and indirectly36,51,52. One of the most well-known HFSC niches is the DP, a mesenchymal cell cluster that modulates HFSC activation, proliferation, and differentiation through multiple signals32. Other niches include those of adipocyte precursor cells, mature dermal adipocytes, dermal macrophages, regulatory T cells, blood vessels, lymphatic vessels, and sympathetic nerves53,54,55 (Fig. 1). These niches secrete signals that either activate HFSCs or maintain their quiescence during the hair growth cycle12.

Hair follicle stem cells can generate various cell types within the HF. The immune system, including T cells, macrophages, dendritic cells, and Treg cells, undergoes fluctuations during the hair growth cycle and plays an important role in HF regeneration. Blood vessels play a crucial role in HF regeneration by providing oxygen, nutrients, and growth factors essential for hair growth and maintenance. The surrounding vasculature enables the delivery of these vital components. The interactions among lymphatic vessels, sympathetic nerves, sensory nerves, and HFs are complex and interconnected, collectively contributing to HF function regulation, including nutrient supply, immune surveillance, and responses to environmental cues.
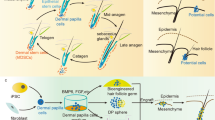
Several signaling pathways, such as the BMP, FGF, Wnt, and JAK/STAT pathways, mediate niche factor effects on HFSC activity. BMP signaling maintains HFSC quiescence during early telogen, and premature HFSC activation occurs upon BMP receptor 1A ablation56,57. In addition, upregulation of DP-derived BMP antagonists, such as Noggin, Bambi, or Sostdc1, promotes the early-to-late telogen transition in adult HFs3,6. Additionally, FGF7 and FGF10 upregulation within the DP in late telogen stimulates HFSC activation in the hair germ located above the DP3. FGF18 is highly expressed throughout the telogen phase and secreted from the inner bulge layer, regulating the HFSC quiescent state34,58. FGF9 from dermal γδ T cells is important in the wound-induced HF neogenesis process59. Pharmacological inhibition of the JAK-STAT pathway enhances hair growth by disrupting HFSC quiescence60. Dermal TREM2+ macrophage-produced oncostatin M regulates HFSC quiescence by acting upstream of JAK-STAT5 signaling61. Wnt/β-catenin signaling promotes HFSC activation and HF growth10,62. However, DP cell stimulation with dihydrotestosterone prevents HFSC differentiation by increasing the expression of the Wnt inhibitor Dkk-163.
The identity of HFSCs and the capacity for HF regeneration are governed by cell-intrinsic molecular factors, such as transcription factors and epigenetic modifiers that control gene expression (Table 1). These cell-intrinsic molecules may function as downstream or upstream signaling pathway targets to coordinate the effects of niche factors on HFSC activity as well as HF growth5,36. Several transcription factors have been found to influence HFSC quiescence, self-renewal, directional migration, or lineage fate specification during the hair growth cycle7,8,9,64,65,66,67,68,69,70,71,72,73,74,75,76. Additionally, chromatin remodeling factors can affect HFSC behaviors. The histone modification activity of polycomb repressive complex 1 (PRC1) is necessary to maintain LGR5+ HFSCs and HF regeneration77. Loss of DNA methyltransferase 1 (DNMT 1) results in delayed HFSC activation after plucking and reduced proliferation of HF progenitor cells78. Further study is needed to elucidate the intricate molecular mechanisms by which transcription factors and epigenetic modulators interact with HFSC niches, thus regulating the expression of genes specific to the hair growth cycle.
MeSCs reside in the same niches as HFSCs and undergo periodic rest, activation, and degeneration phases, coinciding with HFSCs, for hair pigmentation79. The quiescent MeSC state depends on the role of niche-derived TGF-β signaling80. Adjacent HFSCs in the hair germ produce Wnt ligands, including Wnt10b, with the potential to activate MeSCs. Moreover, MeSC activation during early anagen relies on the significance of Wnt/β-catenin signaling79. Interestingly, neighboring HFSCs secrete endothelin-1 (Edn1) during anagen onset, and HFSCs exhibit increased Edn2 expression upon the removal of the NFIB transcription factor79,81. The Edn/EdnrB pathway affects MeSC proliferation and maintenance82. The ligand for c-KIT, stem cell factor (SCF) secreted from the hair matrix and DP is critical for melanocyte maintenance83,84. When exposed to UV irradiation, MeSCs from HFs migrate to the epidermal layer through melanocortin 1 receptor signaling and differentiate into functional melanocytes to protect damaged skin43.
SG regulation is a complex process encompassing both local mediators and potential long-range hormonal signals85. Multiple signaling pathways, including the Notch, Wnt, FGF, retinoid and aryl hydrocarbon receptor pathways, mediate the effects on SG stem cell activity. However, our understanding of the SG regulatory niche remains limited. Notch signaling directly promotes SG stem cell differentiation and indirectly inhibits these glands outside the stem cell population85. FGF signaling influences SG homeostasis, as proven by SG atrophy in the tail skin of mutant mice lacking Fgfr2b86. Retinoids can treat acne by shrinking SGs and reducing sebum production87. The immune system-mediated regulation of sebum secretion is noteworthy. Thymic stromal lymphopoietin (TSLP), an epithelial cell-produced cytokine, stimulates the migration of T cells to SGs, which is necessary for increased sebum secretion88. The absence of skin-resident innate lymphoid cells (ILCs) leads to SG hyperplasia due to the lack of ILC-derived TNF and lymphotoxin production, mediating the antiproliferative effect on sebocytes by Notch signaling inhibition89.
