
Abstract
Introduction:
Learning impairments are often seen in children born with low birth weight (LBW). A model with translational value for long-term effects of LBW in humans is needed to further our understanding of how LBW and cognition are related. The similarities between development stages in human infants and piglets, and the high prevalence of LBW piglets make them a naturally occurring potential model in which to study cognitive impairment associated with LBW in humans.
Results:
Although both groups learned the configurations and rapidly reduced the number of incorrect visits, the LBW piglets showed a transiently retarded acquisition of the first reversal.
Discussion:
The results of the experiment support the hypothesis that LBW is related to (mild) subsequent cognitive impairments. In the future, piglets may be suitable models for testing the effects of putative therapeutics.
Methods:
To examine this potential model, we tested pairs of LBW and NBW (normal-birth-weight) piglets in a spatial hole-board (a matrix with 4 × 4 holes in the floor) task during one acquisition and two reversal phases in their own individual configurations of rewarded holes.
Similar content being viewed by others
Main
In humans, being born with a low birth weight (LBW) after being carried to term (small for gestational age (SGA)) is a phenomenon with a fairly high prevalence that has been extensively studied. LBW occurs in the United States in 8% (2002–2003) of all neonates despite advanced prenatal care. (1). In some developing countries 16–50% of term infants are SGA, and the poor home environment in many of the countries adds to the risk of suboptimal development in these children (2). Various studies in humans have shown that being born underweight is a risk factor for cognitive deficits (3,4,5,6,7,8), learning problems (4,9), spatial orientation difficulties (10), attention problems (9), depression (11), reduced brain volume (8), and reduced academic achievement and professional attainment (12) later in life.
In commercial pig rearing, selection for high fecundity in the sows and consequent increases in litter size have resulted in more piglets being born with LBW in each litter (13,14,15). LBW is associated with a higher risk of pre- and postnatal mortality and preweaning deaths (14,16,17). However, a considerable number of vulnerable underweight piglets survive the critical period after birth.
Growth retardation in the pig fetus is suggested to be an adaptive response to poor perfusion to maintain pregnancy (18). The meat production consequences of being born underweight have been studied extensively (15,16,19). However, nothing is known about the effects of being born underweight on the cognitive capacities of these piglets if they survive. Being born with LBW may have implications for animal welfare if, for example, the ability to adapt to the housing and management conditions is impaired because of insufficient ability to control the environment and reduced adaptive capacities (20).
LBW piglets, like undersized human neonates, show characteristics of immaturity/dismaturity. They are likely to develop postnatal complications (21). We hypothesized that the underweight piglet may be a useful biomedical model for studying implications of LBW in humans, and for assessing, in future research, the effects of putative therapeutic interventions that ameliorate the adverse consequences of SGA.
It has been reported that the hippocampus is a significant predictor of cognitive functioning in human adolescents who were born SGA (8). The cognitive hole-board task is a hippocampus-dependent task (22). Therefore, we tested the hypothesis that birth weight influences cognitive performance of pigs. We did this by using a hole-board task that is suited for assessing spatial learning and memory in pigs (23) and enables the measurement of several behavioral domains (24).
Results
Spatial Memory
Birth weight did not affect the average WM performance (between-subjects effects, all F1,16 ≤ 0.56, nonsignificant; Table 1 and Figure 1a ). Both groups of piglets improved their WM performance during the three phases of the experiment. Birth weight had an effect on the time required to acquire the WM component of spatial memory during the second pattern of reward holes, i.e., during the first reversal of the experiment (birth-weight × trial block interaction, F8,128 = 2.59, P < 0.0117). During the first few trial blocks of the first reversal, the WM performance of the NBW piglets was better than those of the LBW piglets; the latter group reached the same performance level as their normal weight counterparts only in the fourth block of the first reversal. This suggests that LBW slightly retarded the improvement in WM during this phase of the experiment. Acquisition of the second reversal was not affected by birth weight ( Table 1 ).

