
Abstract
To determine the influence of maternal smoking on autonomic nervous system in healthy infants, 36 infants were recorded polygraphically for one night. Their mothers were defined, according to their smoking frequency during pregnancy, as “nonsmokers” (no cigarettes smoked during pregnancy) or “smokers” (10 or more cigarettes per day). The infants had a median postnatal age of 10.5 wk (range 6 to 16 wk); 18 were born to nonsmokers, and 18 to smokers. During the whole night, spectral analyses of heart rate (HR) were evaluated as a function of sleep stages. Two major peaks were recognizable: a low-frequency component (LF) related to sympathetic and parasympathetic activities and a high-frequency component (HF) reflecting parasympathetic tonus. The ratio of LF/HF powers was calculated as an index of sympathovagal interaction. In REM sleep, “smokers” infants were characterized by significantly lower HF powers and normalized HF powers, and higher LF/HF ratios than “nonsmokers.” The finding did not reach statistical significance in NREM sleep. In conclusion, maternal smoking induced changes in autonomic control and maturation in infants. These effects of cigarette smoke exposure can be added to those already reported and offer additional evidence for counseling mothers to stop smoking.
Similar content being viewed by others


Main
Maternal smoking is associated with an increase in postnatal morbidity and mortality (1–10). Smoking has been shown to increase fetal breathing rate (4) and heart rate (HR) (11), with a decrease in fetal HR variability (4) and fetal movements' frequency (4, 11). Smoking adversely affected auditory habituation and orientation during the first 2 weeks of life (12, 13). We have reported that prenatal smoking correlated with an increase in frequency and length of obstructive apneas in newborns and infants (14).
Maternal cigarette smoke was shown to influence the development of the fetus' brain through a complex interplay of vascular, neurochemical, endocrinal, and nutritional factors (15–18). The effects of nicotine are prominent in brainstem tissues rich in cholinergic nicotinic binding receptors associated with arousal, cardiopulmonary integration, REM sleep, and somatic motor control (19).
We hypothesized that when mothers smoked during pregnancy, their offspring develop alteration in autonomic nervous control (ANS). To evaluate ANS, HR spectral analysis (fast Fourier transform) was performed during night recording on 18 infants from smoking and 18 infants from nonsmoking mothers.
METHODS
Patients.
Thirty-six subjects entered the present study if during gestation and after birth, their mothers either did not smoke (defined as “nonsmokers”), or smoked 10 or more cigarettes per day (defined as “smokers”). The infants were selected among infants admitted to join a sleep research program for sleep-related behavior. They had a median postnatal age of 10.5 wk (range: 6 to 16 wk); 18 were born to “nonsmokers” and 18 to “smokers”.
To be included in this study, none of the subjects had a history of sleep problems or apnea. At the time of the investigation, they were healthy, with no signs of infection or neurologic problems, and none was receiving medication or sedation. No infant had been deprived of sleep within the previous 24 h. The present study was approved by the Institutional Review Board, and parents gave their informed consent.
Questionnaires.
These consisted of 50 questions related to the family's medical history, the infant's prenatal and postnatal history, and the infant's behavior while awake and asleep, as well as parental consumption of illegal drugs. Mothers were asked how many cigarettes per day they smoked during the first, second, and third trimesters. The information was compiled for use in the analysis. When necessary, a pack was estimated to contain 20 cigarettes.
Polygraphic studies.
After the histories were obtained, the infants were admitted for a 9-h night monitoring session. Monitoring was conducted in a quiet, dimly lit room at an ambient temperature ranging from 20° and 23°C. All patients slept in their usual supine sleep position, without restraints. They were observed continuously during recordings, their behavior as well as nursing interventions were charted, and feeding was administrated based on demand. The data were collected on computerized systems (Alice III, Healthdyne, Respironics, Pittsburgh, PA). The following variables were simultaneously recorded: scalp EEG (C4/A1 and C3/A2); two electrooculograms; ECG; thoracic and abdominal respiratory movements; and airflow by thermistors taped under each nostril and on the side of the mouth. An actigraph was placed on one arm to measure gross body movements. Oxygen saturation was recorded continuously from a transcutaneous sensor (Nellcor, Pleasanton, CA).