In conclusion, dysfunctional HF niches negatively impact the hair growth cycle, pigmentation, or sebum production by influencing stem cell functions in HFs. Therefore, understanding the stem cell regulation-related signaling pathways and niche factors in HFs is important to developing strategies for HF regeneration and the treatment of various hair and skin disorders.
Immune privilege in hair follicle growth and regeneration
HF-IP is critical for maintaining the hair growth cycle and preventing immune-mediated HF damage90,91. This protective mechanism remains active during the anagen phase and restricts immune surveillance14. Alopecia areata (AA) is a form of nonscarring hair loss in humans characterized by HF-IP breakdown and lymphocyte infiltration around the anagen bulb16. In AA, HFs undergo dystrophy and transition into the catagen and telogen phases, resulting in a reduced proportion of anagen HFs16. MHC class I and II molecules (MHC-I and MHC-II, respectively), which are involved in immune cell-related foreign antigen recognition, are reduced in the bulge and hair bulb during HF-IP92,93,94,95,96. HF-IP effectively suppresses the immune cell response by establishing an immunosuppressive signaling environment in anagen HFs, facilitated by significant CD200 expression in the bulge97,98. In addition, other mediators produced by HFs, including neuropeptides (α-MSH), growth factor family members (TGF-β1/2, IGF-1), and macrophage migration inhibitory factor, contribute to CD8+ T-cell and natural killer cell suppression99,100,101,102,103. These mediators work together to maintain a stable hair growth cycle by preventing the immune system from attacking HFs14,97 (Fig. 2).

Immune cell populations can be found around HFs in a typical hair growth cycle. However, when alopecia areata develops, the anagen phase is significantly reduced, resulting in disordered HF growth. This is accompanied by a characteristic inflammatory cell infiltration in the HF-surrounding area, involving CD4+ and CD8+ T cells, mast cells, NK cells, and dendritic cells. Typically, CD8+ T cells are the first to infiltrate the intrafollicle, followed by dendritic cells and macrophages. CD4+ T cells are rarely found within HFs until late in the disease process. The collapse of HF-IP plays a central role in AA pathogenesis, involving factors such as MHC molecules, cytokines, costimulatory molecules, Tregs, and perifollicular cells that contribute to IP maintenance, disruption, and restoration.
Recent research using “just EGFP death-inducing” (JEDI) mouse models revealed an interesting relationship between stem cell quiescence and immune system evasion ability. JEDI T cells effectively eliminated GFP+ intestinal, mammary gland, and ovarian stem cells. By reducing MHC-I and NLRC5 expression, Lgr5+ quiescent HFSCs in the bulge region exhibited complete resistance to T-cell-mediated attack104. Primary cicatricial alopecia (PCA), including frontal fibrosing alopecia and lichen planopilaris (LPP), is a scarring form of hair loss in humans, distinguished by the presence of a perifollicular lymphocytic infiltrate and inflamed HF replacement with fibrous tissue105. Lesional LPP skin can be characterized by the loss of IP in bulge HFSCs, increased MHC-I and MHC-II expression and reduced CD200 and TGFβ2 expression23,96. According to recent research, nuclear factor IB (NFIB) and IX (NFIX) are essential for maintaining the identity of HFSCs, and their removal causes immune infiltration phenotypes in Nfib/Nfix double conditional knockout mice, which resemble PCA76. Consequently, understanding HF-IP-related mechanisms is essential for developing effective treatments against hair loss disorders.
The impact of aging, metabolism, and stress on hair growth and regeneration
HF growth and regeneration are complex processes influenced by various factors, including aging and metabolism, which could impact HFSC quiescence and activation as well as hair shaft miniaturization106,107. In androgenic alopecia, HFSC loss-associated progressive HF miniaturization can be observed107,108,109. Regenerative HF domains reportedly become asynchronous with aging110, and HFSCs become increasingly quiescent7,111,112, leaving their HF niche108 or differentiating into epithelial keratinocytes during aging107 (Fig. 3). Importantly, not only aged HFSCs but also their niches may contribute to these changes113,114. Therefore, understanding the complex interactions between HFSCs and their niches during aging and metabolic disorders is essential for developing effective hair regeneration promotion and hair loss treatment strategies.

During aging, hair follicle stem cells transition into a quiescent phase, and the niches undergo age-related changes. In aging skin, the regenerative ability of HFSCs becomes impaired. The niche is vital in various aging-related processes, including epidermal differentiation and HF miniaturization. HF aging is characterized by the miniaturization and absence of the dermal papilla. Prior to miniaturization, stem cells undergo dysregulation, including the loss of HFSC markers and maintenance markers. HFSCs undergo epidermal terminal differentiation and escape from the bulge during aging.