Behavior of low-birth-weight (LBW, n = 9) and normal-birth-weight (NBW, n = 9) piglets in a spatial hole-board task. Means and SEM for the six trial blocks of the acquisition phase, the nine trial blocks of the first reversal, and the four trial blocks of the second reversal are shown for (a) WM and RM, (b) trial duration and IVI, and (c) latency of the first visit and first rewarded visit. (a) open circle, WM NBW animals; filled circle, WM LBW animals; open triangle, RM NBW animals; filled triangle, RM LBW animals. (b) open circle, latency of NBW animals; filled circle, latency of LBW animals; open triangle, IVI of NBW animals; filled triangle, IVI of LBW animals. (c) open circle, 1st visit of NBW animals; filled circle, first visit of LBW animals; open triangle, first reward of NBW animals; filled triangle, first reward of LBW animals. IVI, intervisit interval; RM, reference memory; WM, working memory.
The average RM performance in the three phases of the experiment was unaffected by birth weight. Both groups of piglets acquired the different configurations efficiently. The speed of learning was similar in the two groups ( Table 1 and Figure 1a ).
Duration Measures
There were no differences between the two groups of piglets as regards trial duration, averaged over all trials. The trial duration decreased across all blocks within the three phases in a similar manner in the LBW and NBW piglet groups ( Table 1 and Figure 1b ).
A similar picture as for trial duration emerged for the IVIs. Birth weight had no effect on this measure ( Table 1 and Figure 1b ).
The latency until the first hole was visited, irrespective of whether it belonged to the rewarded set or not, did not change across the blocks of phases I, II, and III, nor did it differ between the two groups of piglets ( Table 1 and Figure 1 ).
The average latency until the piglet gained its first reward was unaffected by birth weight. However, the latency until the first rewarded visit decreased across blocks ( Table 1 and Figure 1c ). The decrease was affected by birth weight during the first reversal (birth weight × trial block interaction, F8,128 = 2.82, P < 0.0065). LBW animals initially showed a relatively high mean latency as compared to NBW animals (LBW: 42.74 s vs. NBW: 17.43 s for the first trial block of the first reversal), but this difference disappeared after the second trial block. Although inspection of Figure 1c suggests that a similar effect occurred at the beginning of the acquisition, and after switching the animals to the second reversal, this impression was not confirmed statistically.
Discussion
Animals in the LBW group showed transiently retarded acquisition of the first reversed configuration of reward holes. Acquisition of the WM component of spatial memory in the first reversal was slightly retarded for about 12 trials (three blocks) in LBW animals as compared to their NBW siblings, but this difference disappeared with training. In addition, in the LBW group, the latency to finding the first reward was substantially higher in the first trial block after switching to the first reversal.
Our experiment also corroborates previous findings (23) that piglets are able to acquire the cognitive hole-board task. LBW did not affect the acquisition of the first task.
LBW Piglets: An Animal Model of SGA in Humans?
In our study, specific but mild WM deficits were detected in LBW animals whereas RM seemed to be unaffected. Tasks that would be useful for studying WM are not often applied to SGA human infants, because of the lack of developmentally sensitive clinical measures (25); however, the few studies that have been performed do show a relationship between LBW and WM performance. It was found that 6-year-old children born with LBW show nonverbal WM deficits, and also planning and cognitive flexibility deficits (26). In another study, WM deficits were found in 5½-year old children born with LBW, but these children had all been born preterm (27). The findings in our experiment suggest that the piglets in our experiment can indeed serve as a model for human SGA infants, and the cognitive pig hole board seems to be a task sensitive enough to detect these mild deficits. Furthermore, the finding that WM deficits occurred whereas RM was spared emphasizes that these two parameters of spatial memory are independent of each other, as has been shown by others (28,29).