Data analysis.
Thirty-second periods of the recordings were analyzed visually by two independent scorers to ensure reliability. Evaluation of the records was performed without knowledge of parental smoking habits or study hypothesis. Interrater agreement was 95%. Scoring discrepancies were discussed and codes thus agreed upon were used in the data analysis. The recordings were divided into nonrapid eye movement sleep (NREM), rapid eye movement sleep (REM), indeterminate sleep, or wakefulness according to the criteria recommended in the literature (20, 21). Sleep efficiency was defined as the ratio of the total sleep time divided by the total recording time, expressed as percent. Median values for oxygen saturation, HR, and respiratory rate were calculated on 1-min stable sleep epochs, at least 5 min after any change in body position, movement, sigh, or arousal. Overall HR variability was defined by the standard deviation of the basal HR values.
HR spectral analysis.
ECG channel was recorded at 400 Hz. Thoracic motion measured with a piezoelectric belt was sampled at 10 Hz. QRS patterns were detected on the 400 Hz ECG recording using a software algorithm. The algorithm was based on an adaptive statistical analysis of the second derivative of the ECG signal. QRS complexes were located within an error < 2.5 ms. Premature ventricular contractions and artifact RR intervals due to gross body movements, apneas, sighs, or arousals were eliminated by visual analysis of the HR data. Spectral analyses was restricted to artifact-free segments. Spectral analysis was performed following the method described by Akselrod et al. (22). The resulting power spectral density was the squared absolute value of the Fourier transform of the preprocessed signal. It was expressed in (bpm)2/Hz. The power spectral density was computed from 0 to 2 Hz. Relevant frequencies start where least five periods fit in the analyzed time segment. Therefore the relevant frequency range spanned from 0.04 to 2 Hz. Spectral analysis was performed over the whole night on successive 2-min slices of data. The frequency band was defined from the 25th to the 75th percentile of the spectral power of respiratory signal (23).
In HR power spectrum, two major peaks were recognizable: a low-frequency component (LF) defined by a center frequency of 0.1 Hz (0.04–0.15 Hz) related to sympathetic and parasympathetic activities and a high-frequency component (HF) defined by respiratory band reflecting parasympathetic tonus (24, 25).
Spectral components were represented as, power (in (bpm)2/Hz), bandwidth (in Hz), and normalized power obtained by dividing the power of the period by the total power component (in %) (22), the ratio of LF/HF powers was calculated as an index of sympathovagal interaction (26). For each patient, median HR spectral power values reached during the night in both NREM and REM sleep were calculated.
Statistical analysis.
Wilcoxon test for independent samples and χ2 test were applied. Significance was defined by a level of 0.05. Values were expressed as median and range or mean and standard deviation. Spearman test was used for correlation studies.
RESULTS
Of the 18 smoking mothers, 16 smoked 10 to 20 cigarettes per day, and 2 smoked more than 20 cigarettes per day. No difference was found between trimesters regarding smoking frequency. No smoking mother reported stopping smoking during pregnancy. After their child's birth, 7 of the 18 mothers who smoked while being pregnant increased their smoking frequency. Regular smoking of 10 to 50 cigarettes per day was reported by 13 of 18 husbands of “smokers” mothers, and 2 of 18 husbands of “nonsmokers” mothers (p < 0.001).
General Characteristics
No significant difference was seen between “smokers” and “nonsmokers” for the following variables: social class (85% class II, white collar in both groups), mothers' medical history during pregnancy: uterine bleeding, medications, consumption of alcohol (1/36 in each group), illegal drugs (no mother in each group), caffeine, frequency of breast-feeding (2/18 in “smokers” and 6/18 in “nonsmokers”), as well as the infants' gender, gestational age, or postnatal age (Table 1). Smoking mothers were younger than nonsmoking mothers (median 28 y for “smokers,” and 31 y for “nonsmokers”; range 23–45 y (p = 0.045)).