Recent research investigating the role of the skin niche during aging has yielded valuable insights into the promotion of HF regeneration. The substantial changes in the HFSC niches leading to altered HFSC behavior and declining skin regeneration ability have been elucidated113. The study revealed that aged HFSCs could regenerate HFs in the presence of young dermis113,114, while young HFSCs could only regenerate HFs when supported by young dermis113. These results indicate that niche aging is key to determining stem cell behavior. Moreover, age-related changes were observed in various nonepithelial niche cell types, including immune cells, dermal fibroblasts, and sensory neurons113. In addition, increased cellular identity noise and a gain of proadipogenic characteristics in dermal fibroblasts could be potential underlying mechanisms of aging115. With aging, the hemidesmosomal collagen COL17A1, a basement membrane anchor for HFSCs, is depleted, thereby contributing to HF miniaturization and thinning107. Furthermore, extracellular matrix (ECM) component expression-related changes could increase basement membrane thickness and stiffness, resulting in the suppression of important HFSC activation promoters in aged HFs114.
In addition to skin niche alternations, systemic and cellular metabolic changes could also impact hair growth and regeneration. High-fat diet-induced obesity reportedly accelerates HF thinning and loss by suppressing Shh signaling and inducing HFSC differentiation into epithelial keratinocytes116. The importance of several metabolic pathways in HFSC activation has also been revealed (e.g., lactate production is potentially essential for HFSC activation)117. In addition, glutamate signaling is significant in HFSC activation, as the glutamate transporter Slc1a3 is critical for anagen entry118. Ceramide synthase 4 inactivation reportedly caused HFSC activation, suggesting that epidermal ceramide composition is significant in regulating HFSC behaviors119. Furthermore, inhibiting glutamine metabolism is necessary for progenitor cells to return to the quiescent HFSC state120.
Hair graying is among the most obvious aging phenomena in mammals. The MeSC number reportedly decreases, and MeSCs undergo premature differentiation in aged HFs121,122. In addition, ionizing radiation (IR)-induced genotoxic stress induces premature hair graying in mice. The ATM-mediated DNA damage response plays a critical role in determining MeSC fate, preventing premature differentiation122. Recently, organ-cultured human HFs have shown that exposure to IR results in pigmentation loss, establishing accumulated DNA damage as an important contributing factor in human hair graying123. Under resiniferatoxin-induced acute stress in mice, the sympathetic nervous system becomes hyperactivated, resulting in significant norepinephrine release, inducing the entry of MeSCs to a rapid proliferation state. These MeSCs migrate, differentiate, and eventually deplete from the bulge. The β2 adrenergic receptor on MeSCs is a critical molecule in stress-induced hair-graying mice124. However, whether MeSC loss during the aging process is influenced by DNA damage or the sympathetic nervous system remains unclear and needs further elucidation.
Chronic stress is reportedly a contributing factor to hair loss. Stress-induced changes in cellular signaling pathways and corticosterone inhibitory effects on hair regrowth have also been observed112. Chronic stress reportedly inhibits HFSC activation by reducing Gas6 expression. Restoring Gas6 levels reversed the effects of chronic stress and promoted hair growth in mice. Suppressing corticosterone (the mouse equivalent of cortisol) signaling could activate HFSCs and alleviate the negative impact of stress on hair growth.
Perspectives
In this study, we reviewed our current understanding of various skin stem cell-related regulatory aspects within their niches to achieve hair regeneration. Since these stem cells or niches are affected by factors such as immune dysfunction, aging, metabolic changes, and stress, finding the right target for each condition is important for overcoming hair loss, which causes psychological distress in many people.
Several hair loss treatments have already been developed. Among them, noninvasive treatments such as low-level laser therapy and microneedling therapy reportedly stimulate partial hair growth125,126. Low-level laser therapy works by stimulating HF activity and promoting hair growth through photobiomodulation, a process that involves light energy absorption by the cells125,127. Microneedling therapy is another novel treatment involving the creation of small puncture wounds on the scalp with a tiny needle-bearing device, thereby stimulating hair growth by increasing blood flow to the scalp and promoting growth factor release126. However, the underlying stem cell- and niche-specific regulatory mechanisms of these methods remain unknown.
In addition to noninvasive treatments, several hair loss treatment drugs are also available. Finasteride is an FDA-approved drug that inhibits 5-alpha reductase, an enzyme responsible for testosterone-to-dihydrotestosterone conversion, and is effective in reducing hair loss128. By reducing dihydrotestosterone production, this medication could help slow or reverse hair loss in individuals with male-pattern baldness129. JAK inhibitors have also gained attention as a potential treatment for AA, an autoimmune disorder130,131,132. These medications work by blocking the activity of JAK enzymes, which are involved in immune system function and inflammation.
Taken together, the existence of these treatments provides hope for individuals experiencing hair loss. However, the efficacy of these treatments still differs depending on the underlying cause of hair loss and personal factors. However, further understanding of the skin stem cell and niche regulation processes that affect hair growth and reverse alopecia is needed for developing improved treatment approaches.
References
Schneider, M. R., Schmidt-Ullrich, R. & Paus, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 19, R132–R142 (2009).
