
Abstract
Zebrafish have emerged as a powerful animal model for investigating the genetic basis of hematopoiesis. Owing to its close genetic and developmental similarities to humans, combined with its rapid reproduction and extensive genomic resources, zebrafish have become a versatile and efficient platform for genetic studies. In particular, the forward genetic screening approach has enabled the unbiased identification of novel genes and pathways related to blood development, from hematopoietic stem cell formation to terminal differentiation. Recent advances in mutant gene mapping have further expanded the scope of forward genetic screening, facilitating the identification of previously unknown genes and pathways relevant to hematopoiesis. In this review, we provide an overview of the zebrafish forward screening approach for hematopoietic gene discovery and highlight the key genes and pathways identified using this method. This review emphasizes the importance of zebrafish as a model system for understanding the genetic basis of hematopoiesis and its associated disorders.
Similar content being viewed by others
Introduction
The zebrafish (Danio rerio) has been used as a model organism for studying hematopoiesis since the 1960s because of its experimental advantages, including high fecundity, optical transparency of embryos, and diverse gene manipulation techniques1. In addition, zebrafish possess evolutionarily conserved hematopoietic genes and regulatory networks, with a comprehensive characterization of all mature blood lineages and intermediates emphasizing its significance in this field2. Utilizing diverse genetic approaches in zebrafish, multiple pathways and molecular mechanisms that underlie hematopoiesis have been defined3.
Various gene editing approaches have been employed to elucidate the genes associated with hematopoietic processes using zebrafish. The knockdown approach, commonly performed by morpholinos, provides a convenient mechanism to suppress gene expression, allowing rapid insights into gene function4. However, this approach has limitations due to concerns about possible off-target effects and inconsistent knockdown efficiencies5.
Reverse genetic screening techniques, exemplified by transcription activator-like effector nucleases (TALENs) and clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9, enable the precise targeting and modification of specific genes, serving as powerful tools for validation6,7. Notably, the CRISPR system, utilizing guide RNA (gRNA) and the Cas9 enzyme, facilitates efficient gene targeting in terms of cost and time8. Over the past decade, the zebrafish model has harnessed the CRISPR system not only to produce knockout (KO) animals but also to execute precise mutagenesis through techniques such as base editing and prime editing9,10,11. While challenges such as off-target effects persist, continuous advancements are being made in these gene-editing techniques.
In addition to reverse genetic methodologies, forward genetic screening is another hallmark of zebrafish research. This unbiased approach provides a platform for identifying novel genes critical for hematopoiesis without predetermined notions. Large-scale forward genetic screening allows the identification of unsuspected players in hematopoiesis, offering insights into the intricate regulatory networks governing this process12,13.
To perform a general forward genetic screening in zebrafish, random point mutations are initially induced throughout the genome of adult male zebrafish using a chemical mutagen14,15. Inbreeding of heterozygous progeny generated from mutagenized zebrafish produces homozygous mutations in the F3 generation. Through various phenotypic characterizations of hematopoietic events, numerous mutants with hematopoietic defects have been obtained12,16. Genetic linkage mapping, along with known polymorphic repeat markers, is critical for identifying the candidate genes responsible for the phenotypes of interest. However, the identification of these causative mutations has been severely limited owing to insufficient genetic markers and laborious work demands14,17.
Targeting Induced Local Lesions in Genomes (TILLING) is a hybrid technique that combines elements from both forward and reverse genetic screening. This method involves examining genomic DNA samples from ENU-mutagenized zebrafish to detect mutations in a selected gene. Using sperm from the respective animal, the mutant line is re-established for phenotype characterization18. TILLING employs the advantages of forward genetics while also facilitating targeted analysis of specific candidate genes, resulting in the identification of over 150 loss-of-function mutations19. Although TILLING provides powerful genetic capabilities, it can be resource intensive, time-consuming, and relatively less effective at inducing complete KOs.
Recently, the advent of next-generation sequencing (NGS) has introduced new genomic and transcriptomic approaches that can sequence nearly the entire zebrafish genome of interest20,21. Using a bioinformatics pipeline, thousands of markers across almost any genome of interest can be used to identify causative mutations found by forward screening in a fast, simple, and reliable manner.
In this review, we discuss the application of forward genetic screening in zebrafish to identify significant genes and pathways related to hematopoiesis. We also summarize the key genes discovered through forward genetic approaches and technologies that have helped to dissect the molecular mechanisms of genes involved in hematopoiesis, especially in hematopoietic stem cell (HSC) development. This review provides a comprehensive overview of our current understanding of the molecular mechanisms underlying the development of the hematopoietic system, with a particular focus on zebrafish as a powerful genetic model organism.
Zebrafish hematopoiesis
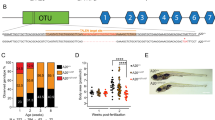
The genetic and regulatory networks of hematopoiesis in zebrafish resemble those in humans. In zebrafish, similar to other vertebrates, hematopoiesis occurs in two distinct waves: primitive and definitive hematopoiesis22. Primitive hematopoiesis produces primitive erythroid and myeloid cells by differentiating hemangioblasts, which give rise to both blood cells and vessels3,23. It mainly occurs in the anterior and posterior lateral mesoderm, which later becomes the intermediate cell mass where HSCs are congregated (Fig. 1a)22,24,25. The generated primitive erythroid and myeloid cells play a crucial role in the transfer of oxygen to tissues and immunity of early embryos and start to circulate throughout the embryos at approximately 24 h post-fertilization (hpf)26,27.

