
Abstract
In children with congenital heart disease, female sex has been linked to greater in-hospital mortality associated with low cardiac output, yet the reasons for this are unclear. Therefore, we examined whether newborn sex differences in the heart's metabolic response to ischemia exist. Left ventricular (LV) in vivo and ischemic biopsies of newborn male and female piglets were compared. Tissue ATP, creatine phosphate (CP), glycogen, anaerobic end-products lactate and hydrogen ion (H+), and key regulatory enzymes were measured. Compared with males, newborn females displayed 14% lower ATP, 22% lower CP, and 32% lower glycogen reserves (p < 0.05) at baseline. During ischemia, newborn females accumulated 17% greater lactate and 40% greater H+ accumulation (p < 0.02), which was associated with earlier cessation of glycolysis and lower ischemic ATP levels (p < 0.02) compared with males. Newborn females demonstrated a greater ability to use their glycogen reserves, resulting in significantly lower (p < 0.003) glycogen levels throughout the ischemic period. Thus, newborn females are at a metabolic disadvantage because they exhibited lower energy levels and greater tissue lactic acidosis, both linked to an increase susceptibility to ischemic injury and impair myocardial function on reperfusion.
Similar content being viewed by others

Main
Several studies in children have documented worse outcome of females relative to males during management or after repair for congenital heart disease (1–4). Chang et al. (2) reported that female sex was associated with 51% higher odds of mortality after cardiac surgery with the highest odds of mortality being in newborns. A 2007 nationwide study confirmed this finding and reported a 31% greater risk of mortality in female children during hospitalization for cardiac surgery (4). Compared with males, other studies have also reported lower survival of female children requiring mechanical cardiopulmonary support for severe postoperative ventricular dysfunction (1,3). Although these findings suggest that even in children, female sex may influence outcome following periods of myocardial ischemia or oxygen stress, the metabolic explanation by which this may occur is currently unknown.
In the adult heart, experimental studies have already reported sex-specific differences in myocardial metabolism and response to ischemia that may potentially impair ischemic tolerance and reduce postischemic myocardial functional recovery. Compared with males, the adult female myocardium is believed to have a greater reliance on carbohydrate and glycolytic metabolism, which during ischemia is associated with a more rapid accumulation of anaerobic end-products (5–7) and increased postischemic ventricular dysfunction (6,8). In the newborn heart, experimental studies have also associated a more rapid accumulation of anaerobic end-products with premature inhibition of glycolysis (9–11), more rapid depletion of ATP, decreased myocardial ischemic tolerance (9,10), and lower postischemic functional recovery (12,13). Because in the pediatric population, low cardiac output following myocardial ischemia has been reported to be the most common cause of morbidity and mortality after repair of congenital lesions (14,15), sex-specific differences in anaerobic end-product accumulation and/or cellular energy levels during ischemia may offer a potential explanation for differences in operative outcome in male versus female children. The purpose of this study was therefore to identify sex-specific differences in baseline and ischemic metabolism in the newborn myocardium and to explore the possible metabolic explanation for this.
MATERIALS AND METHODS
All experimental procedures were approved by the University of Toronto Animal Care and Use Committee and follow the rules set out by the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (National Institutes of Health publication No. 85-23, revised 1996).
Neonatal Yorkshire piglets were used in this study because the degree of maturity of the cardiovascular system is structurally and functionally similar to that of newborn humans (16,17). In addition, their plasma hormone levels have been reported as low to nondetectible at this age (18). Male (n = 7) and female (n = 13) age-matched newborn (3 d old) piglets (male: 2.0 ± 0.07 kg versus female: 2.0 ± 0.11 kg, p = 0.883) were anesthetized with Somnotol (Sodium Pentobarbital 65 mg/mL; MTC Pharmaceuticals, Cambridge, Canada), intubated, and mechanically ventilated. An indwelling cannula was placed in the right carotid artery to monitor blood gases and blood pressure. Appropriate adjustments were made to ventilation to ensure normal blood gas and pH (Table 1). After a midline sternotomy, the heart was exposed, and a full-thickness freeze-clamp left ventricular (LV) biopsy was taken (19). Immediately after the in vivo biopsies, the heart was excised marking the onset of global myocardial ischemia and placed in substrate-free Krebs-Henseleit physiologic solution at 38°C during which serial LV biopsies were taken at 30, 45, and 60 min of ischemia. Previous work with this animal model demonstrated that at 30 min of ischemia, moderate impairments of myocardial function are noted, whereas 45 min or longer of ischemia resulted in profound functional impairment (13). All biopsies were stored at −85°C until biochemical analysis.