The LBW piglets showed mild, transient deficits when faced with the reversal task. Although these deficits were mild, it is important to consider that we found these differences between groups of piglets that consisted of animal pairs with relatively small birth-weight differences. In pigs, no standard for “underweight” or SGA animals has been defined. This makes it difficult to determine the minimal difference between an NBW or “appropriate for gestational age” animal and an SGA animal. Several standards are in use to define SGA in human neonates. For example ≥2 SD below the mean weight for the nursery of the hospital where the children were born (29,30), or according to the Finnish birth-weight charts (30). Others prefer to define as SGA all term infants weighing less than (or equal to) the sex-specific 10th percentile for gestational age at birth (31); this would include all term born children weighing between 1,500 and 2,499 g (32) or 2,500 g (2) at birth. Given these variations in the definition of SGA in humans, it is possible that more stringent criteria for definitions of LBW and NBW in piglets are needed. In our experiment, an LBW piglet was defined as one weighing at least 1 SD below the litter average. On the third day after birth, the average weight difference between pairs of siblings was 549 g (SD 272 g, minimal difference within pairs 300 g, maximal difference 980 g) with absolute weights varying between 960 and 2,525 g. In the future, this type of research would require testing pairs of siblings with larger birth-weight differences, in accordance with the stricter definition in human research.
All of the piglets had been delivered naturally without complications. However, the possibility that hypoxic–ischemic episodes may have occurred during delivery could not be entirely excluded. We did not observe motor or other deficits in any of the animals, and therefore, the possibility of hypoxia is expected to be limited. In future studies it would be advantageous to control the delivery process to prevent hypoxia, so as to avoid any interference from extraneous factors other than those related to SGA and growth retardation.
Motivation and Response Flexibility
Hand in hand with the difficulty in reversal learning comes an increased latency to finding the first rewarded bowl. Indeed, LBW animals did need more time as compared to their NBW siblings in the first trials after switching to the new reward pattern. Although LBW animals showed a retarded acquisition of the first reversal, they completed a trial in the same time period as their NBW siblings (trial duration). As with trial duration, no differences were found in IVI, indicating that LBW and NBW animals are equally motivated and physically able to perform the cognitive hole-board task.
The initial latency until an animal found its first reward differed between LBW and NBW animals during the initial stage of acquiring the first reversal. As the latency to reach the first bowl in general (rewarded or unrewarded) did not differ between groups, we hypothesize that the LBW animals showed reduced response flexibility or behavioral inhibition when confronted with an unexpected change in the test environment, which would have become noticeable when the first expected reward was not found. This hypothesis is in line with the finding that growth-retarded children with LBW appear to be less adaptive to changing test conditions. These children also showed difficulties in producing correct strategy solutions (10). Therefore future studies should include measurements for “flexibility to change” and “strategy-related differences.” The cognitive pig hole board is a suitable apparatus for collecting such measures.
Modifications of the Hole-Board Apparatus and Procedure
Compared with the setup used by others (23) our hole-board apparatus and training procedure had some modifications. We provided the hole-board apparatus with four entries instead of one, thereby reducing the likelihood that the task is solved in a nonspatial manner (33). Randomizing entry positions across trials did not seem to influence learning speed. On average, the WM and RM scores reported in a previous pig hole-board study (23) were reached in our study in ~25 trials, thereby indicating that the cognitive pig hole board yields stable and repeatable results.
Methods
Animals
Pigs ((Terra × Finnish landrace) × Duroc) were bred at the farm at Utrecht University under conventional Dutch commercial pig housing conditions. Nine pairs of female piglets were selected (each pair from a different litter), based on the body mass measured on day 3 after birth (= day 0). Male piglets were not selected, because they were routinely castrated.
Each pair consisted of one LBW piglet and one NBW piglet. Based on the average weight and SD of a litter of piglets, all those with a body mass at least 1 SD below the litter average were defined as LBW. Of these, the female piglet with the lowest body weight was selected. Next, the average weight of the litter was recalculated excluding all LBW piglets, and the female with a weight closest to this newly defined litter average was selected as the NBW piglet.