a Primitive hematopoiesis takes place in two locations, the anterior lateral mesoderm (green) and posterior lateral mesoderm (red), both of which are critical for blood and endothelial formation during the somitogenesis stages. b Definitive hematopoietic stem cells (HSCs) arise from the ventral wall of the dorsal aorta (DA) and migrate to the caudal hematopoietic tissue (CHT). HSCs then migrate from the CHT to the kidney, where they undergo lifelong hematopoiesis. HSCs then seed the thymus, where they mature and differentiate into T lymphocytes. c Definitive HSCs and other blood cells are distributed within the kidney marrow of adult zebrafish. AGM, aorta gonad mesonephros; ICM, intermediate cellular mass. Figure generated with BioRender.com.
Definitive HSCs, which are pluripotent stem cells that give rise to all blood cell lineages, begin to emerge from the ventral wall of the dorsal aorta (DA), a region known as the aorta-gonad-mesonephros, beginning approximately 30 hpf (Fig. 1b)28,29,30,31. Subsequently, at 2 days post-fertilization (dpf), these cells start migrating toward the caudal hematopoietic tissue (CHT), where they mature and continuously circulate before finally settling in the kidney (Fig. 1b)32,33. These definitive HSCs reside in the kidney marrow, which is analogous to the mammalian bone marrow, and produce all the blood lineages necessary for normal blood function from embryonic development to adulthood (Fig. 1c)22,30,34. Although many genes and pathways responsible for HSC formation and differentiation have been identified, much remains to be understood. To study the precise steps of HSC specification, differentiation, and maintenance, researchers have utilized zebrafish mutant models generated through various mutagenesis techniques.
Forward genetic screening for zebrafish hematopoiesis
N-ethyl-N-nitrosourea (ENU) random mutagenesis
Zebrafish have become a valuable model for large-scale genetic screening because of their ease of genetic manipulation. They are particularly suitable for the unbiased forward screening of specific phenotypes, which is almost exclusively practical among vertebrate models35. In forward genetics, random mutations are induced in an organism, and the resulting phenotypes are analyzed to identify mutations that affect the process of interest. Random mutagenesis can be achieved through chemical mutagens such as ENU, physical mutagens such as ionizing radiation, or transposon mutagenesis, all of which randomly induce DNA damage. Among these methods, ENU mutagenesis is a common and efficient chemical mutagenesis method in zebrafish that can induce point mutations randomly in germ cells36. In ENU mutagenesis, adult male fish are exposed to water containing ENU for approximately one hour per week over the course of three weeks. The use of anesthetics can increase the survival rate of male fish during this process. ENU mutagenesis induces random single base pair mutations with a high success rate for mutagenic loads in zebrafish premeiotic germ cells17,37. Mutagenized zebrafish can exhibit phenotypes that are discretely linked to mutated lesions in a single gene.
Phenotype screening for zebrafish hematopoiesis
Early zebrafish forward screens primarily relied on visually observing live embryos to detect specific morphological defects38. Currently, to identify mutants with hematopoietic deficiencies, physically collected mutants displaying distinct morphological defects can be screened for blood development using molecular markers for a given cell or tissue type. Whole-mount in situ hybridization (WISH) is a simple and commonly used approach for detecting the expression site of genes of interest using digoxigenin-labeled antisense RNA probes39. Using various hematopoietic markers, the emergence and differentiation of HSCs and other leukocytes can be easily examined in mutant embryos, which can elucidate whether the mutants have hematopoietic developmental defects40,41. Moreover, WISH is a useful tool for exploring various tissues and cells important for hematopoiesis, such as primitive erythroid and myeloid cells, as well as definitive HSCs and lymphoid cells (Fig. 2)42. Additionally, immunostaining using specific antibodies to observe mutant phenotypes can visualize the development of tissues essential for hematopoiesis processes41.

Representative images of WISH using probes labeling primitive erythroid (gata1), primitive myeloid (mpx, l-plastin), definitive HSCs (runx1, cmyb), and thymocytes (rag1) at distinct developmental stages.
Although WISH and immunostaining are beneficial for increasing the range of phenotypes that can be identified in mutants, these methods require sample fixation, which limits subsequent observations. Tissue-specific transgenic lines have been developed in zebrafish to mark specific cell lineages with fluorescent proteins, enabling the visualization of circulating blood cells in live animals43,44. Real-time monitoring of leukocytes facilitates the identification of hematopoietic defects in mutant zebrafish. Large-scale screening using transgenic lines is a feasible approach to assess the development of distinct blood cells and other tissues essential for hematopoiesis. By marking multiple cells and tissues with different fluorescent proteins, transgenic lines facilitate the simultaneous monitoring of the development of blood cells and other hematopoietic tissues45,46. Additionally, imaging with transgenic lines yields higher-resolution images compared to those obtained through immunostaining and WISH, leading to more reliable analyses.