To determine possible sex-specific differences in metabolic potential, the maximum activities of key enzymes involved in the creatine phosphate (CP) shuttle and anaerobic glycolysis were measured using fluorometric assays (20). The following enzyme activities (mmoles/g protein/h) were measured: creatine kinase (CK, CP shuttle), glycogen phosphorylase (GP, glycogenolysis), hexokinase (HK, glucose phosphorylation), phosphofructokinase (PFK, glycolysis), and lactate dehydrogenase (LDH, anaerobic glycolysis). Sex-specific differences in myocardial baseline and ischemic metabolism were characterized through the measurement of tissue energy levels (ATP and CP), anaerobic substrate reserves (glycogen), and metabolic end-products [lactate and hydrogen ions (H+)]. Biochemical analysis for ATP, CP, glycogen, and lactate (μmoles/g dry weight) involved using fluorometric assays (21), whereas H+ content (mol/L) was determined using the homogenate method (22,23).
All values are expressed as mean ± SEM. In vivo male and female data were compared using t tests (two-tailed), whereas a two-way repeated measures ANOVA with Bonferroni post hoc test was used to compare differences between sexes over time. Significance was defined as p < 0.05.
RESULTS
Hemodynamics and blood gas parameters.
Male and female hemodynamics and blood gas parameters are presented in Table 1. Systolic, diastolic, and mean arterial pressures were similar between males and female newborn piglets. Females, however, demonstrated a significant 22% higher resting heart rate compared with males. Blood gas, pH, electrolytes, and hematocrit were similar between newborn male and female piglets. All hemodynamic and blood gas parameters fall within the normal range reported in the literature for male piglets (24).
ATP.
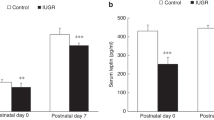
Newborn male and female in vivo and ischemic ATP levels are presented in Figure 1. Newborn female in vivo myocardial ATP levels were a significant 14% lower compared with males (males: 25.5 ± 0.9; females: 22.0 ± 0.7; p = 0.005). Overall, during ischemia, there was a significant decline in myocardial ATP levels (p < 0.03) at each ischemic time interval in both sexes. Despite females having lower levels at baseline, at 30 min of ischemia both sexes reached a similar level of ATP (p = 0.66). With continued ischemia, however, females exhibited 40% lower ATP content (p = 0.016), which was exacerbated after 60 min, where females had 70% lower ATP (p = 0.009) compared with males.

ATP levels in vivo and at 30, 45, and 60 min of ischemia in newborn male and female piglets. Values expressed as mean ± SEM: *p < 0.02 vs opposite gender at same time point, †p < 0.003 vs previous time within a gender. Males (□); females (▪).
Creatine phosphate.
In vivo CP levels were 22% lower in newborn females compared with males (males: 29.3 ± 3.0; females: 23.0 ± 1.6; p = 0.043; Fig. 2). At 30 min of ischemia, CP reserves were completely depleted in both newborn males and females.

CP levels in vivo and at 30, 45, and 60 min of ischemia in newborn male and female piglets. Values expressed as mean ± SEM: *p < 0.05 vs opposite gender at same time point and †p < 0.001 vs previous time point within a gender. Males (□); females (▪).
Glycogen.