Housing
The piglets were mixed and moved to the experimental unit 2 days after weaning, at 4 weeks of age. Litter pairs were randomly assigned to one of two adjacent, enriched pens, each measuring 4 × 5 m, in a naturally ventilated stable. Consequently, one pen housed four pairs of piglets, and the other housed five pairs. Minimal and maximal temperatures in the stable were registered daily, and ranged between 3 °C and 31 °C.
Each pen contained a covered piglet nest, and straw bedding covered the concrete floor. The ambient temperature and humidity conditions inside the nest fell within the piglets’ thermoneutral zone. Food and water were provided ad libitum to all animals until week 10 of the experiment. To see whether motivational differences were associated with changes in feeding schedules, we introduced the following feeding schedule during the final 4 weeks of the experiment: 10 of the animals (which were housed together) were fed 1/3 of the daily food amount ~1 h before the start of behavioral testing (~8 AM) and the remaining 2/3 of the daily food was fed after testing, at ~4 PM; the other eight animals were fed ad libitum. The building had natural lighting and a radio played between 7 AM and 7 PM.
Testing Room
The testing room was located adjacent to the housing room, and the animals could access it through a corridor leading to a (straw-enriched) waiting area. The testing apparatus contained an abundance of extramaze cues. All animals housed together in a pen were let out through the corridor to the waiting area at the same time, and were tested individually in the order in which they arrived at the door into the waiting area. During testing a radio was played continuously to minimize the effects of sudden background noises. Individual pigs entering the testing apparatus were still able to hear and smell their pen mates.
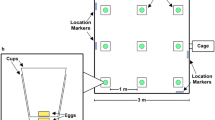
The apparatus, manufactured by Ossendrijver BV (Achterveld, The Netherlands), was a cognitive pig hole board ( Figure 2a ) consisting of a square arena with a 4 × 4 matrix of food bowls. The blue synthetic floor was slatted and the grey synthetic walls (height: 80 cm) had a steel bar across the top. The arena could be entered through any one of the guillotine doors positioned at each of the four side walls, operated from the outside. The entry door for each animal was determined individually by permutations of the numbers 1–4. By voluntarily walking down a small corridor that surrounded the arena, the animals would find an open door and enter the hole-board. The apparatus (arena and corridor) was of the testing area elevated above the floor.

Schematic hole-board outline. (a) Outline of the hole-board used to test spatial memory in piglets. Through the main entrance pigs enter the corridor (width: 40 cm) leading to the open guillotine door (any one of four doors) giving access to the test arena (530 × 530 cm) with 16 symmetrically arranged food bowls (space between bowls: 95 cm, space between wall and bowls: 73 cm, wall height: 80 cm). (b) Configurations. One of the four different hole-board configurations (a–d) was assigned to each LBW–NBW pair of siblings in the spatial hole-board task as starting configuration (acquisition). For the first reversal a new configuration was chosen from the patterns a–d, and for the second reversal from patterns e–h). LBW, low birth weight; NBW, normal birth weight.
To prevent the animals from locating rewards on the basis of smell, the food rewards (fresh M&M milk chocolates replaced daily) were placed under the false bottom of the food bowls. To prevent the animals from locating the rewards visually, each bowl was covered with a synthetic red ball (JollyBall Dog Toy, diameter: 24 cm, weight: 400 g). The animal would have to lift this ball with its snout, and the reward would then be available for consumption. The design of the apparatus was such that the ball would fall back into place and cover the bowl once the animal retracted its head.
Whenever an animal soiled the apparatus during a trial, the area was cleaned with water before the trial of the next animal. The entire apparatus was rinsed with water daily.
The experiment lasted for a total of 14 weeks. After weaning at 4 weeks of age, the animals were allowed to become habituated to their pen mates and to the new environment in the experimental unit. The animals were gradually exposed to their handlers, the testing room, and the apparatus (two daily sessions, 40 min/pen). Before formal training started, the animals were trained to lift the balls to find rewards and habituated to being alone in the testing apparatus. The group size in the apparatus was therefore gradually decreased (from 8–10 animals to 1 animal) during the 5-week habituation period. After successful habituation, when the animals were 9 weeks of age, formal training started, consisting of three phases (acquisition, first reversal, and second reversal). The animals underwent one trial in the morning and another in the afternoon (spaced trials).