Linkage genetic mapping through NGS
Traditional linkage analysis
After generating zebrafish mutants and determining their phenotypes through forward screening, the causative genes responsible for the observed phenotype should be identified to further understand the underlying genetic mechanisms. Linkage mapping is a genetic approach used to identify the genomic region responsible for a specific phenotype of interest in mutants41. This method involves using known genetic markers on zebrafish chromosomes to map the phenotypes to specific regions of the genome. Genetic markers include a variety of forms, including restriction fragment length polymorphisms, simple sequence length polymorphisms (SSLPs), insertion‒deletion polymorphisms and single-nucleotide polymorphisms (SNPs). These markers are used to track inheritance patterns and map the locations of genes of interest on the chromosome. Once the phenotype of the mutant is linked to a specific region of the genome, the genes responsible for the observed phenotypes can be identified by sequencing the genes in the identified genomic region of the mutants and recovering the altered phenotypes of the mutants by overexpressing the normal gene products of the mutated genes. Although traditional linkage mapping is a powerful tool for identifying novel genes associated with mutants generated via forward genetics, it has several limitations. This method is time-consuming and requires significant resources and expertise. Due to these difficulties, in the first large-scale zebrafish forward genetics study, only 30% of the genes in the mutants could be identified using traditional linkage mapping14.
Whole-genome sequencing (WGS)-oriented linkage mapping
Advances in genetics have led to the development of molecular tools that enable genome-wide analyses to identify the genomic region linked to a mutation. More recently, NGS has become an innovative method for identifying putative genes that may cause the phenotype of interest in zebrafish whole-genome studies15,47,48. Although WGS has been widely used to identify candidate genes in Caenorhabditis elegans and Drosophila49,50, it is not a particularly efficient method for zebrafish because of their large and highly polymorphic genome51. Thus, sequencing of a single zebrafish genome using WGS is inadequate to accurately distinguish potential causative mutations from other polymorphisms. Instead, bioinformatics approaches have been applied to improve the reliability of sequencing and genetic mapping methods. However, it does not directly address the issue of cost-effectiveness frequently associated with WGS.
To enhance the process of identifying candidate genes from forward genetic screening, RNA-seq-based bulk segregant analysis has been introduced52. This approach is not only cost-effective but also capable of mapping and identifying both genomic regions linked to mutations of interest and mutant genes. A bioinformatics pipeline that employs RNA-seq-based mapping has been developed to identify numerous genes associated with forward genetics. After conducting forward genetic screening, pools of wild-type and mutant zebrafish are separated according to their phenotypes, and their total RNA is processed to prepare RNA-seq samples (Fig. 3a). Following RNA extraction, sequencing libraries are generated for each pool, resulting in multiple samples from several different mutants during sequencing. To analyze the differential expression, RNA sequencing reads from each wild-type or mutant sample are independently aligned and assembled using the TopHat suite53. Differential expression is determined, and differentially expressed genes are identified using Cufflinks and the DEGseq R package (Fig. 3b)54.

a Diagram of the forward genetic screening approach. b Workflow for transcriptome analysis and identification of differentially expressed genes using TopHat suite and Cufflinks. c Alignment of RNA-seq reads to the reference genome using the RNA mapper pipeline, which allows visualization of the whole genome as well as the linked interval on a specific chromosome with high homozygous allele frequency. d Position and RNA-seq coverage of SNPs in the gene of interest, which resulted in the introduction of a premature stop codon.
The RNAmapper pipeline is used to input the aligned RNA sequencing datasets and identify the genomic region linked to the mutant phenotype using SNP markers52. When forward genetics yields a recessive mutation, the region linked to the mutation exhibits homozygosity. Consequently, a significant peak, indicating a high frequency of the mutant allele at and near the mutation site close to 1, is detected in the linked position of specific chromosomes (Fig. 3c). Once a linkage region is identified, all SNPs within the region are determined from the RNA-seq data of the mutant for candidate SNP filtering. The Ensembl Variant Effect Predictor tool is used to verify whether these candidate SNPs result in nonsynonymous changes55. Using the Integrative Genomics Viewer, each SNP is visually observed to ensure that the wild-type and mutant SNPs are segregated appropriately (Fig. 3d)56. Ideally, the putative mutant SNP is expected to be present in 100% of the mutants and 33% of the sibling wild-type embryos because of the presence of heterozygous siblings in the pool. Further confirmation is performed using Sanger sequencing to ensure that the candidate SNPs are present in all the mutants.
Overall, RNA-seq-based linkage mapping greatly enhances the identification of a selected number of high-priority candidate mutations associated with phenotypic traits within the linked region. The integration of computational methods has further accelerated and facilitated the reliable and cost-effective identification of causative mutations from forward genetic screens. These ongoing developments and refinements in genetic mapping methods are driving advancements that are improving and reinforcing the knowledge of vertebrate embryogenesis and human disease.
Primitive hematopoietic genes identified via forward genetics
Over the past 30 years, forward genetic screening studies using zebrafish have identified numerous genes and elucidated the genetic mechanisms involved in hematopoiesis. This section provides examples of the studies that have elucidated the specific roles of genes identified by forward screening in distinct types of hematopoiesis during zebrafish embryogenesis (Table 1).