Newborn male and female in vivo and ischemic myocardial glycogen levels are presented in Figure 3. Females displayed 32% significantly lower in vivo myocardial glycogen reserves compared with males (males: 199.8 ± 21.8; females: 133.5 ± 9.7; p = 0.007). At 30 min of ischemia, myocardial glycogen reserves declined significantly in both genders (p < 0.001); however, with continued ischemia, only the hearts of newborn females showed further significant declines (p < 0.001) in glycogen, whereas males reached a plateau after the 30 min mark. By 60 min of ischemia, females broke down a significantly greater amount of myocardial glycogen reserves compared with males (male: 50.4 ± 8.6 versus female: 75.4 ± 6.1; p = 0.028) and as a result, females continued to have lower myocardial glycogen throughout the entire ischemic period (p < 0.003).

Glycogen levels in vivo and at 30, 45, and 60 min of ischemia in newborn male and female piglets. Values expressed as mean ± SEM: *p < 0.005 vs opposite gender at same time point †p < 0.001 vs previous time within a gender, and ‡p = 0.001 vs female at 30 min of ischemia. Males (□); females (▪).
Lactate.
In vivo myocardial lactate levels were comparable between newborn males and females (males: 8.5 ± 1.2; females: 6.7 ± 0.6; p = 0.165; Fig. 4A) and were within the normal range for newborn piglets (13). After 30 min of ischemia, despite a significant accumulation (p < 0.001) of lactate in both male and female hearts, females accumulated significantly (p = 0.049) more lactate compared with males. This difference was exacerbated at 45 min of ischemia (p < 0.001) as both sexes continued to significantly (p < 0.001) accumulate lactate, after which lactate accumulation in males continued (p < 0.001), whereas in females it reached a plateau. Despite this, females still maintained higher lactate levels even at 60 min of ischemia (p = 0.01).

(A and B) Lactate and H+ levels in vivo and at 15, 30, 45, and 60 min of ischemia in newborn male and female piglets. Values expressed as mean ± SEM: *p < 0.05 vs opposite gender at same time interval, †p < 0.03 vs previous time within a gender. Males (□); females (▪).
Hydrogen ion.
There were no significant differences between newborn male and female in vivo H+ levels (males: 6.1 ± 0.3; females: 5.9 ± 0.2; p = 0.599; Fig. 4B), and these values were within the normal range for newborn piglets. After 30 min of ischemia, despite a significant increase (p < 0.001) in H+ levels in both males and females, females accumulated 31% greater H+ compared with males. This sex difference was exacerbated with continued ischemia H+ (p < 0.03) such that after 60 min of ischemia, females demonstrated 45% higher H+ content (p = 0.002) compared with males.
Myocardial enzyme activities.
There were no differences in the maximum potential activities of CPK, GP, HK, PFK, and LDH between the newborn male and female left ventricle (Table 2).
DISCUSSION
Previous work demonstrated that in newborns, compared with the right ventricle, the left ventricle had enhanced glycolytic capacity which resulted in greater anaerobic end-product accumulation and lower energy levels during ischemia (25). This study is the first to investigate and characterize sex-specific differences in baseline and ischemic metabolism in this high-risk ventricle of the newborn. The data show newborn females may be at a greater metabolic risk even before a hypoxic or ischemic stress as demonstrated by significantly lower baseline high-energy phosphates (ATP and CP) and significantly lower myocardial glycogen reserves compared with newborn males. Although no sex differences in metabolic enzymatic potential were noted, the newborn female myocardium demonstrated a greater and more rapid accumulation of anaerobic end-products indicating a greater ability to undergo anaerobic glycolysis during ischemia. Despite this, females had lower ischemic ATP levels. Greater and more rapid development of tissue lactate acidosis in the face of lower energy levels during ischemia may place the newborn female LV at greater metabolic risk compared with males, potentially negatively affecting postischemic myocardial functional recovery.