Birth weights of “smokers” infants tended to be lower than that of “nonsmokers” but the difference did not reach statistical significance. There was one small-for-gestational age subject in the “smokers” group and none in the “nonsmokers” population. No neonatal complications differentiated the two groups of subjects (one neonatal infection in the two groups).
Sleep General Characteristics
Comparing the sleep characteristics of “smokers” and “nonsmokers” infants, no difference was found for the following variables: total recording time (median of 507 min for “smokers,” and 510 min for “nonsmokers,” range values: 214–550 min), percent of awake time (median of 19.6% for “smokers,” 22.9% for “nonsmokers”; range values: 5.7–45.5%), total sleep time (TST) (median of 405.2 min for “smokers,” 381.5 min for “nonsmokers,” range values: 200–500 min), sleep efficiency (median of 80.4% for “smokers” and 77.5% for “nonsmokers”; range values: 54.5–93.7%), percent of TST spent in NREM sleep (median of 35.7% for “smokers,” 32.3% for “nonsmokers”; range values: 21.4–46.8%), or percent of TST spent in REM sleep (median of 41.9% for “smokers,” 42.1% for “nonsmokers”; range values: 16.6–63%), breathing rate in REM sleep (median of 29.5 breath/min for “smokers” and 28 breath/min for “nonsmokers”; range values: 24–42 breath/min), breathing rate in NREM sleep (median of 28 breath/min for “smokers” and 27.5 breath/min for “nonsmokers”; range values: 22–40 breath/min), basal HR in REM sleep (median of 129.5 bpm for “smokers” and 128 bpm for “nonsmokers”; range values: 111–152 bpm), basal HR in NREM sleep (median of 129 bpm for “smokers” and 125 bpm for “nonsmokers”; range values: 104–152 bpm), HR variability in REM sleep (median of 10.4 bpm for “smokers” and 12.25 bpm for “nonsmokers”; range values: 7–19.7 bpm), HR variability in NREM sleep (median of 5.9 bpm for “smokers” and 6.7 bpm for “nonsmokers”; range values: 4.5–10.1 bpm), basal oxygen saturation in REM sleep (median of 97% in both groups; range values 93–100%), and basal oxygen saturation in NREM sleep (median of 98.5% for “smokers” and 99% for “nonsmokers”; range values: 93–100%).
HR Spectral Analysis
Sleep stages.
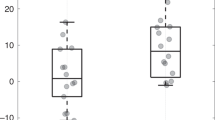
Table 2 provides a summary of the HR spectral analysis in REM and NREM sleep. Compared with “nonsmokers,” “smokers” infants were characterized during REM sleep by significantly lower HF powers and HF normalized powers, and higher LF/HF power ratios. These differences were not found in NREM sleep. No difference was seen between the two groups for total spectrum power, RR intervals, normalized or non-normalized LF powers and LF and HF bandwidth, LF and HF center frequency between the two populations in REM and NREM sleep.
Age.
With advancing postnatal age in REM sleep, HF powers (r = 0.54) and HF normalized powers (r = 0.42) increased, while LF/HF power ratios (r = −0.39) decreased in “nonsmokers.” There was no correlation with age in the “smokers” group. In NREM sleep, the same tendency was seen in the “nonsmokers” group (HF powers (r = 0.53), HF normalized powers (r = 0.30), LF/HF power ratio (r = −0.26). No correlation was seen in the “smokers” population.
Breathing frequency decreased with age in the “nonsmokers” infants in REM sleep (r = −0.37) and in NREM sleep (r = −0.28). No changes were found in the “smokers.”
Because of the small number of infants in the two groups, these differences in autonomic maturation could not be considered as significant, and have to be studied in larger groups of infants.
Paternal Smoking
In both groups of infants, the HF powers and HF normalized powers tended to be lower and the LF/HF ratios higher for infants of “smoking” fathers but the differences did not reach statistical significance.