Hsu, Y. C., Li, L. & Fuchs, E. Transit-amplifying cells orchestrate stem cell activity and tissue regeneration. Cell 157, 935–949 (2014).
Greco, V. et al. A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell 4, 155–169 (2009).
Ito, M. et al. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat. Med. 11, 1351–1354 (2005).
Lee, S. A., Li, K. N. & Tumbar, T. Stem cell-intrinsic mechanisms regulating adult hair follicle homeostasis. Exp. Dermatol. 30, 430–447 (2021).
Plikus, M. V. et al. Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration. Nature 451, 340–344 (2008).
Lay, K., Kume, T. & Fuchs, E. FOXC1 maintains the hair follicle stem cell niche and governs stem cell quiescence to preserve long-term tissue-regenerating potential. Proc. Natl Acad. Sci. USA 113, E1506–E1515 (2016).
Wang, L., Siegenthaler, J. A., Dowell, R. D. & Yi, R. Foxc1 reinforces quiescence in self-renewing hair follicle stem cells. Science 351, 613–617 (2016).
Horsley, V., Aliprantis, A. O., Polak, L., Glimcher, L. H. & Fuchs, E. NFATc1 balances quiescence and proliferation of skin stem cells. Cell 132, 299–310 (2008).
Choi, Y. S. et al. Distinct functions for Wnt/beta-catenin in hair follicle stem cell proliferation and survival and interfollicular epidermal homeostasis. Cell Stem Cell 13, 720–733 (2013).
Sennett, R. & Rendl, M. Mesenchymal-epithelial interactions during hair follicle morphogenesis and cycling. Semin. Cell Dev. Biol. 23, 917–927 (2012).
Chen, C. L., Huang, W. Y., Wang, E. H. C., Tai, K. Y. & Lin, S. J. Functional complexity of hair follicle stem cell niche and therapeutic targeting of niche dysfunction for hair regeneration. J. Biomed. Sci. 27, 43 (2020).
Jang, H., Jo, Y., Lee, J. H. & Choi, S. Aging of hair follicle stem cells and their niches. BMB Rep. 56, 2–9 (2023).
Paus, R., Nickoloff, B. J. & Ito, T. A. ‘hairy’ privilege. Trends Immunol. 26, 32–40 (2005).
Bertolini, M., McElwee, K., Gilhar, A., Bulfone-Paus, S. & Paus, R. Hair follicle immune privilege and its collapse in alopecia areata. Exp. Dermatol 29, 703–725 (2020).
Pratt, C. H., King, L. E. Jr., Messenger, A. G., Christiano, A. M. & Sundberg, J. P. Alopecia areata. Nat. Rev. Dis. Prim. 3, 17011 (2017).
Inui, S. & Itami, S. Androgen actions on the human hair follicle: perspectives. Exp. Dermatol 22, 168–171 (2013).
Ohnemus, U. et al. Hair cycle control by estrogens: catagen induction via estrogen receptor (ER)-alpha is checked by ER beta signaling. Endocrinology 146, 1214–1225 (2005).
Craven, A. J. et al. Prolactin signaling influences the timing mechanism of the hair follicle: analysis of hair growth cycles in prolactin receptor knockout mice. Endocrinology 142, 2533–2539 (2001).
Redler, S., Messenger, A. G. & Betz, R. C. Genetics and other factors in the aetiology of female pattern hair loss. Exp. Dermatol 26, 510–517 (2017).
Lutz, G. Hair loss and hyperprolactinemia in women. Dermatoendocrinol 4, 65–71 (2012).
Paus, R., Bulfone-Paus, S. & Bertolini, M. Hair follicle immune privilege revisited: the key to alopecia areata management. J. Investig. Dermatol Symp. Proc. 19, S12–S17 (2018).
Anzai, A., Wang, E. H. C., Lee, E. Y., Aoki, V. & Christiano, A. M. Pathomechanisms of immune-mediated alopecia. Int Immunol. 31, 439–447 (2019).
Randall, V. A. Hormonal regulation of hair follicles exhibits a biological paradox. Semin. Cell Dev. Biol. 18, 274–285 (2007).
Costin, G. E. & Hearing, V. J. Human skin pigmentation: melanocytes modulate skin color in response to stress. FASEB J. 21, 976–994 (2007).
Lin, J. Y. & Fisher, D. E. Melanocyte biology and skin pigmentation. Nature 445, 843–850 (2007).
Dreno, B. et al. Microbiome in healthy skin, update for dermatologists. J. Eur. Acad. Dermatol Venereol. 30, 2038–2047 (2016).
Geueke, A. & Niemann, C. Stem and progenitor cells in sebaceous gland development, homeostasis and pathologies. Exp. Dermatol 30, 588–597 (2021).
Torkamani, N., Rufaut, N. W., Jones, L. & Sinclair, R. Destruction of the arrector pili muscle and fat infiltration in androgenic alopecia. Br. J. Dermatol 170, 1291–1298 (2014).
Muller-Rover, S. et al. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Invest Dermatol 117, 3–15 (2001).
Augustyniak, A. & Mc Mahon, H. Dietary marine-derived ingredients for stimulating hair cell cycle. Biomed. Pharmacother. 163, 114838 (2023).