Erythropoiesis
Primitive hematopoiesis is regulated by a complex interplay of genes, and this area of research is constantly evolving. Nusslein-Volhard et al. conducted a pioneering forward genetic screening study in the 1990s to characterize the phenotypes of mutants associated with primitive hematopoiesis12,16,57. Subsequently, mapping identified the relevant genes, and further research was conducted to determine the causes of hematopoietic defects in these mutants58,59. For instance, by confirming ENU-induced mutations based on GATA-1/GATA-2 expression and analyzing isolated blood cells, a moonshine (mon) mutant with a bloodless phenotype in the embryo was obtained12. Subsequent investigation identified the mon gene as transcriptional intermediary factor 1 gamma (tif1γ) through positional cloning using microsatellite markers, revealing that mutations in tif1γ trigger apoptosis in erythroid progenitor cells58.
In addition to tif1γ, the gata1 gene has also been identified as a primitive erythropoiesis marker through forward genetic screening. The initial mutant, named vlad tepes (vlt), was isolated by visually observing a decrease in the number of red blood cells16. Diploid mapping using centromeric markers confirmed that vlt is the gata1 gene59. Further investigation revealed that the mutated form of gata1 could not bind to the promoter of its target genes, resulting in erythropoietic defects. Another erythropoietic gene identified through forward screening is tal1, which was mapped using SSLP markers60,61. Initially discovered in a screening study to identify genes involved in blood vessel development, subsequent studies revealed that tal1 plays a critical role in red blood cell differentiation, with a lack of tal1 leading to anemia60,62. This discovery implies that hemangioblasts, which are precursors of primitive hematopoiesis, can differentiate into endothelial cells. Consequently, mutations affecting blood vessel development can also impact primitive hematopoiesis3,23.
Myelopoiesis
Forward genetic screening has also yielded mutants that cause defects in primitive myelopoiesis, which is crucial for early embryonic immunity23,27. Liu et al. conducted ENU mutagenesis and screened mutant embryos via WISH using probes for the myeloid cell markers cbfb and l-plastin63,64. A mutant named mummy (mmy) with reduced expression of both myeloid markers was identified. The mmy mutant displayed no blood circulation and a reduction in markers related to erythropoiesis, indicating that both myelopoiesis and erythropoiesis were affected. Subsequent positional cloning of the mmy mutant identified the causative mutation in the splicing factor gene dhx864.
Another mutant, moli, related to primitive myelopoiesis, was generated by ENU mutagenesis and screened using neutral red and Sudan Black to identify macrophages and neutrophils, respectively57. These mutants showed a reduction in both macrophages and neutrophils and induction of cell cycle arrest in myeloid progenitor cells. Through linkage mapping using SSLP markers, the gene encoded by the moli mutant gene was found to be a bZIP transcription factor gene, cebpα, indicating that Cebpα plays an essential role in the maintenance of neutrophils during zebrafish embryogenesis.
Forward screening for HSC development
Given that blood cell lineages are generated through HSC development, understanding the detailed molecular mechanisms of HSC emergence and differentiation is crucial. Numerous key genes associated with HSC development have been identified through forward genetic approaches involving ENU-induced mutations. For instance, the functional role of cmyb, a widely recognized HSC marker, was identified through ENU-induced forward genetic screening65,66. Zebrafish embryos carrying cmyb mutations show failure of definitive hematopoiesis, indicating the essential role of this gene in regulating HSC development. Additionally, various forward screening studies have provided valuable insights into the function of novel genes related to HSC precursor formation, HSC specification and maintenance, and the regulation of HSC development by environmental signals.
Early endothelial and blood cells
Many genetic aberrations associated with HSC emergence and differentiation have been identified through ENU mutagenesis screening. For example, the tal1 mutant phenotype exhibited a profound underdevelopment of the vasculature with the loss of not only gata1, a primitive erythrocyte marker, but also runx1, an early definitive HSC marker61. In addition, the cloche mutant embryos displayed cardiac abnormalities with a substantial reduction in both blood and vasculature formation35. The absence of GATA-2 expression, a key marker for early hematopoietic stem and progenitor cells in intermediate cellular mass (ICM), underscores the indispensable role of cloche in HSC generation26. Recent studies have revealed that the causative gene for cloche, which plays a significant role in both vascular and blood specification, is a PAS domain-containing bHLH transcription factor67.
Other mutants, such as plcγ1, tbx16, and hdac1, manifest defective axial vascular organization, artery specification, and vessel or artery formation, respectively68. Notably, all of these mutants exhibit a deficiency in HSC formation, as evidenced by the complete loss of runx1 expression. These genetic anomalies collectively accentuate the interplay between vascular and hematopoietic morphogenesis. Therefore, the development of the vasculature, including the DA, veins, and arteries, emerges as a fundamental orchestrator of HSC generation from endothelial cells in the ventral wall of the DA. Consequently, ensuring proper vascular morphogenesis is necessary for the initiation and maintenance of HSCs.