Although no parameter which may reflect ischemic tolerance or postischemic myocardial function was measured in this study, ample evidence from previous metabolic studies have demonstrated the potentially detrimental effects of the metabolic profiles identified in this study. For example, in the newborn myocardium, lower preischemic high-energy phosphate and glycogen reserves have been associated with lower ATP levels during ischemia, more rapid development of ischemic contracture, and increased postischemic ventricular dysfunction (10,13,26). In children undergoing congenital cardiac repair, slightly lower levels of preischemic ATP (∼4 μmol/g dry weight) was also strongly associated with more rapid ATP depletion during ischemia, worse postoperative myocardial function, longer time spent in the intensive care unit (ICU), and longer hospital stays (27). Other investigators have suggested that accumulation of anaerobic end-products during ischemia and not reduced ATP levels is responsible for damage to the myocardium during ischemia and reperfusion (28). The development of lactic acidosis during ischemia has been reported to trigger an ion exchange sequence resulting in Ca2+ overload, increased mitochondrial damage and cell death, and myocardial dysfunction after reperfusion (12,29,30). Based on these and other studies, the combination of lower high-energy phosphates, lower anaerobic substrate reserves (glycogen), and greater lactic acidosis during ischemia would place the newborn female myocardium at a metabolic disadvantage, resulting in significantly greater postischemic ventricular dysfunction.
Baseline metabolism.
During the perinatal period, LV workloads increase dramatically and result in rapid physiologic hypertrophy of the ventricle (31). In the adult population, both clinical and experimental studies suggest that sex of an individual may influence the response of the myocardium to pressure overload with females developing a greater degree of myocardial hypertrophy in response to similar workloads (32–34). Because increases in myocardial workloads and myocardial hypertrophy are associated with a greater reliance on carbohydrate and glycolytic metabolism (35–37), a greater response of the newborn female myocardium to perinatal changes in systemic workloads may offer a potential explanation for sex-specific differences identified in this study.
Currently, there are no studies in the literature that have reported sex-specific differences in newborn heart metabolism. Several studies in adults, however, suggest that the female myocardium has a greater reliance on glycolytic versus oxidative pathways for ATP production (6–8) and that females are more reliant on carbohydrate versus fatty acid substrates when compared with males (5). Adult females are also reported to have lower myocardial glycogen (6). Because myocardial glycogen reserves are preferentially oxidized (36,38), and because adult females are less reliant on this pathway, a lower baseline reserve of glycogen is maintained compared with that seen in males. Furthermore, because metabolism of carbohydrates are a less efficient substrate for ATP production compared with fatty acids, the greater glucose reliance of females in part explains lower levels of baseline myocardial high-energy phosphates. If these sex-specific differences in adults are also present in the newborn myocardium, this could offer a potential explanation for lower baseline myocardial glycogen reserves and lower high-energy phosphates (ATP and CP) in newborn females.
Lower baseline energy reserves in the newborn female myocardium may also be influenced by sex-specific differences in cardiac workloads and energy demand. In this study, no sex-specific differences in blood gas, pH, and pressures were noted. Interestingly, newborn female piglets demonstrated significantly higher resting heart rates compared with males. A higher resting heart rate in females has also been reported previously in both the adult and newborn myocardium (39–41). Experimental studies in adults have demonstrated that myocardial energy charge and glycogen reserves decrease in proportion to increasing heart rate (42) and this seems to also be the case in this study, because females demonstrated significantly lower myocardial high-energy phosphate and glycogen reserves.
Response during ischemia.
During ischemia, because oxygen availability begins to decline, the myocardium shifts away from oxidative processes and relies on anaerobic glycolysis as the major pathway by which to continue generating ATP. Because there is no blood flow during myocardial ischemia, anaerobic end-products such as lactate and H+ accumulate and thus serve as an indirect indicator of the degree of anaerobic glycolysis (43). During ischemia, newborn females accumulated a more rapid and greater degree of both lactate and H+, indicating that they underwent a greater amount of anaerobic glycolysis. In the newborn heart, the more rapid accumulation of anaerobic end-products during ischemia compared with adults has been reported to feedback earlier on glycolytic enzymes such as PFK to slow down or completely inhibit the glycolytic pathway (9,10,44). In this study, cessation of glycolysis occurred earlier in the newborn female myocardium, as demonstrated by lactate accumulation reaching a plateau by 45 min of ischemia, whereas in males, lactate continued to significantly accumulate until 60 min. Greater feedback inhibition of glycolysis in the newborn female myocardium may further reduce the ATP-producing ability and offer a potential explanation for their lower levels of ATP during ischemia. This is confirmed as sex-specific differences in ischemic ATP levels coincided with cessation of glycolysis in the newborn female myocardium.