Each of the 9-week old animals was assigned its own configuration of reward holes (4 of 16). Four different configurations were used ( Figure 2b ). Each piglet was trained on its particular configuration for 26 spaced trials (13 week days). The entrance door was randomly chosen for each trial. After 26 trials, asymptotic working memory (WM) performance was achieved, and training moved to the next phase.
The first reversal consisted of 38 spaced trials (19 week days). Each individual animal was assigned a changed configuration of rewarded bowls (e.g., a change from A to B–D, Figure 2b ). At this stage, we also wished to investigate the effects of food restriction on task performance. For this purpose, after 13 days (26 trials) of first reversal training, the feeding schedule of the animals in one of the pens was changed to two feedings a day (see Housing section).
The second reversal was performed for 8 week days. In this phase, each of the pigs received 16 spaced training trials on a new pattern of reward holes (e.g., a change from A to E–H, Figure 2b ). After 10 trials, apple pieces were substituted in place of chocolates for all animals, to look for possible reward-related motivational differences.
Because multiple performance measures were registered, no specific criterion of learning was employed. When WM and RM levels of performance became comparable to those reported by others (23), we introduced a reversed configuration.
The number of rewarded (correct) and unrewarded (error) visits, the number of revisits to previously rewarded bowls (errors), the latency (time elapsed) between the first (general) visit and rewarded visit, and the total trial duration were recorded in real time using ANY-maze software (Stoelting, Dublin, Ireland). A visit was scored when a pig lifted a ball with its snout and an opening between the bowl and the ball became visible. A visit to a previously visited rewarded bowl was scored as a revisit only if at least one other bowl was visited in between. Scoring started when a pig entered the arena with both forelegs. A trial was terminated when a piglet had found and consumed all four food rewards or when 600 s had elapsed, whichever event occurred earlier.
The measures “WM,” “RM,” “trial duration,” “IVIs”, and “latency to first (rewarded) hole visit” were calculated and analyzed statistically. Shapiro–Wilk tests for normality of small samples were performed per group for all parameters. This analysis revealed that 65% of all measures were distributed normally. In view of this finding, and because ANOVAs are robust with respect to deviations from normal distribution, we decided to perform all statistical analyses on the untransformed measures.
WM ratio, a measure expressing the percentage of all visits to the set of holes that yielded a food reward, was calculated as (number of rewarded visits)/(number of visits and revisits to the reward set of holes) (23).
RM ratio, a measure expressing the number of visits to the reward set of holes as a percentage of the total number of visits to all the holes, was calculated as (number of visits and revisits to the reward set of holes)/(number of visits and revisits to all holes) (23).
Trial duration was the time elapsed between entering the hole-board and finding the last reward. If the piglet did not find all of the rewards, this measure was assigned the maximum value (600 s).
IVI, the time per hole visit, was the average time elapsed between two successive hole visits. This measure was calculated as (time elapsed between the first and the last hole visit)/(the number of hole visits – 1).
Latency of first hole visit (the time elapsed up to the first hole visit) was the duration between entering the hole board and the first contact with a hole, irrespective of whether it belonged to the reward or no-reward set.
Latency to gaining first reward was the time (s) between entering the hole board and finding the first food reward.
For each of these measures block mean values of four trials each were calculated (with the exception of the first block mean value, which was the average of six trial values).
First, we analyzed whether the procedural adjustments during the course of the hole-board experiment—the change in feeding schedule starting during the first reversal and the reward change during the second reversal—affected hole-board behavior. For this purpose, ANOVAs were performed, with the within-subject (repeated-measures) factor being trial block and the between-subjects factors being birth weight (LBW vs. NBW), feeding schedule (ad libitum vs. twice daily), and type of reward (M&M chocolates vs. pieces of apple). Because these procedural modifications did not differentially affect performance, we excluded these factors from further analyses, and assessed the effects of LBW per phase of the study using a repeated-measures ANOVA with the within-subjects factor being trial block and the between-subjects factor being birth weight supplemented with ANOVAs per trial block.