No relationship was found between HR spectral values and time of feeding, type of feeding, or gestational age. No correlation was found between the number of cigarettes smoked and spectral analysis values.
DISCUSSION
Compared with “nonsmokers” infants, “smokers” had lower values of HF powers and normalized powers and higher LF/HF power ratios in REM sleep.
The differences in HR spectral analysis seen in the present study cannot be attributed to experimental conditions. Infants of “smokers” and “nonsmokers” mothers could not be differentiated by frequency of maternal alcohol or drug exposure, use of sedatives, previous sleep deprivation, body position during sleep (27), environmental temperature, time of the night, type or time of feeding.
Four major caveats due to the study design should be acknowledged.
First, as women who smoked while pregnant also smoked after delivery, we do not know if the effects of smoking occurred during pregnancy or after birth. In a previous study, we had found that prenatal smoking, but not postnatal smoking by mothers correlated with an increase in frequency and length of obstructive apneas in newborn infants, suggesting that the increase frequency of obstructive apnea resulted from prenatal exposure to cigarette smoking (14). The high levels of nicotine cholinergic receptors in the brainstem at midgestation and its rapidly changing profile over late gestation would suggest that mid-to-late gestation is a developmental period most vulnerable to the harmful effects of nicotine in maternal cigarette smoke (19).
Second, as smoking habits were derived from parental reports, we cannot exclude underreporting of smoking frequency, especially from pregnant women exposed to anti-smoking information (28). Measurement of biologic markers, such as cotinine in amniotic fluids, cord blood, saliva or urine of the infants could have assessed recent exposure to nicotine (28). Such measurements were not performed, but other studies showed a good correlation between urinary cotinine levels and self-reports of maternal smoking during pregnancy, indicating that purposeful misreporting of smoking habits is unusual (29, 30). In addition, only parental smoking was considered in the study and no additional source of cigarette smoke or carbon monoxide exposure was considered (31, 32). While some of the infants from “nonsmokers” could have been exposed to nicotine in utero and during the first weeks of life, the differences in spectral analysis values remained significantly different between the infants from “smokers” and “nonsmokers.”
Finally, the groups were composed of relatively small numbers of infants. Other group differences could have emerged, had more subjects been studied.
Maternal smoking apparently alters infants' autonomic nervous tonus and maturation in REM sleep. In HR power spectrum, two main frequency bands can be observed: the higher frequency band or the respiratory peak, and the lower end of the spectrum, below 0.15 Hz. The respiratory peak has been shown to be mainly vagally mediated (22, 24–26). Three mechanisms are generally proposed to explain the modulation of HR associated with respiration (33–36):1) a direct influence of medullary respiratory neurons on cardiomotor neurons (33, 37, 38);2) an indirect influence on HR of blood pressure changes secondary to respiratory movements that is mediated via arterial baroreceptors (35, 39, 40) or arterial stretch receptors (41), and 3) a reflex response to lung inflation mediated by thoracic stretch receptors (42), most likely from lungs (37, 43) and chest wall (41).
The respiratory generator is related to neural networks focused in the dorsal medullary/nucleus tractus solitarius region (dorsal respiratory group) and the periambigual area of the medulla (ventral respiratory group) (44). The cardiomotor generator has foci in the medullary lateral tegmental field, ventrolateral medulla, and raphe nuclei (45). Although normally entrained by respiration and the beat of the heart, both respiratory and cardiac rhythms can emerge from intrinsic central activities and can persist despite pulmonary and baroreceptor deafferentation (44, 45).