Rezza, A., Sennett, R. & Rendl, M. Adult stem cell niches: cellular and molecular components. Curr. Top. Dev. Biol. 107, 333–372 (2014).
Yi, R. Concise review: mechanisms of quiescent hair follicle stem cell regulation. Stem Cells 35, 2323–2330 (2017).
Hsu, Y. C., Pasolli, H. A. & Fuchs, E. Dynamics between stem cells, niche, and progeny in the hair follicle. Cell 144, 92–105 (2011).
Hagner, A. et al. Transcriptional profiling of the adult hair follicle mesenchyme reveals r-spondin as a novel regulator of dermal progenitor function. iScience 23, 101019 (2020).
Fuchs, E. & Blau, H. M. Tissue stem cells: architects of their niches. Cell Stem Cell 27, 532–556 (2020).
Yang, H., Adam, R. C., Ge, Y., Hua, Z. L. & Fuchs, E. Epithelial-mesenchymal micro-niches govern stem cell lineage choices. Cell 169, 483–496.e413 (2017).
Rompolas, P., Mesa, K. R. & Greco, V. Spatial organization within a niche as a determinant of stem-cell fate. Nature 502, 513–518 (2013).
Heitman, N. et al. Dermal sheath contraction powers stem cell niche relocation during hair cycle regression. Science 367, 161–166 (2020).
Nishimura, E. K. et al. Dominant role of the niche in melanocyte stem-cell fate determination. Nature 416, 854–860 (2002).
Nishimura, E. K. Melanocyte stem cells: a melanocyte reservoir in hair follicles for hair and skin pigmentation. Pigment Cell Melanoma Res. 24, 401–410 (2011).
Qiu, W., Chuong, C. M. & Lei, M. Regulation of melanocyte stem cells in the pigmentation of skin and its appendages: Biological patterning and therapeutic potentials. Exp. Dermatol 28, 395–405 (2019).
Chou, W. C. et al. Direct migration of follicular melanocyte stem cells to the epidermis after wounding or UVB irradiation is dependent on Mc1r signaling. Nat. Med. 19, 924–929 (2013).
Niemann, C. & Horsley, V. Development and homeostasis of the sebaceous gland. Semin. Cell Dev. Biol. 23, 928–936 (2012).
Frances, D. & Niemann, C. Stem cell dynamics in sebaceous gland morphogenesis in mouse skin. Dev. Biol. 363, 138–146 (2012).
Fischer, H. et al. Holocrine secretion of sebum is a unique dnase2-dependent mode of programmed cell death. J. Invest. Dermatol. 137, 587–594 (2017).
Schneider, M. R. & Paus, R. Sebocytes, multifaceted epithelial cells: lipid production and holocrine secretion. Int. J. Biochem. Cell Biol. 42, 181–185 (2010).
Zouboulis, C. C. et al. Beyond acne: current aspects of sebaceous gland biology and function. Rev. Endocr. Metab. Disord. 17, 319–334 (2016).
Moore, K. A. & Lemischka, I. R. Stem cells and their niches. Science 311, 1880–1885 (2006).
Li, K. N. & Tumbar, T. Hair follicle stem cells as a skin-organizing signaling center during adult homeostasis. EMBO J. 40, e107135 (2021).
Hsu, Y. C., Li, L. & Fuchs, E. Emerging interactions between skin stem cells and their niches. Nat. Med. 20, 847–856 (2014).
Brunet, A., Goodell, M. A. & Rando, T. A. Ageing and rejuvenation of tissue stem cells and their niches. Nat. Rev. Mol. Cell Biol. 24, 45–62 (2023).
Hsu, Y. C. & Fuchs, E. Building and maintaining the skin. Cold Spring Harb. Perspect. Biol. 14, a040840 (2022).
Castellana, D., Paus, R. & Perez-Moreno, M. Macrophages contribute to the cyclic activation of adult hair follicle stem cells. PLoS Biol. 12, e1002002 (2014).
Gur-Cohen, S. et al. Stem cell-driven lymphatic remodeling coordinates tissue regeneration. Science 366, 1218–1225 (2019).
Genander, M. et al. BMP signaling and its pSMAD1/5 target genes differentially regulate hair follicle stem cell lineages. Cell Stem Cell 15, 619–633 (2014).
Kobielak, K., Stokes, N., de la Cruz, J., Polak, L. & Fuchs, E. Loss of a quiescent niche but not follicle stem cells in the absence of bone morphogenetic protein signaling. Proc. Natl Acad. Sci. USA 104, 10063–10068 (2007).
Kimura-Ueki, M. et al. Hair cycle resting phase is regulated by cyclic epithelial FGF18 signaling. J. Invest. Dermatol. 132, 1338–1345 (2012).
Gay, D. et al. Fgf9 from dermal gammadelta T cells induces hair follicle neogenesis after wounding. Nat. Med. 19, 916–923 (2013).
Harel, S. et al. Pharmacologic inhibition of JAK-STAT signaling promotes hair growth. Sci. Adv. 1, e1500973 (2015).