HSC specification
Forward genetic approaches have identified genes that specifically impact the formation and specification of definitive HSCs without affecting primitive hematopoiesis and vasculogenesis. Recently, we identified a novel gene, supt16h, using forward genetics69. The supt16h mutants displayed an enlarged hindbrain and a deformed tail at 32 hpf. These traits were linked to the failure of HSC formation and proliferation, and the mutants displayed normal formation of upstream or adjacent tissues and cells, including posterior lateral mesoderm, sclerotome, vasculature, primitive erythrocytes, and primitive leukocytes. Furthermore, the loss of supt16h disrupts Notch activation, a pathway that is crucial for HSC emergence and specification. Subsequent investigations revealed that the absence of supt16h leads to increased expression of p53, which elevates the expression of phc1, a Notch repressor gene, resulting in the inhibition of Notch activation and HSC formation. This study demonstrated the significant role of supt16h in the Notch pathway and the regulation of HSC development. The study also exemplifies how forward genetic screens can not only identify novel genes of interest but also guide further research on the molecular mechanisms relevant to hematopoietic development.
Expansion and maintenance of HSCs
Through forward genetic screening, numerous novel genes involved in the expansion and maintenance of definitive HSCs have been identified. Three distinct mutants generated by ENU mutagenesis (rumba, samba, and tango) were identified through reduced expression of thymic rag1, a definitive hematopoiesis marker. Subsequent positional cloning revealed that rumba encodes a novel zinc finger protein, samba encodes the mitotic spindle protein Haus3, and tango encodes SUMO-activating enzyme subunit 1 (sae1)70,71. Loss of rumba and haus3 results in defective HSC maintenance due to alterations in the cell cycle profile, whereas loss of sae1 reduces HSC survival owing to SUMOylation defects. Another ENU screening study using thymic rag1 expression identified mutated RNA polymerase III component 9 (rpc9) in the mutant line with definitive hematopoiesis defects. Loss of rpc9 leads to a defect in HSC maintenance by inducing p53-dependent apoptosis72. The ceylon (cey) mutants identified through definitive rag1 WISH exhibit severe defects in HSC maintenance40,73. SSLP-based linkage mapping identified the causative mutation in the cey mutants within the tbl3 gene, which is associated with cell cycle regulation. Moreover, grechetto mutants were identified by employing the pan-leukocyte marker mpx in the CHT at 5 dpf. Positional cloning revealed that cleavage and polyadenylation specificity factor 1 (cpsf1) is the mutated gene in the grechetto mutant74. Further characterization of the mutant suggested that proper mRNA maturation is needed for HSC survival during definitive hematopoiesis.
In addition, the cmyb WISH screening method was used to identify multiple genes related to HSC maintenance and expansion. Pan et al. obtained HSC-defective mutants exhibiting reduced cmyb expression via ENU mutagenesis and subsequently mapped the mutated genes using bulk segregation analysis along with SSLP markers. Three genes, topbp1, kri1l, and gemin5, were found to be linked to the mutant phenotypes75,76,77. Further characterization of these mutants showed that TopBP1 is involved in HSC maintenance through regulation of p53-dependent apoptosis in the CHT. Loss of the rRNA maturation factor kri1l leads to misfolded proteins in HSCs and induces autophagy via PERK signaling. gemin5 mutants exhibited a significantly decreased number of HSCs in the CHT due to defective HSC proliferation. Additionally, in a separate study, slc20a1b was found to be essential for HSC expansion in the CHT through βe1-globin WISH forward screening and subsequent analysis of cmyb expression in the CHT78,79.
Niche for HSC expansion
Forward genetic studies have provided valuable insights into the crucial roles of niches where HSC expansion and maintenance occur. Utilizing cmyb WISH screening, a genetic mutation in integrin alpha 4 (itga4) that causes severe defects in definitive hematopoiesis has been identified80. Further investigation using photoconvertible transgenic lines revealed that the HSC retention time in the CHT is reduced in the itga4 mutants. The loss of vcam1, a ligand of the Itga4-Itgb1 complex, leads to similar defects in definitive hematopoiesis as those observed in itga4 mutants. Notably, macrophages expressing vcam1 emerge as critical players in HSC retention within the CHT niche, highlighting the importance of the niche microenvironment in regulating HSC function80.
In another forward genetic study, researchers identified the oloca mutant using Sudan Black staining, which exhibited a decreased number of neutrophils compared to wild-type embryos at 3.5 dpf81. The oloca mutant carries a mutation in the naca gene, resulting in a reduced number of HSCs in the CHT, accompanied by abnormal proliferation and differentiation. Further investigations showed that Naca within the stromal niche plays a non-cell autonomous role in facilitating the settlement and formation of HSCs in the CHT. These findings highlight the significant genetic factors influencing HSC expansion and underscore the pivotal role of the CHT niche in regulating HSC function.
Summary and conclusion
In recent years, zebrafish have emerged as a powerful animal model for studying the genetic basis of hematopoiesis. Various gene editing and analysis techniques, including forward genetic screening, reverse screening, CRISPR-based approaches, and NGS, have been employed to identify genes associated with hematopoiesis and blood disorders. Notably, forward genetic screening, exemplified by methods such as ENU mutagenesis combined with linkage mapping techniques, has proven particularly advantageous in the zebrafish model. This strategy has facilitated the unbiased identification of numerous genes and pathways that regulate various aspects of blood development. In this review, we have explored the diverse genes and pathways involved in hematopoietic processes, providing critical insights into the molecular mechanisms underlying hematopoiesis via forward genetic screening.