Maternal cigarette smoke can influence the development of the fetus' brain through a complex interplay of vascular, neurochemical, endocrinal and nutritional factors (15–18). Nicotine, transferred across placenta, can also directly interfere with brain development. In the fetal nervous system, nicotine alters cellular development and leads to early termination of cell division, mis-timing of cellular differentiation, and alters the synaptogenesis and synaptic activities of a variety of neurotransmitter pathways (46–48). These changes affect both central and peripheral neural tissues and are most prominent in tissues rich in nicotinic cholinergic receptors, such as the brainstem (47–48). Prenatal exposure to nicotine produces alterations in nicotine binding sites which were heavily concentrated in midgestation fetus, in tegmental nuclei related to cardiopulmonary integration (nucleus tractus solitarii, parabrachial complex), to regulation of arousal, attention, REM sleep (mesencephalic and pontine reticular formation), to somatic motor control (paramedian pontine and medullary reticular formation) and hypoglossal nucleus involved in tongue control and upper airway patency (19). The concentration of these receptors decreases from mid gestation toward term. These areas could be susceptible to nicotine toxicity during critical periods of brain development (19). These morphologic and metabolic changes could interfere with cardiorespiratory and autonomic maturation, especially in REM sleep.
Exposure of transdermal nicotine during pregnancy often induced an increased in baseline HR and a loss of fetal HR reactivity (49). Maternal smoking could induce autonomic changes such as a decrease in parasympathetic activity or an increase in orthosympathetic tone or a combination of both and an alteration in the maturation of these two components. There is a predominance of sympathetic over the parasympathetic activity in the fetus and in premature infants (50, 51). From 35–36 wk gestational age and after birth (52), the parasympathetic tone has a pronounced increase; nicotine exposure could change the autonomic maturation.
The clinical significance of the observed changes in autonomic cardiac controls is unknown. The observed changes in autonomic cardiac controls could reduce the electrical stability of the heart and precipitate ventricular fibrillation and sudden cardiac death (53–55). An attenuated vagal or an increased sympathetic activity could also reduce behavioral adaptation to environmental stresses (56, 57). In rats, fetal nicotine exposure produced intolerance to neonatal hypoxia by interfering with HR control, resulting in a rapid and profound fall in HR compared with normal neonates who display initial tachycardia and a gradual, slight subsequent fall in HR (58). The differences could result from impaired release of adrenal catecholamines and reduced sinoatrial responsiveness to adrenergic stimulation (58). An intact adrenal function is necessary for gasping and surviving from anoxia in newborn rats (59). But increase basal sympathoadrenal activity blunted the sympathoadrenal response to acute stresses such as hypoxia, in growth-retarded newborn rats (60).
An autonomic dysregulation could explain the inhibition of some homeostatic reflexes seen in infants exposed to tobacco smoking including a deficiency in arousal responsiveness to hypoxia or hypercapnia (61) or an increased frequency of obstructive events (14).
In conclusion, cigarette smoking induced changes in autonomic control and maturation in infants. These effects of maternal smoking can be added to those already reported and offer additional evidence for obstetricians and pediatricians counseling mothers to stop smoking during pregnancy (62).