Wang, E. C. E., Dai, Z., Ferrante, A. W., Drake, C. G. & Christiano, A. M. A subset of TREM2(+) dermal macrophages secretes oncostatin M to maintain hair follicle stem cell quiescence and inhibit hair growth. Cell Stem Cell 24, 654–669 e656 (2019).
Myung, P. S., Takeo, M., Ito, M. & Atit, R. P. Epithelial Wnt ligand secretion is required for adult hair follicle growth and regeneration. J. Invest. Dermatol. 133, 31–41 (2013).
Leiros, G. J., Ceruti, J. M., Castellanos, M. L., Kusinsky, A. G. & Balana, M. E. Androgens modify Wnt agonists/antagonists expression balance in dermal papilla cells preventing hair follicle stem cell differentiation in androgenetic alopecia. Mol. Cell. Endocrinol. 439, 26–34 (2017).
Lien, W. H. et al. In vivo transcriptional governance of hair follicle stem cells by canonical Wnt regulators. Nat. Cell Biol. 16, 179–190 (2014).
Folgueras, A. R. et al. Architectural niche organization by LHX2 is linked to hair follicle stem cell function. Cell Stem Cell 13, 314–327 (2013).
Leishman, E. et al. Foxp1 maintains hair follicle stem cell quiescence through regulation of Fgf18. Development 140, 3809–3818 (2013).
Osorio, K. M. et al. Runx1 modulates developmental, but not injury-driven, hair follicle stem cell activation. Development 135, 1059–1068 (2008).
Chen, J. K. et al. IRX5 promotes DNA damage repair and activation of hair follicle stem cells. Stem Cell Rep. 18, 1227–1243 (2023).
Haensel, D. et al. An Ovol2-Zeb1 transcriptional circuit regulates epithelial directional migration and proliferation. EMBO Rep. 20, e46273 (2019).
Shirokova, V. et al. Foxi3 deficiency compromises hair follicle stem cell specification and activation. Stem Cells 34, 1896–1908 (2016).
Adam, R. C. et al. Pioneer factors govern super-enhancer dynamics in stem cell plasticity and lineage choice. Nature 521, 366–370 (2015).
Ren, X. et al. Lgr4 deletion delays the hair cycle and inhibits the activation of hair follicle stem cells. J. Invest. Dermatol. 140, 1706–1712 e1704 (2020).
Jaks, V. et al. Lgr5 marks cycling, yet long-lived, hair follicle stem cells. Nat. Genet. 40, 1291–1299 (2008).
Merrill, B. J., Gat, U., DasGupta, R. & Fuchs, E. Tcf3 and Lef1 regulate lineage differentiation of multipotent stem cells in skin. Genes Dev. 15, 1688–1705 (2001).
Wang, A. B., Zhang, Y. V. & Tumbar, T. Gata6 promotes hair follicle progenitor cell renewal by genome maintenance during proliferation. EMBO J. 36, 61–78 (2017).
Adam, R. C. et al. NFI transcription factors provide chromatin access to maintain stem cell identity while preventing unintended lineage fate choices. Nat. Cell Biol. 22, 640–650 (2020).
Pivetti, S. et al. Loss of PRC1 activity in different stem cell compartments activates a common transcriptional program with cell type-dependent outcomes. Sci. Adv. 5, eaav1594 (2019).
Li, J. et al. Progressive alopecia reveals decreasing stem cell activation probability during aging of mice with epidermal deletion of DNA methyltransferase 1. J. Invest. Dermatol. 132, 2681–2690 (2012).
Rabbani, P. et al. Coordinated activation of Wnt in epithelial and melanocyte stem cells initiates pigmented hair regeneration. Cell 145, 941–955 (2011).
Nishimura, E. K. et al. Key roles for transforming growth factor beta in melanocyte stem cell maintenance. Cell Stem Cell 6, 130–140 (2010).
Chang, C. Y. et al. NFIB is a governor of epithelial-melanocyte stem cell behaviour in a shared niche. Nature 495, 98–102 (2013).
Takeo, M. et al. EdnrB governs regenerative response of melanocyte stem cells by crosstalk with Wnt signaling. Cell Rep. 15, 1291–1302 (2016).
Botchkareva, N. V., Khlgatian, M., Longley, B. J., Botchkarev, V. A. & Gilchrest, B. A. SCF/c-kit signaling is required for cyclic regeneration of the hair pigmentation unit. FASEB J. 15, 645–658 (2001).
Liao, C. P., Booker, R. C., Morrison, S. J. & Le, L. Q. Identification of hair shaft progenitors that create a niche for hair pigmentation. Genes Dev. 31, 744–756 (2017).
Veniaminova, N. A. et al. Niche-specific factors dynamically regulate sebaceous gland stem cells in the skin. Dev. Cell 51, 326–340 e324 (2019).
Grose, R. et al. The role of fibroblast growth factor receptor 2b in skin homeostasis and cancer development. EMBO J. 26, 1268–1278 (2007).
Clayton, R. W. et al. Homeostasis of the sebaceous gland and mechanisms of acne pathogenesis. Br. J. Dermatol. 181, 677–690 (2019).