As mutant mapping technologies advance and our knowledge of zebrafish genetics expands, we anticipate that this genetic model system will continue to revolutionize our understanding of hematopoiesis and blood disorders. Recent analyses using NGS and genome-wide association studies (GWASs) on extensive databases have highlighted potential blood disease-related genes and their associated pathological mechanisms82,83. Building on data-driven studies, zebrafish gene editing via the CRISPR system enhances our understanding of the related disease mechanisms and facilitates the development of disease models84. Collectively, these multifaceted gene editing approaches epitomize the utility of the zebrafish model in deciphering the complexities of hematopoiesis and its relevance to human health3.
References
Bahary, N. & Zon, L. I. Use of the zebrafish (Danio rerio) to define hematopoiesis. Stem Cells 16 Suppl 2, 67–78 (1998).
Traver, D. et al. Transplantation and in vivo imaging of multilineage engraftment in zebrafish bloodless mutants. Nat. Immunol. 4, 1238–1246 (2003).
Gore, A. V., Pillay, L. M., Venero Galanternik, M. & Weinstein, B. M. The zebrafish: a fintastic model for hematopoietic development and disease. Wiley Interdiscip. Rev. Dev. Biol. 7, e312 (2018).
Nasevicius, A. & Ekker, S. C. Effective targeted gene ‘knockdown’ in zebrafish. Nat. Genet 26, 216–220 (2000).
Bedell, V. M., Westcot, S. E. & Ekker, S. C. Lessons from morpholino-based screening in zebrafish. Brief. Funct. Genom. 10, 181–188 (2011).
Auer, T. O. & Del Bene, F. CRISPR/Cas9 and TALEN-mediated knock-in approaches in zebrafish. Methods 69, 142–150 (2014).
Shah, A. N., Davey, C. F., Whitebirch, A. C., Miller, A. C. & Moens, C. B. Rapid reverse genetic screening using CRISPR in zebrafish. Nat. Methods 12, 535–540 (2015).
Blackburn, P. R., Campbell, J. M., Clark, K. J. & Ekker, S. C. The CRISPR system–keeping zebrafish gene targeting fresh. Zebrafish 10, 116–118 (2013).
Li, M., Zhao, L., Page-McCaw, P. S. & Chen, W. Zebrafish genome engineering using the CRISPR-Cas9 system. Trends Genet 32, 815–827 (2016).
Petri, K. et al. CRISPR prime editing with ribonucleoprotein complexes in zebrafish and primary human cells. Nat. Biotechnol. 40, 189–193 (2022).
Rosello, M., Serafini, M., Concordet, J. P. & Del Bene, F. Precise mutagenesis in zebrafish using cytosine base editors. Nat. Protoc. 18, 2794–2813 (2023).
Ransom, D. G. et al. Characterization of zebrafish mutants with defects in embryonic hematopoiesis. Development 123, 311–319 (1996).
Driever, W. et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development 123, 37–46 (1996).
Haffter, P. et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development 123, 1–36 (1996).
Voz, M. L. et al. Fast homozygosity mapping and identification of a zebrafish ENU-induced mutation by whole-genome sequencing. PLoS One 7, e34671 (2012).
Weinstein, B. M. et al. Hematopoietic mutations in the zebrafish. Development 123, 303–309 (1996).
Patton, E. E. & Zon, L. I. The art and design of genetic screens: zebrafish. Nat. Rev. Genet 2, 956–966 (2001).
Raby, L., Volkel, P., Le Bourhis, X. & Angrand, P. O. Genetic Engineering of Zebrafish in Cancer Research. Cancers (Basel) 12, https://doi.org/10.3390/cancers12082168 (2020).
Moens, C. B., Donn, T. M., Wolf-Saxon, E. R. & Ma, T. P. Reverse genetics in zebrafish by TILLING. Brief. Funct. Genom. Proteom. 7, 454–459 (2008).
Henke, K., Bowen, M. E. & Harris, M. P. Perspectives for identification of mutations in the zebrafish: making use of next-generation sequencing technologies for forward genetic approaches. Methods 62, 185–196 (2013).
Davey, J. W. et al. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nat. Rev. Genet 12, 499–510 (2011).
de Jong, J. L. & Zon, L. I. Use of the zebrafish system to study primitive and definitive hematopoiesis. Annu Rev. Genet. 39, 481–501 (2005).
Chen, A. T. & Zon, L. I. Zebrafish blood stem cells. J. Cell Biochem 108, 35–42 (2009).
Bertrand, J. Y., Cisson, J. L., Stachura, D. L. & Traver, D. Notch signaling distinguishes 2 waves of definitive hematopoiesis in the zebrafish embryo. Blood 115, 2777–2783 (2010).
Hadland, B. K. et al. A requirement for Notch1 distinguishes 2 phases of definitive hematopoiesis during development. Blood 104, 3097–3105 (2004).
Davidson, A. J. & Zon, L. I. The ‘definitive’ (and ‘primitive’) guide to zebrafish hematopoiesis. Oncogene 23, 7233–7246 (2004).