Abbreviations
- ANS:
-
autonomic nervous system
- HF:
-
high frequency
- HR:
-
heart rate
- HRSA:
-
heart rate power spectral analysis
- LF:
-
low frequency
- LF/HF:
-
low frequency to high frequency power ratio
- NREM:
-
non-rapid eye movement sleep
- REM:
-
rapid eye movement sleep
References
Rantakallio P 1978 The effect of smoking on birth weight and the subsequent health of the child. Early Hum Dev 2: 371–382
Rantakallio P 1978 Relationship of maternal smoking to morbidity and mortality of the child up to the age of five. Acta Paediatr Scand 67: 621–631
Wall M, Brooks J, Holsclaw D, Redding G 1985 Health effects of smoking on children. Am Rev Resp Dis 132: 1137–1138
Thaler I, Goodman JDS, Dawes GS 1980 Effects of maternal cigarette smoking on fetal breathing and fetal movements. Am J Obstet Gynecol 138: 282–287
Meyer MB, Jonas BS, Tonascia JA 1976 Perinatal events associated with maternal smoking during pregnancy. Am J Epidemiol 103: 464–476
Naeye RL 1978 Relationship of cigarette smoking to congenital anomalies and perinatal death. Am J Pathol 90: 289–294
Wen SW, Goldenberg RL, Cutter GR, Hoffman HJ, Cliver SP, Davis RO, DuBard MB 1990 Smoking, maternal age, fetal growth, and gestational age at delivery. Am J Obstet Gynecol 162: 53–58
Hoffman HF, Damus K, Hillman L, Krongrad E 1988 Risk factors for SIDS: results of the National Institute of Child Health and Human Development SIDS Cooperative Epidemiological Study. Ann NY Acad Sci 533: 13–30
Martin TR, Bracken MB 1986 Association of low birth weight with passive smoke exposure in pregnancy. Am J Epidemiol 124: 633–642
American Academy of Pediatrics, Committee on Environmental Hazards. 1986 Involuntary smoking—a hazard to children. Pediatrics 77: 755–757
Kelly J, Mathews KA, O'Conor M 1984 Smoking in pregnancy effects on mother and fetus. Br J Obstet Gynaecol 91: 111–117
Saxton DW 1978 The behavior of infants whose mothers smoke in pregnancy. Early Hum Dev 2: 363–369
Picone TA, Allen LH, Olsen PN, Ferris ME 1982 Pregnancy outcome in North American women. Am J Clin Nutr 36: 1205–1213
Kahn A, Groswasser J, Sottiaux M, Kelmanson I, Rebuffat E, Franco P, Dramaix M, Wayenberg JL 1994 Prenatal exposure to cigarettes in infants with obstructive sleep apneas. Pediatrics 93: 778–783
Arbeille P, Bosc M, Vaillant MC, Tranquart F 1992 Nicotine-induced changes in the cerebral circulation in ovine fetuses. Am J Perinatol 9: 270–274
Mochizuki M, Maruo T, Masuko K, Ohtsu T 1984 Effects of smoking on fetoplacental-maternal system during pregnancy. Am J Obstet Gynecol 149: 413–420
Jauniaux E, Burton GJ 1992 The effect of smoking in pregnancy on early placental morphology. Obstet Gynecol 79: 645–648
Kuhnert BR, Kuhnert PM, Debanne S 1987 The relationship between cadmium, zinc, and birth weight in pregnant women who smoke. Am J Obstet Gynecol 157: 1247–1251
Kinney HC, O'Donnal TJ, Kriger P, White WS 1993 Early development changes in (3H) nicotine binding in human brainstem. Neuroscience 55: 1127–1130
Anders T, Emde R, Parmelee A 1971 A Manual of Standardized Terminology, Technology and Criteria for Scoring States of Sleep and Wakefulness in Newborn Infants. UCLA Brain Information Service/BRI Publication Office, Los Angeles, pp 8–11
Guilleminault C, Souquet M 1979 Sleep states and related pathology. In: Korobkin R, Guilleminault C (eds) Advances in Perinatal Neurology, Vol I. Spectrum Publications, New York, pp 225–247
Akselrod S, Gordon D, Ubel FA, Shannon DC, Barger AC, Cohen RJ 1981 Power spectral analysis of heart rate fluctuations: a quantitative probe of beat-to-beat cardiovascular control. Science 213: 220–222
Van Ravenswaaij-Arts C, Hopman J, Kollee L, Stoelingan Geijn H 1984 Spectral analysis of heart rate variability in spontaneously breathing in very preterm infants. Acta Paediatr 83: 473–480