Conclusion
Using a cognitive hole-board task, we found some evidence to suggest that piglets born with LBW have more difficulty in switching from one learned configuration to a new one as compared to their NBW siblings. Further research is required to clarify to what extent LBW influences cognitive performance in pigs. The hole-board task proved to be suitable for testing spatial discrimination learning in pigs. However, further validation of hole-board tasks in pigs is needed to determine the level of difficulty and sensitivity necessary to reveal small differences between groups. By fine-tuning the definition for LBW/SGA in piglets, and by adding additional behavioral, physiological, and brain measures to these types of studies, this promising model for long-term cognitive effects of LBW in humans can be refined and extended.
Ethics Note
The study was approved by the ethics committee at Utrecht University, and was conducted in accordance with the recommendations of the EU directive 86/609/EEC. All efforts were taken to minimize the number of animals used, and also to minimize their suffering.
References
Ergaz Z, Avgil M, Ornoy A . Intrauterine growth restriction-etiology and consequences: what do we know about the human situation and experimental animal models? Reprod Toxicol 2005;20:301–22.
Gardner JM, Walker SP, Powell CA, Grantham-McGregor S . A randomized controlled trial of a home-visiting intervention on cognition and behavior in term low birth weight infants. J Pediatr 2003;143:634–9.
Chaudhari S, Otiv M, Chitale A, Pandit A, Hoge M . Pune low birth weight study–cognitive abilities and educational performance at twelve years. Indian Pediatr 2004;41:121–8.
Frisk V, Amsel R, Whyte HE . The importance of head growth patterns in predicting the cognitive abilities and literacy skills of small-for-gestational-age children. Dev Neuropsychol 2002;22:565–93.
Kessenich M. Developmental outcomes of premature, low birth weight, and medically fragile infants. NAINR 2003; 3:80–7.
Paz I, Gale R, Laor A, Danon YL, Stevenson DK, Seidman DS . The cognitive outcome of full-term small for gestational age infants at late adolescence. Obstet Gynecol 1995;85:452–6.
Silva A, Metha Z, O’callaghan FJ . The relative effect of size at birth, postnatal growth and social factors on cognitive function in late childhood. Ann Epidemiol 2006;16:469–76.
Martinussen M, Flanders DW, Fischl B, et al. Segmental brain volumes and cognitive and perceptual correlates in 15-year-old adolescents with low birth weight. J Pediatr 2009; 155:848–53.e1.
O’Keeffe MJ, O’Callaghan M, Williams GM, Najman JM, Bor W . Learning, cognitive, and attentional problems in adolescents born small for gestational age. Pediatrics 2003;112:301–7.
Leitner Y, Heldman D, Harel S, Pick CG . Deficits in spatial orientation of children with intrauterine growth retardation. Brain Res Bull 2005;67:13–8.
Räikkönen K, Pesonen AK, Heinonen K, et al. Depression in young adults with very low birth weight: the Helsinki study of very low-birth-weight adults. Arch Gen Psychiatry 2008;65:290–6.
Strauss RS. Adult functional outcome of those born small for gestational age: twenty-six–year follow-up of the 1970 British birth cohort. Obstet Gynecol Surv 2000;55:417–8.
Beaulieu AD, Aalhus JL, Williams NH, Patience JF . Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork. J Anim Sci 2010;88:2767–78.
Milligan BN, Fraser D, Kramer DL. Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest Prod Sci 2002; 76:181–91.
Quiniou N, Dagorn J, Gaudré D. Variation of piglets’ birth weight and consequences on subsequent performance. Livest Prod Sci 2002;78:63–70.
Gondret F, Lefaucheur L, Louveau I, Lebret B, Pichodo X, Le Cozler Y. Influence of piglet birth weight on postnatal growth performance, tissue lipogenic capacity and muscle histological traits at market weight. Livest Prod Sci 2005;93:137–46.