Forrester, A. M., Berman, J. N. & Payne, E. M. Myelopoiesis and myeloid leukaemogenesis in the zebrafish. Adv. Hematol. 2012, 358518 (2012).
Thompson, M. A. et al. The cloche and spadetail genes differentially affect hematopoiesis and vasculogenesis. Dev. Biol. 197, 248–269 (1998).
Burns, C. E. et al. Isolation and characterization of runxa and runxb, zebrafish members of the runt family of transcriptional regulators. Exp. Hematol. 30, 1381–1389 (2002).
Bertrand, J. Y. et al. Haematopoietic stem cells derive directly from aortic endothelium during development. Nature 464, 108–111 (2010).
Kissa, K. & Herbomel, P. Blood stem cells emerge from aortic endothelium by a novel type of cell transition. Nature 464, 112–115 (2010).
Murayama, E. et al. Tracing hematopoietic precursor migration to successive hematopoietic organs during zebrafish development. Immunity 25, 963–975 (2006).
Jin, H., Xu, J. & Wen, Z. Migratory path of definitive hematopoietic stem/progenitor cells during zebrafish development. Blood 109, 5208–5214 (2007).
Kissa, K. et al. Live imaging of emerging hematopoietic stem cells and early thymus colonization. Blood 111, 1147–1156 (2008).
Stainier, D. Y. Zebrafish genetics and vertebrate heart formation. Nat. Rev. Genet 2, 39–48 (2001).
Solnica-Krezel, L., Schier, A. F. & Driever, W. Efficient recovery of ENU-induced mutations from the zebrafish germline. Genetics 136, 1401–1420 (1994).
Knapik, E. W. ENU mutagenesis in zebrafish–from genes to complex diseases. Mamm. Genome 11, 511–519 (2000).
Solnica-Krezel, L. et al. Mutations affecting cell fates and cellular rearrangements during gastrulation in zebrafish. Development 123, 67–80 (1996).
Thisse, C. & Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 3, 59–69 (2008).
Trede, N. S. et al. Zebrafish mutants with disrupted early T-cell and thymus development identified in early pressure screen. Dev. Dyn. 237, 2575–2584 (2008).
Lawson, N. D. & Wolfe, S. A. Forward and reverse genetic approaches for the analysis of vertebrate development in the zebrafish. Dev. Cell 21, 48–64 (2011).
Clements, W. K. & Traver, D. Signalling pathways that control vertebrate haematopoietic stem cell specification. Nat. Rev. Immunol. 13, 336–348 (2013).
Jin, S. W. et al. A transgene-assisted genetic screen identifies essential regulators of vascular development in vertebrate embryos. Dev. Biol. 307, 29–42 (2007).
Chi, N. C. et al. Genetic and physiologic dissection of the vertebrate cardiac conduction system. PLoS Biol. 6, e109 (2008).
Lawson, N. D. & Weinstein, B. M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 248, 307–318 (2002).
Covassin, L. D. et al. A genetic screen for vascular mutants in zebrafish reveals dynamic roles for Vegf/Plcg1 signaling during artery development. Dev. Biol. 329, 212–226 (2009).
Leshchiner, I. et al. Mutation mapping and identification by whole-genome sequencing. Genome Res. 22, 1541–1548 (2012).
Obholzer, N. et al. Rapid positional cloning of zebrafish mutations by linkage and homozygosity mapping using whole-genome sequencing. Development 139, 4280–4290 (2012).
Sarin, S., Prabhu, S., O’Meara, M. M., Pe’er, I. & Hobert, O. Caenorhabditis elegans mutant allele identification by whole-genome sequencing. Nat. Methods 5, 865–867 (2008).
Blumenstiel, J. P. et al. Identification of EMS-induced mutations in Drosophila melanogaster by whole-genome sequencing. Genetics 182, 25–32 (2009).
Bowen, M. E., Henke, K., Siegfried, K. R., Warman, M. L. & Harris, M. P. Efficient mapping and cloning of mutations in zebrafish by low-coverage whole-genome sequencing. Genetics 190, 1017–1024 (2012).
Miller, A. C., Obholzer, N. D., Shah, A. N., Megason, S. G. & Moens, C. B. RNA-seq-based mapping and candidate identification of mutations from forward genetic screens. Genome Res. 23, 679–686 (2013).
Trapnell, C., Pachter, L. & Salzberg, S. L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111 (2009).
Trapnell, C. et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 7, 562–578 (2012).
Flicek, P. et al. Ensembl 2011. Nucleic Acids Res 39, D800–806, (2011).
Thorvaldsdottir, H., Robinson, J. T. & Mesirov, J. P. Integrative Genomics Viewer (IGV): high-performance genomics data visualization and exploration. Brief. Bioinform. 14, 178–192 (2013).
Dai, Y. et al. Cebpalpha is essential for the embryonic myeloid progenitor and neutrophil maintenance in zebrafish. J. Genet Genom. 43, 593–600 (2016).
Ransom, D. G. et al. The zebrafish moonshine gene encodes transcriptional intermediary factor 1gamma, an essential regulator of hematopoiesis. PLoS Biol. 2, E237 (2004).
Lyons, S. E. et al. A nonsense mutation in zebrafish gata1 causes the bloodless phenotype in vlad tepes. Proc. Natl Acad. Sci. USA 99, 5454–5459 (2002).