Baselli G, Cerutti S, Civardi S, Liberati D, Lombardi F, Malliani A, Pagani M 1986 Spectral and cross-spectral analysis of heart rate and arterial blood pressure variability signals. Comput Biomed Res 19: 520–534
Pagani M, Lombardi F, Guzzetti S, Rimoldi O, Furlan R, Pizzinelli P, Sandrone G, Malfatto G, Dell'orto S, Piccaluga E 1986 Power spectral analysis of heart rate and arterial pressure variabilities as a marker of sympathovagal interaction in man and conscious dog. Circ Res 59: 178–193
Malliani A, Pagani M, Lombardi F, Cerutti S 1991 Cardiovascular neural regulation explored in the frequency domain. Circulation 84: 482–492
Franco P, Groswasser J, Sottiaux M, Broadfield E, Kahn A 1996 Decreased cardiac responses to auditory stimulation during prone sleep. Pediatrics 97: 174–178
Boyle P 1993 The hazards of passive- and active-smoking. N Engl J Med 328: 1708–1709
Stick SM, Burton PR, Gurrin L, Sly PD, LeSouef PN 1996 Effects of maternal smoking during pregnancy and a family history of asthma on respiratory function in newborn infants. Lancet 348: 1060–1064
Jaakkola MS, Jaakkola JJK 1978 Assessment of exposure to environmental tobacco smoke. Eur Respir J 10: 2384–2397
U.S. Department of Heath and Human Services. 1986 The health consequences of involuntary smoking: a report of the Surgeon General, No. (CDC) 87–8398. DHHS Publication, Rockville, MD, Office of smoking and Heath, pp 38–49
Greenberg RA, Bauman KE, Glover LH, Strecher VJ, Kleinbaum DG, Haley NJ, Stedman HC, Fowler MG, Loda FA 1989 Ecology of passive smoking by young infants. J Pediatr 114: 774–780
Bernston GG, Cacioppo JT, Quigley KS, Strecher VJ, Kleinbaum DG, Haley NS, Stedman HC, Fowler MG, Loda FA 1993 Respiratory sinus arrhythmia: autonomic origins, physiological mechanisms, and psychophysiological implications. Psychophysiology 30: 183–196
Hirsch JA, Bischop B 1981 Respiratory sinus arrhythmia in humans: how breathing pattern modulates heart rate. Am J Physiol 241: H620–H629
Katona PG, Jih F 1975 Respiratory sinus arrhythmia: non-invasive measure of parasympathetic cardiac control. J Appl Physiol 39: 801–805
Koizumi K, Teruii N, Kollai M 1985 Effects of cardiac vagal and sympathetic nerve activity on heart rate in rhythmic fluctuations. J Auton Nerv Syst 12: 251–259
Anrep GV, Pascual W, Rossler R 1936 Respiratory variation of the heart rate. Proc R Soc London Ser B 119: 191–230
Levy MN, Degeest H, Zieske H 1966 Effects of respiratory center. Circ Res 18: 67–78
Davies CTM, Neilson JMM 1967 Sinus arrhythmia in man at rest. J Appl Physiol 22: 947–955
Haymet BT, McCloskey DI 1975 Baroreceptor and chemoreceptor influence on heart rate during respiratory cycle in the dog. J Physiol Lond 245: 699–712
Melcher A 1976 Respiratory sinus arrhythmia in man. Acta Physiol Scand Suppl 435: 1–35
Clynes M 1960 Respiratory sinus arrhythmia: laws derived from computer simulation. J Appl Physiol 15: 863–874
Hainsworth R 1974 Circulatory responses from lung inflation in anesthetized dogs. Am J Physiol 226: 247–255
Richter DW, Spyer KM 1990 Cardiorespiratory control. In: Loewy AD, Spyer KM (eds) Central Regulation of Autonomic Function. Oxford University Press, New York, pp 189–207
Gebber GL 1990 Central determinants of sympathetic nerve discharge. In: Loewy AD, Spyer KM (eds). Central Regulation of Autonomic Function. Oxford University Press, New York, pp 126–144
Slotkin TA, Navarro HA, EC McCook EC, Seider FJ 1990 Fetal nicotine exposure produces postnatal up-regulation of adenylate cyclase activity in peripheral tissues. Life Sci 47: 1561–1567
Slotkin TA, Lappi SE, McCook EC, Lorber BA, Seider FJ 1995 Loss of neonatal hypoxia tolerance after prenatal nicotine exposure: implications for sudden infant death syndrome. Brain Res Bull 38: 69–75