During ischemia, substrate delivery is impaired and glycogen becomes one of the sole substrates available for metabolism via anaerobic glycolysis. Despite having lower levels at baseline, newborn females demonstrated a greater ability to use their glycogen reserves by breaking down over 54% by 60 min of ischemia compared with males who broke down only 25%. These results therefore demonstrate that greater accumulation of lactate in females can also be attributed to sex-specific differences in ability to undergo glycogenolysis during ischemia. It should also be noted however, that both sexes fail to completely use their glycogen reserves by 60 min of ischemia, even in the face of a rapid depletion in myocardial ATP. Incomplete utilization of glycogen reserves during ischemia has been previously reported in the newborn male myocardium (10,11) and is believed to be due to anaerobic end-product feedback inhibition of glycolysis. Interestingly, this study demonstrated that despite greater and more rapid accumulation of anaerobic end-products, the newborn female myocardium had a greater ability to undergo glycogenolysis during ischemia suggesting the presence of sex-specific differences in glycolytic enzyme sensitivity to anaerobic end-products in the newborn myocardium. In addition, previous work demonstrated that in newborns, the left ventricle despite having higher glycogen levels than the right ventricle had greater anaerobic end-product accumulation and lower energy levels during ischemia (25). The current work suggests this ischemic profile is exacerbated in females, putting them at even greater risk.
In conclusion, this study is the first to demonstrate that even in the newborn, sex-specific differences exist in myocardial metabolism and response to ischemia. Compared with males, the left ventricle of newborn females demonstrated significantly lower levels of baseline high-energy phosphates and anaerobic substrate reserves (glycogen) which may place them at potential metabolic risk even before oxygen stress. During ischemia, the newborn female myocardium demonstrated a greater ability to use glycogen reserves and accumulated a greater amount of anaerobic end-products, which was associated with earlier inhibition of glycolysis and lower ischemic ATP levels compared males. These results offer one potential explanation for clinical studies that have reported worse outcome of female children during management and after repair of congenital heart disease (1–4). Tailored cardioprotective strategies may therefore be required to help reduce the metabolic risks associated with female sex during clinical management in children.
Abbreviations
- CK:
-
creatine kinase
- CP:
-
creatine phosphate
- GP:
-
glycogen phosphorylase
- HK:
-
hexokinase
- LDH:
-
lactate dehydrogenase
- LV:
-
left ventricular
- PFK:
-
phosphofructokinase
References
Blume ED, Naftel DC, Bastardi HJ, Duncan BW, Kirklin JK, Webber SA 2006 Pediatric Heart Transplantation Investigators. Outcomes of children bridged to heart transplantation with ventricular assist devices: a multi-institutional study. Circulation 113: 2313–2319
Chang RK, Chen AY, Klitzner TS 2002 Female sex as a risk factor for in-hospital mortality among children undergoing cardiac surgery. Circulation 106: 1514–1522
Johnson TR, Schamberger MS, Hart JC, Turrentine MW, Brown JW 2003 After repair, atrioventricular valve regurgitation during cardiac extracorporeal membrane oxygenation predicts survival. Ann Thorac Surg 76: 848–852
Seifert HA, Howard DL, Silber JH, Jobes DR 2007 Female gender increases the risk of death during hospitalization for pediatric cardiac surgery. J Thorac Cardiovasc Surg 133: 668–675