Choa, R. et al. Thymic stromal lymphopoietin induces adipose loss through sebum hypersecretion. Science 373, eabd2893 (2021).
Kobayashi, T. et al. Homeostatic control of sebaceous glands by innate lymphoid cells regulates commensal bacteria equilibrium. Cell 176, 982–997.e916 (2019).
Paus, R., Ito, N., Takigawa, M. & Ito, T. The hair follicle and immune privilege. J. Investig. Dermatol Symp. Proc. 8, 188–194 (2003).
Rajabi, F., Drake, L. A., Senna, M. M. & Rezaei, N. Alopecia areata: a review of disease pathogenesis. Br. J. Dermatol. 179, 1033–1048 (2018).
Paus, R., Eichmuller, S., Hofmann, U., Czarnetzki, B. M. & Robinson, P. Expression of classical and non-classical MHC class I antigens in murine hair follicles. Br. J. Dermatol. 131, 177–183 (1994).
Ito, T. et al. Collapse and restoration of MHC class-I-dependent immune privilege: exploiting the human hair follicle as a model. Am. J. Pathol. 164, 623–634 (2004).
Hamed, F. N. et al. Alopecia areata patients show deficiency of FOXP3+CD39+ T regulatory cells and clonotypic restriction of Treg TCRbeta-chain, which highlights the immunopathological aspect of the disease. PLoS ONE 14, e0210308 (2019).
Raffin, C., Vo, L. T. & Bluestone, J. A. T(reg) cell-based therapies: challenges and perspectives. Nat. Rev. Immunol. 20, 158–172 (2020).
Harries, M. J. et al. Lichen planopilaris is characterized by immune privilege collapse of the hair follicle’s epithelial stem cell niche. J. Pathol. 231, 236–247 (2013).
Meyer, K. C. et al. Evidence that the bulge region is a site of relative immune privilege in human hair follicles. Br. J. Dermatol. 159, 1077–1085 (2008).
Rosenblum, M. D., Yancey, K. B., Olasz, E. B. & Truitt, R. L. CD200, a “no danger” signal for hair follicles. J. Dermatol. Sci. 41, 165–174 (2006).
Harrist, T. J., Ruiter, D. J., Mihm, M. C. Jr. & Bhan, A. K. Distribution of major histocompatibility antigens in normal skin. Br. J. Dermatol. 109, 623–633 (1983).
Yoshida, R., Tanaka, K., Amagai, M. & Ohyama, M. Involvement of the bulge region with decreased expression of hair follicle stem cell markers in senile female cases of alopecia areata. J. Eur. Acad. Dermatol. Venereol. 25, 1346–1350 (2011).
Ito, T. et al. Maintenance of hair follicle immune privilege is linked to prevention of NK cell attack. J. Invest. Dermatol 128, 1196–1206 (2008).
Kono, M., Nagata, H., Umemura, S., Kawana, S. & Osamura, R. Y. In situ expression of corticotropin-releasing hormone (CRH) and proopiomelanocortin (POMC) genes in human skin. FASEB J. 15, 2297–2299 (2001).
Ito, N. et al. Corticotropin-releasing hormone stimulates the in situ generation of mast cells from precursors in the human hair follicle mesenchyme. J. Invest. Dermatol. 130, 995–1004 (2010).
Agudo, J. et al. Quiescent tissue stem cells evade immune surveillance. Immunity 48, 271–285 e275 (2018).
Harries, M. J. et al. Lichen planopilaris and frontal fibrosing alopecia as model epithelial stem cell diseases. Trends Mol. Med. 24, 435–448 (2018).
Ho, B. S. et al. Progressive expression of PPARGC1alpha is associated with hair miniaturization in androgenetic alopecia. Sci. Rep. 9, 8771 (2019).
Matsumura, H. et al. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science 351, aad4395 (2016).
Zhang, C. et al. Escape of hair follicle stem cells causes stem cell exhaustion during aging. Nat. Aging 1, 889–903 (2021).
Rathnayake, D. & Sinclair, R. Male androgenetic alopecia. Expert Opin. Pharmacother. 11, 1295–1304 (2010).
Chen, C. C. et al. Regenerative hair waves in aging mice and extra-follicular modulators follistatin, dkk1, and sfrp4. J. Invest. Dermatol. 134, 2086–2096 (2014).
Keyes, B. E. et al. Nfatc1 orchestrates aging in hair follicle stem cells. Proc. Natl Acad. Sci. USA 110, E4950–E4959 (2013).
Choi, S. et al. Corticosterone inhibits GAS6 to govern hair follicle stem-cell quiescence. Nature 592, 428–432 (2021).
Ge, Y. et al. The aging skin microenvironment dictates stem cell behavior. Proc. Natl Acad. Sci. USA 117, 5339–5350 (2020).
Koester, J. et al. Niche stiffening compromises hair follicle stem cell potential during ageing by reducing bivalent promoter accessibility. Nat. Cell Biol. 23, 771–781 (2021).
Salzer, M. C. et al. Identity noise and adipogenic traits characterize dermal fibroblast aging. Cell 175, 1575–1590 e1522 (2018).
Morinaga, H. et al. Obesity accelerates hair thinning by stem cell-centric converging mechanisms. Nature 595, 266–271 (2021).