O’Reilly KM, Harris MJ, Mendl M, et al. Factors associated with preweaning mortality on commercial pig farms in England and Wales. Vet Rec 2006;159:193–6.
Blomberg LA, Schreier LL, Guthrie HD, et al. The effect of intrauterine growth retardation on the expression of developmental factors in porcine placenta subsequent to the initiation of placentation. Placenta 2010;31:549–52.
van der Lende T, de Jager D. Death risk and preweaning growth rate of piglets in relation to the within-litter weight distribution at birth. Livest Prod Sci 1991;28:73–84.
Broom DM . Animal welfare: concepts and measurement. J Anim Sci 1991;69:4167–75.
Cooper JE . The use of the pig as an animal model to study problems associated with low birthweight. Lab Anim 1975;9:329–36.
Oades RD, Isaacson RL . The development of food search behavior by rats: the effects of hippocampal damage and haloperidol. Behav Biol 1978;24:327–37.
Arts JW, van der Staay FJ, Ekkel ED . Working and reference memory of pigs in the spatial holeboard discrimination task. Behav Brain Res 2009;205:303–6.
Gieling ET, Nordquist RE, van der Staay FJ . Assessing learning and memory in pigs. Anim Cogn 2011;14:151–73.
Baron IS, Erickson K, Ahronovich MD, Litman FR, Brandt J . Spatial location memory discriminates children born at extremely low birth weight and late-preterm at age three. Neuropsychology 2010;24:787–94.
Ni TL, Huang CC, Guo NW . Executive function deficit in preschool children born very low birth weight with normal early development. Early Hum Dev 2011;87:137–41.
Böhm B, Smedler AC, Forssberg H . Impulse control, working memory and other executive functions in preterm children when starting school. Acta Paediatr 2004;93:1363–71.
Prickaerts J, Honig W, Schmidt BH, Blokland A . Metrifonate improves working but not reference memory performance in a spatial cone field task. Eur J Pharmacol 1999;380:61–5.
van der Staay FJ, van Nies J, Raaijmakers W . The effects of aging in rats on working and reference memory performance in a spatial holeboard discrimination task. Behav Neural Biol 1990;53:356–70.
Pihkala J, Hakala T, Voutilainen P, Raivio K . [Characteristic of recent fetal growth curves in Finland]. Duodecim 1989;105:1540–6.
Theodore RF, Thompson JM, Waldie KE, et al. Determinants of cognitive ability at 7 years: a longitudinal case-control study of children born small-for-gestational age at term. Eur J Pediatr 2009;168:1217–24.
Grantham-McGregor SM, Lira PI, Ashworth A, Morris SS, Assunçao AM . The development of low birth weight term infants and the effects of the environment in northeast Brazil. J Pediatr 1998;132:661–6.
van der Staay FJ, Krechting B, Blokland A, Raaijmakers W . The cone 
field: a spatial discrimination task for the automatic and simultaneous assessment of working and reference memory in rats. J Neurosci Methods 1990;31:13–22.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gieling, E., Park, S., Nordquist, R. et al. Cognitive performance of low- and normal-birth-weight piglets in a spatial hole-board discrimination task. Pediatr Res 71, 71–76 (2012). https://doi.org/10.1038/pr.2011.5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/pr.2011.5
This article is cited by
-
Low birth weight influences the postnatal abundance and characteristics of satellite cell subpopulations in pigs
Scientific Reports (2020)
-
Exploratory study of the effects of intra-uterine growth retardation and neonatal energy supplementation of low birth-weight piglets on their post-weaning cognitive abilities
Animal Cognition (2019)
-
Discrimination learning and judgment bias in low birth weight pigs
Animal Cognition (2019)
-
A milk formula containing maltodextrin, vs. lactose, as main carbohydrate source, improves cognitive performance of piglets in a spatial task
Scientific Reports (2018)
-
Judgement bias in pigs is independent of performance in a spatial holeboard task and conditional discrimination learning
Animal Cognition (2017)