Navarro HA, Seidler FJ, Schwartz RD, Baker FE, Dobbins SS, Slotkin TA 1989 Prenatal exposure to nicotine impairs nervous system development at a dose which does not affect viability or growth. Brain Res Bull 23: 187–192
Oncken CA, Hardardottir H, Hatsukami DK, Lupo VR, Rodis JF, Smeltzer JS 1997 Effects of transdermal nicotine or smoking on nicotine concentrations and maternal-fetal hemodynamics. Obstet Gynecol 90: 569–574
Siimes AS, Valimaki IA, Antila KJ, Julkunen MK, Metsala TH, Halkola LT, Sarafas HS 1990 Regulation of HR variation by the autonomic nervous system in neonatal lambs. Pediatr Res 27: 383–391
Curzi-Dascalova L 1995 Développement du sommeil et des fonctions sous controle du système nerveux autonome chez le nouveau-né prématuré et à terme. Arch Pédiatr 2: 255–262
Ramet J, Dehan M, Gaultier C 1991 Cardiac and respiratory vagal reactivity in infants In: Gaultier C, Escourrou P, Curzi-Dascolova L (eds) Sleep and Cardiorespiratory Control. Colloque, INSERM 217, pp 145–153
Verrier RL, Lown B 1981 Autonomic nervous system and malignant cardiac arrhythmias. In: Wiener H, Hofer MA, Stunkard AJ (eds) Brain, Behaviour and Bodily Disease, Vol 59. Raven Press, New York, pp 273–291
Schwartz PJ 1976 Cardiac sympathetic innervation and the sudden infant death syndrome: a possible pathogenetic link. Am J Med 60: 167–172
Schwartz PJ, Stone HL 1982 The role of the autonomic nervous system in sudden coronary death. Ann NY Acad Sci 382: 162–180
Porges SW 1992 Vagal tone: a physiologic marker of stress vulnerability. Pediatrics 90: 498–504
Skinner JE, Molnar M, Harper RM 1994 Higher cerebral regulation of cardiovascular and respiratory functions. In: Kryger MH, Roth T, Dement WC (eds) Principles and Practice of Sleep Medicine. WB Saunders, Philadelphia, pp 231–251
Slotkin TA, Saleh JL, McCook EC, Seidler FJ 1997 Impaired cardiac function during postnatal hypoxia in rats exposed to nicotine prenatally: implications for perinatal morbidity and mortality, and for sudden infant death syndrome. Teratology 55: 177–184
Yuan SZ, Runold M, Lagercrantz H 1997 Adrenalectomy reduces the ability of newborns rats to gasp and survive anoxia. Acta Physiol Scand 159: 285–292
Shaul PW, Cha CM, Oh W 1989 Neonatal sympathoadrenal responses to acute hypoxia: impairment after experimental intrauterine growth retardation. Pediatr Res 25: 466–472
Lewis KW, Bosque EM 1995 Deficient hypoxia awakening response in infants mothers: possible relationship to sudden infant death syndrome. J Pediatr 127: 691–696
Kessler DA, Natanblut SL, Wilkenfeld JP, Lorraine CC, May SL, Berstein IBG, Thompson L 1997 Nicotine addiction: a predictive disease. J Pediatr 130: 518–524
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Franco, P., Chabanski, S., Szliwowski, H. et al. Influence of Maternal Smoking on Autonomic Nervous System in Healthy Infants. Pediatr Res 47, 215 (2000). https://doi.org/10.1203/00006450-200002000-00011
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-200002000-00011
This article is cited by
-
Prenatal smoking and drinking are associated with altered newborn autonomic functions
Pediatric Research (2023)
-
Interaction of the pre- and postnatal environment in the maternal immune activation model
Discover Mental Health (2023)
-
Assessment of tobacco smoke effects on neonatal cardiorespiratory control using a semi-automated processing approach
Medical & Biological Engineering & Computing (2018)
-
Cardiovascular autonomic dysfunction in sudden infant death syndrome
Clinical Autonomic Research (2018)
-
Altered autonomic control in preterm newborns with impaired neurological outcomes
Clinical Autonomic Research (2015)