Gabel SA, Walker VR, London RE, Steenbergen C, Korach KS, Murphy E 2005 Estrogen receptor beta mediates gender differences in ischemic/reperfusion injury. J Mol Cell Cardiol 38: 289–297
Saeedi R, Wambolt RB, Parsons H, Antler C, Leong HS, Keller A, Dunaway GA, Popov KM, Allard MF 2006 Gender and post-ischemic recovery of hypertrophied rat hearts. BMC Cardiovasc Disord 6: 8–20
Wittnich C, Wallen WJ 1997 Sex differences in responses to dietary manipulation in rats with hypertension and myocardial hypertrophy. Am J Clin Nutr 66: 1428–1435
Glick B, Nguyen Q, Broderick TL 2003 Depression in mechanical function following ischemia in the female rat heart: role of fatty acids and altered mitochondrial respiration. J Gend Specif Med 6: 22–26
Chiu RC, Bindon W 1987 Why are newborn hearts vulnerable to global ischemia? The lactate hypothesis. Circulation 76: V146–V149
Wittnich C, Peniston C, Ianuzzo D, Abel JG, Salerno TA 1987 Relative vulnerability of neonatal and adult hearts to ischemic injury. Circulation 76: V156–V160
Wittnich C, Belanger MP, Bandali KS 2007 Newborn hearts are at greater ‘metabolic risk' during global ischemia–advantages of continuous coronary washout. Can J Cardiol 23: 195–200
Parrish MD, Payne A, Fixler DE 1987 Global myocardial ischemia in the newborn, juvenile, and adult isolated isovolumic rabbit heart: age-related differences in systolic function, diastolic stiffness, coronary resistance, myocardial oxygen consumption, and extracellular pH. Circ Res 61: 609–615
Torrance SM, Belanger MP, Wallen WJ, Wittnich C 2000 Metabolic and functional response of neonatal pig hearts to the development of ischemic contracture: is recovery possible?. Pediatr Res 48: 191–199
Cheung MM, Smallhorn JF, Vogel M, Van Arsdell G, Redington AN 2006 Disruption of the ventricular myocardial force-frequency relationship after cardiac surgery in children: Noninvasive assessment by means of tissue Doppler imaging. J Thorac Cardiovasc Surg 131: 625–631
Hammon JW Jr, 1995 Myocardial protection in the immature heart. Ann Thorac Surg 60: 839–842
DeRoth L, Downie HG 1978 Basic cardiovascular parameters in the underweight neonatal swine. Biol Neonate 34: 155–160
Glauser EM 1966 Advantages of piglets as experimental animals in pediatric research. Exp Med Surg 24: 181–190
Bate LA, Hacker RR 1982 Estrogens and piglet viability. II. Serum estrogen concentrations in piglets. J Anim Sci 54: 1017–1022
Belanger MP, Torrance SM, Panos A, Wittnich C 1992 Multiple in vivo full-thickness myocardial biopsies by freeze-clamping. J Invest Surg 5: 143–147
Chi MM, Hintz CS, Coyle EF, Martin WH III, Ivy JL, Nemeth PM, Holloszy JO, Lowry OH 1983 Effects of detraining on enzymes of energy metabolism in individual human muscle fibers. Am J Physiol 244: C276–C287
Passonneau JV, Lowry OH 1993 Enzymatic Analysis: A Practical Guide. Humana Press, Totowa,
Bates RG 1973 Determination of pH: Theory and Practice. Wiley-Interscience, New York,
Van Slyke DD 1922 On the measurement of buffer values and on the relationship of buffer value to the dissociation constant of the buffer and the concentration and reaction of the buffer solution. J Biochem 52: 525–570
Torrance SM, Wittnich C 2001 Postischemic functional recovery in immature hearts is influenced by performance index and assessment technique. Am J Physiol Heart Circ Physiol 281: H2446–H2455
Quaglietta D, Belanger MP, Wittnich C 2008 Ventricle-specific metabolic differences in the newborn piglet myocardium in vivo and during arrested global ischemia. Pediatr Res 63: 15–19