Habeck, H. et al. Analysis of a zebrafish VEGF receptor mutant reveals specific disruption of angiogenesis. Curr. Biol. 12, 1405–1412 (2002).
Bussmann, J., Bakkers, J. & Schulte-Merker, S. Early endocardial morphogenesis requires Scl/Tal1. PLoS Genet 3, e140 (2007).
Qian, F. et al. Distinct functions for different scl isoforms in zebrafish primitive and definitive hematopoiesis. PLoS Biol. 5, e132 (2007).
Blake, T. et al. Zebrafish homolog of the leukemia gene CBFB: its expression during embryogenesis and its relationship to scl and gata-1 in hematopoiesis. Blood 96, 4178–4184 (2000).
English, M. A. et al. Incomplete splicing, cell division defects, and hematopoietic blockage in dhx8 mutant zebrafish. Dev. Dyn. 241, 879–889 (2012).
Paik, E. J. & Zon, L. I. Hematopoietic development in the zebrafish. Int J. Dev. Biol. 54, 1127–1137 (2010).
Soza-Ried, C., Hess, I., Netuschil, N., Schorpp, M. & Boehm, T. Essential role of c-myb in definitive hematopoiesis is evolutionarily conserved. Proc. Natl Acad. Sci. USA 107, 17304–17308 (2010).
Reischauer, S. et al. Cloche is a bHLH-PAS transcription factor that drives haemato-vascular specification. Nature 535, 294–298 (2016).
Burns, C. E. et al. A genetic screen in zebrafish defines a hierarchical network of pathways required for hematopoietic stem cell emergence. Blood 113, 5776–5782 (2009).
Espanola, S. G. et al. Haematopoietic stem cell-dependent Notch transcription is mediated by p53 through the Histone chaperone Supt16h. Nat. Cell Biol. 22, 1411–1422 (2020).
Du, L. et al. Rumba and Haus3 are essential factors for the maintenance of hematopoietic stem/progenitor cells during zebrafish hematopoiesis. Development 138, 619–629 (2011).
Li, X., Lan, Y., Xu, J., Zhang, W. & Wen, Z. SUMO1-activating enzyme subunit 1 is essential for the survival of hematopoietic stem/progenitor cells in zebrafish. Development 139, 4321–4329 (2012).
Wei, Y., Xu, J., Zhang, W., Wen, Z. & Liu, F. RNA polymerase III component Rpc9 regulates hematopoietic stem and progenitor cell maintenance in zebrafish. Development 143, 2103–2110 (2016).
Hutchinson, S. A. et al. Tbl3 regulates cell cycle length during zebrafish development. Dev. Biol. 368, 261–272 (2012).
Bolli, N. et al. cpsf1 is required for definitive HSC survival in zebrafish. Blood 117, 3996–4007 (2011).
Gao, L. et al. TopBP1 governs hematopoietic stem/progenitor cells survival in zebrafish definitive hematopoiesis. PLoS Genet 11, e1005346 (2015).
Jia, X. E. et al. Mutation of kri1l causes definitive hematopoiesis failure via PERK-dependent excessive autophagy induction. Cell Res. 25, 946–962 (2015).
Liu, X. et al. Mutation of Gemin5 causes defective hematopoietic stem/progenitor cells proliferation in zebrafish embryonic hematopoiesis. Front Cell Dev. Biol. 9, 670654 (2021).
Chen, J. et al. Slc20a1b is essential for hematopoietic stem/progenitor cell expansion in zebrafish. Sci. China Life Sci. 64, 2186–2201 (2021).
Wang, K. et al. Large-scale forward genetic screening analysis of development of hematopoiesis in zebrafish. J. Genet Genom. 39, 473–480 (2012).
Li, D. et al. VCAM-1(+) macrophages guide the homing of HSPCs to a vascular niche. Nature 564, 119–124 (2018).
Murayama, E. et al. NACA deficiency reveals the crucial role of somite-derived stromal cells in haematopoietic niche formation. Nat. Commun. 6, 8375 (2015).
Manier, S., Salem, K., Glavey, S. V., Roccaro, A. M. & Ghobrial, I. M. Genomic aberrations in multiple myeloma. Cancer Treat. Res 169, 23–34 (2016).
Boddu, P. C. & Kadia, T. M. Molecular pathogenesis of acquired aplastic anemia. Eur. J. Haematol. 102, 103–110 (2019).
Xie, Y. et al. ARHGEF12 regulates erythropoiesis and is involved in erythroid regeneration after chemotherapy in acute lymphoblastic leukemia patients. Haematologica 105, 925–936 (2020).
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (2022R1A2C100677812). The illustrations were generated using BioRender.com.
Author information
Authors and Affiliations
Contributions
H.S., U.S., U.N., and Y.L. discussed and commented on the paper. H.S. and Y.L. created the figures. H.S. and Y.L. revised and reviewed the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Song, H., Shin, U., Nam, U. et al. Exploring hematopoiesis in zebrafish using forward genetic screening. Exp Mol Med 56, 51–58 (2024). https://doi.org/10.1038/s12276-023-01138-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s12276-023-01138-2