Flores, A. et al. Lactate dehydrogenase activity drives hair follicle stem cell activation. Nat. Cell Biol. 19, 1017–1026 (2017).
Reichenbach, B. et al. Glutamate transporter Slc1a3 mediates inter-niche stem cell activation during skin growth. EMBO J. 37, e98280 (2018).
Peters, F. et al. Ceramide synthase 4 regulates stem cell homeostasis and hair follicle cycling. J. Invest. Dermatol. 135, 1501–1509 (2015).
Kim, C. S. et al. Glutamine metabolism controls stem cell fate reversibility and long-term maintenance in the hair follicle. Cell Metab. 32, 629–642 e628 (2020).
Nishimura, E. K., Granter, S. R. & Fisher, D. E. Mechanisms of hair graying: incomplete melanocyte stem cell maintenance in the niche. Science 307, 720–724 (2005).
Inomata, K. et al. Genotoxic stress abrogates renewal of melanocyte stem cells by triggering their differentiation. Cell 137, 1088–1099 (2009).
Dai, D. M. et al. Modeling human gray hair by irradiation as a valuable tool to study aspects of tissue aging. Geroscience 45, 1215–1230 (2023).
Zhang, B. et al. Hyperactivation of sympathetic nerves drives depletion of melanocyte stem cells. Nature 577, 676–681 (2020).
Zarei, M., Wikramanayake, T. C., Falto-Aizpurua, L., Schachner, L. A. & Jimenez, J. J. Low level laser therapy and hair regrowth: an evidence-based review. Lasers Med. Sci. 31, 363–371 (2016).
Fertig, R. M., Gamret, A. C., Cervantes, J. & Tosti, A. Microneedling for the treatment of hair loss? J. Eur. Acad. Dermatol. Venereol. 32, 564–569 (2018).
Avci, P., Gupta, G. K., Clark, J., Wikonkal, N. & Hamblin, M. R. Low-level laser (light) therapy (LLLT) for treatment of hair loss. Lasers Surg. Med. 46, 144–151 (2014).
Varothai, S. & Bergfeld, W. F. Androgenetic alopecia: an evidence-based treatment update. Am. J. Clin. Dermatol. 15, 217–230 (2014).
Ellis, J. A. & Sinclair, R. D. Male pattern baldness: current treatments, future prospects. Drug Discov. Today 13, 791–797 (2008).
Xing, L. et al. Alopecia areata is driven by cytotoxic T lymphocytes and is reversed by JAK inhibition. Nat. Med. 20, 1043–1049 (2014).
Damsky, W. & King, B. A. JAK inhibitors in dermatology: the promise of a new drug class. J. Am. Acad. Dermatol. 76, 736–744 (2017).
King, B. et al. Two phase 3 trials of baricitinib for alopecia areata. N. Engl. J. Med. 386, 1687–1699 (2022).
Goldstein, J. et al. Calcineurin/Nfatc1 signaling links skin stem cell quiescence to hormonal signaling during pregnancy and lactation. Genes Dev. 28, 983–994 (2014).
Lee, J. et al. Runx1 and p21 synergistically limit the extent of hair follicle stem cell quiescence in vivo. Proc. Natl Acad. Sci. USA 110, 4634–4639 (2013).
Kadaja, M. et al. SOX9: a stem cell transcriptional regulator of secreted niche signaling factors. Genes Dev. 28, 328–341 (2014).
Hoeck, J. D. et al. Stem cell plasticity enables hair regeneration following Lgr5(+) cell loss. Nat. Cell Biol. 19, 666–676 (2017).
Adam, R. C. et al. Temporal layering of signaling effectors drives chromatin remodeling during hair follicle stem cell lineage progression. Cell Stem Cell 22, 398–413 e397 (2018).
Suen, W. J., Li, S. T. & Yang, L. T. Hes1 regulates anagen initiation and hair follicle regeneration through modulation of hedgehog signaling. Stem Cells 38, 301–314 (2020).
Li, G. et al. SIRT7 activates quiescent hair follicle stem cells to ensure hair growth in mice. EMBO J. 39, e104365 (2020).
Acknowledgements
We thank members of the Choi laboratory for their helpful discussions and comments on the manuscript. This work was supported by the National Research Foundation of Korea (NRF) grants funded by the Korea government (MSIT) (NRF-2022R1C1C1011895, NRF-2022M3A9D3016848, NRF-RS-2023-00221112) and the Ministry of Trade, Industry and Energy and the Korea Institute for Advancement of Technology through the Program for Smart Specialization Infrastructure Construction (No. P0021109). J.H.L. was supported by the Elsa U Pardee Foundation and MCC patient gift fund. This research has been supported by a POSCO Science Fellowship from the POSCO TJ Park Foundation. This work was also supported by a BK21 FOUR Research Fellowship funded by the Ministry of Education, Republic of Korea.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lee, J.H., Choi, S. Deciphering the molecular mechanisms of stem cell dynamics in hair follicle regeneration. Exp Mol Med 56, 110–117 (2024). https://doi.org/10.1038/s12276-023-01151-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s12276-023-01151-5