Wittnich C, Torrance SM 1994 Does the severity of acute hypoxia influence neonatal myocardial metabolism and sensitivity to ischemia?. J Mol Cell Cardiol 26: 675–682
Najm HK, Wallen WJ, Belanger MP, Williams WG, Coles JG, Van Arsdell GS, Black MD, Boutin C, Wittnich C 2000 Does the degree of cyanosis affect myocardial adenosine triphosphate levels and function in children undergoing surgical procedures for congenital heart disease?. J Thorac Cardiovasc Surg 119: 515–524
Allard MF, Emanuel PG, Russell JA, Bishop SP, Digerness SB, Anderson P 1994 Preischemic glycogen reduction or glycogen inhibition improves postischemic recovery of hypertrophied rat hearts. Am J Physiol 267: H66–H74
Stanley WC, Lopaschuk GD, Hall JL, McCormack JG 1997 Regulation of myocardial carbohydrate metabolism under normal and ischemic conditions. Potential for pharmacological interventions. Cardiovasc Res 33: 243–257
Théroux P 1999 Protection of the myocardium cell during ischemia. Am J Cardiol 83: 3G–9G
Peterson CJ, Whitman V, Watson PA, Schuler HG, Morgan HE 1989 Mechanisms of differential growth of heart ventricles in newborn pigs. Circ Res 64: 360–369
Carroll JD, Carroll EP, Feldman T, Ward DM, Lang RM, McGaughey D, Karp RB 1992 Sex-associated differences in left ventricular function in aortic stenosis of the elderly. Circulation 86: 1099–1107
Douglas PS, Katz SE, Weinberg EO, Chen MH, Bishop SP, Lorell BH 1998 Hypertrophic remodeling: gender differences in the early response to left ventricular pressure overload. J Am Coll Cardiol 32: 1118–1125
Wallen WJ, Cserti C, Belanger MP, Wittnich C 2000 Gender-differences in myocardial adaptation to afterload in normotensive and hypertensive rates. Hypertension 36: 774–779
Taegtmeyer H, Overturf ML 1988 Effects of moderate hypertension on cardiac function and metabolism in the rabbit. Hypertension 11: 416–426
Goodwin GW, Ahmad F, Taegtmeyer H 1996 Preferential oxidation of glycogen in isolated working rat heart. J Clin Invest 97: 1409–1416
Allard MF, Schonekess BO, Henning SL, English DR, Lopaschuk GD 1994 Contribution of oxidative metabolism and glycolysis to ATP production in hypertrophied hearts. Am J Physiol 267: H742–H750
Henning SL, Wambolt R, Schonekess BO, Lopaschuk GD, Allard M 1996 Contribution of glycogen to aerobic myocardial glucose utilization. Circulation 93: 1549–1555
Nagy E, Orvos H, Bardos G, Molnar P 2000 Gender-related heart rate differences in human neonates. Pediatr Res 47: 778–780
Stein PK, Kleiger RE, Rottman JN 1997 Differing effects of age on heart rate variability in men and women. Am J Cardiol 80: 302–305
Pearl W 1996 Effects of gender, age and heart rate on QT intervals in children. Pediatr Cardiol 17: 135–136
Montgomery C, Hamilton N, Ianuzzo CD 1991 Effects of different rates of cardiac pacing on rat myocardial energy status. Mol Cell Biochem 102: 95–100
Leong HS, Grist M, Parsons H, Wambolt RB, Lopaschuk GD, Brownsey R, Allard MF 2002 Accelerated rates of glycolysis in the hypertrophied heart: are they a methodological artifact?. Am J Physiol Endocrinol Metab 282: E1039–E1045
Rovetto MJ, Lamberton WF, Neeley JR 1975 Mechanisms of glycolytic inhibition in ischemic rat hearts. Circ Res 37: 742–751
Author information
Authors and Affiliations
Corresponding author
Additional information
Supported by an operating grant (T-4926) from the Heart and Stroke Foundation of Ontario.
Rights and permissions
About this article
Cite this article
Wittnich, C., Quaglietta, D., Tan, L. et al. Sex Differences in Newborn Myocardial Metabolism and Response to Ischemia. Pediatr Res 70, 148–152 (2011). https://doi.org/10.1203/PDR.0b013e3182218c6c
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/PDR.0b013e3182218c6c
